transport in vascular plants
advertisement

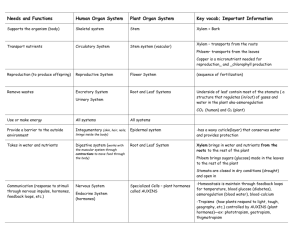
TRANSPORT IN VASCULAR PLANTS Plants absorb water and minerals through their roots and transport them to the leaves and stems for metabolic use, e. g. photosynthesis. Xylem transport water and minerals from roots to shoots. Phloem transport sugars from where they are produced or stored to where they are needed for growth and metabolism. PHYSICAL FORCES DRIVE THE TRANSPORT OF MATERIALS IN PLANTS. A variety of physical processes are involved in the transport of materials in plants. The transport in plants occurs at three scales: 1. Within cells, e. g root hairs. 2. Short-distance transport from cell to cell. 3. Long-distance transport within xylem and phloem. SELECTIVE PERMEABILITY OF MEMBRANES Transport at the cellular level depends on the selective permeability of membranes. Biological membranes are usually permeable to small molecules and lipid-soluble substance. Water, gases (O2, N2, CO2, CO), small polar molecules (glycerol), larger non-polar molecules (hydrophobic substances like hydrocarbons). Biological membranes are impermeable to and use proteins to transport the following types of molecules, Ions, amino acids and sugars, e. g. glucose. Transport methods (review chapter 7): 1. 2. 3. 4. 5. 6. Simple diffusion: Along concentration gradient. Facilitated diffusion: protein channels, along concentration gradient. Carrier-mediated transport (active transport): pumps require ATP. Cotransport Endocytosis Exocytosis Proton pumps Proton pumps play an important role in transport across the membrane. The energy stored in a proton gradient is used to transport solutes across the membrane. In chemiosmosis, the proton gradient is used to synthesize ATP. Because the proton pump moves positive charge (H+) out of the cell, the pump contributes to a voltage known as a membrane potential, a separation of opposite charges across a membrane. The proton gradient across the membrane creates membrane potential that can be harnessed to perform cellular work. Plants use the energy stored in the proton gradient and membrane potential to drive the transport of many different solutes, e. g. chemiosmosis, cotransport. Osmosis Osmosis is the passive transport of water across a membrane. Water will move across a cell membrane in the hypotonic (low solute concentration) → hypertonic direction (high solute concentration). Remember that high solute concentration means relatively low water concentration In plants, the presence of the cell wall that limits the expansion of the cell adds another factor that affects osmosis. The combine effect of solute concentration and pressure makes what is called the water potential, represented by the letter psi, ψ. Water moves across the membrane from the region of high water potential to that of low water potential. Water potential is measured in megapascals, MPa. 1 MPa = 10 atmospheres or 14.5 pounds/inch2. Examples: Your lungs exert less than 0.1 MPa. A car tire is usually inflated to about 0.2 MPa. Water pressure in home plumbing is about 0.25 MPa. Plant cells exist at about 1 MPa. Water potential for pure water in an open container is standardized a 0 MPa. Adding solutes lowers the water potential because the water molecules surrounding the solute have less freedom of movement due to intermolecular attractions. Any solution at atmospheric pressure has a negative water potential. There is an inverse relationship of ψ to solute concentration (osmotic potential). Plant cells have a cell wall, which adds another factor affecting osmosis: physical pressure. Increasing the pressure on water increases the ψ. There is direct relationship of ψ to physical pressure. Water potential equation Water potential equals the combine pressure and solute concentration (osmotic potential). ψ = ψp + ψs ψp = Pressure potential ψs = Solute potential; it is proportional to the number of dissolved solute molecules. It is also called osmotic potential. Water potential determines the direction of movement of water. Free water, water that is not bound to solutes or surfaces, moves from the area of higher water potential to the area of lower water potential. Adding solutes always lowers the water potential and the solute potential (ψs) is negative. See and understand the examples in fig 36.8. A plant cell placed in pure water will absorb water and become turgid. Turgor pressure will increase and the cell will push against the rigid wall. The partially elastic wall will push back increasing the ψp until it becomes great enough to offset the tendency of water to enter. A cell placed in a hypertonic solution will become plasmolyzed. There are water channels that allow the flux of water in and out of the cell. These transport proteins are called aquaporins. Aquaporins affect only the rate at which the water flows. They do not affect the concentration gradient or the direction of the water flow. Vacuolated cells have three major compartments: cell wall, cytosol and central vacuole. 1. The plasma membrane regulates transport between the cytosol and cell wall. 2. The membrane that bounds the vacuole is called tonoplast and regulates the traffic between the vacuole and the cytosol. THREE MAJOR PATHWAYS OF TRANSPORT In most plant tissues, the cell wall and the cytosol is continuous from cell to cell. Plasmodesmata are channels that connect the cytosol of adjacent cells. The symplast is continuum of the cytosol connected by the plasmodesmata. The cell walls of adjacent cells are in contact and form the apoplast. Lateral transport in plant tissues can occur via the symplast, the apoplast or through the cell membranes between cells. Bulk flow in long distance transport in the xylem and phloem is due to pressure differences at opposite ends of the xylem and phloem. ABSORPTION OF WATER AND MINERALS THROUGH THE ROOTS. Water and minerals from the soil enter the epidermis of roots, cross the root cortex, pass into the vascular bundle and into the xylem vessels where they move up the plant body. Epidermis → cortex → vascular cylinder → xylem → shoot system Root hairs are the most important avenue of absorption near root tips. The hyphae involved in mycorrhizae are important in the absorption of water and minerals. Symbiotic structures made of fungal hyphae and plant roots. The mycelium of the fungus provides an enormous surface area for absorption. Endodermis, the innermost layer of the cortex, controls mineral uptake into the xylem. Cells have a Casparian strip around the radial and transverse walls, that is impermeable to water and minerals. The Casparian strip is made of suberin. Minerals most pass through carrier proteins in the plasma membrane of the endodermal walls. It requires ATP. Parenchyma cells within the stele discharge minerals into the xylem vessels and tracheids. Vessels and tracheids lack protoplast and consist of tubes made mostly of cellulose. Their cell wall and lumen form part of the apoplast. BULK FLOW DRIVEN BY NEAGATIVE PRESSURE IN THE XYLEM Water and minerals that have been transferred to the xylem are transported upwards in the xylem sap. Plants lose a large amount of water by transpiration, the loss of water vapor from leaves and other aerial parts of the plant. An average maple tree loses an average of 200 liter/hour in the summer time or about 53 gallons/hour. Water lost by transpiration must be replaced by water transported from the roots. 1. Root pressure Roots have many solutes dissolved in their cells, which lowers their water potential in relation to the soil in which they grow. Water moves in from the soil into the roots by osmosis. When soil is very dry, its water potential is very low. Unless the soil is extremely dry, roots have a lower water potential (very negative) than the soil and water tends to move by osmosis from the soil into the roots. Cells in the root pump ions into the root stele. The endodermis prevents these ions from leaking back into the cortex. The water potential in the stele is lowered and water flows in from the root cortex generating a positive pressure that forces fluid up the xylem. This pressure is called root pressure. Root pressure pushes water from the root up the stem. Not strong enough to push the up tall plants. It is very low or non-existent during the summer months. Movement of water is greatest in the summer months when root pressure is the lowest. Guttation is the release of water droplets through small openings on leaves of plants. Guttation is the result of root pressure. Root pressure can force the water up a few meters only. It is not the main mechanism that brings water to the top of the plant but it contributes. Many plants do not generate root pressure at all. 2. Transpiration-Cohesion-Tension Theory Also known as the Transpiration-Cohesion Theory. Transpirational pull Water is constantly being lost through the stomata. This water is replaced with water vapor from the mesophyll cells. 1. On most days, the air is drier than the air in the mesophyll air spaces; the outside air has a lower water potential than the air inside the leaf. 2. Water vapor in the air spaces diffuses down its water potential and out of the leaf through the stomata. 3. As water evaporates, it is replaced by water found in the water film that surrounds the mesophyll cells. Water adheres to the hydrophilic areas of the cellulose microfibrils of the cell wall. 4. As water evaporates, a meniscus is formed in the interphase of the water and air in the air spaces of the mesophyll. This curvature increases the surface tension and the rate of transpiration. 5. Cohesive forces also operate on the surface of the water film. 6. These two forces, adhesion and cohesion, create the meniscus that has a negative force. 7. This negative pressure draws water out of the xylem through the mesophyll, and toward the cells and surface film bordering the air spaces near stomata. See figure 36.14 on page 774. There is a gradient in water potential from the atmosphere down to the soil. The atmosphere has very negative water potential. Leaves have higher water potential than the atmosphere and lose water to it. Stems have higher water potential than the leaves; the roots higher than the stem; and the soil higher than the roots. The gradient creates a pull of the column of water in the xylem due to the hydrogen bonds that exist between the water molecules (cohesion). Adhesion of the water molecules to the xylem walls maintains an unbroken column of water. The walls of the vessels and tracheids are hydrophilic and increase the adhesion of water molecules. The transpirational pull is transmitted from the leaves to the root tips and even into the soil solution. The plant does not spend any of its energy in bringing the water up to the top. Solar energy drives transpiration by causing water to evaporate from the moist walls of mesophyll cells and by maintaining a high humidity in the air spaces with a leaf. CONTROL OF TRANSPIRATION Leaves have a high surface area-to-volume ratio. This ratio facilitates the uptake of CO2 needed in photosynthesis and the release of the waste product, O2. Photosynthesis consumes CO2 and produces O2. Both gases diffuse in and out of the leaf respectively through the stomata. The spongy mesophyll of the leaf increases the surface area exposed to CO2 but also increases the surface area of evaporation. The internal surface area of the leaf may be 10 to 30 times greater than the external surface area. Stomata: major pathway of water loss. A plant loses 95% of the water through the open stomata. The waxy cuticle covering most of the leaf surface prevents evaporation. The stoma is the opening located between two kidney or dumbbell-shaped guard cells. Dumbbell shape in monocots. Kidney shape in dicots. The guard cells are suspended over an air chamber by subsidiary epidermal cells. Guard cells control the diameter of the stoma by changing shape. When the guard cells become turgid, the stoma opens. When flaccid, the stoma closes. Potassium ion mechanism. The changes in turgor pressure in the guard cells are the result of the reversible uptake and loss of K+. Light triggers an influx of K+ into the guard cells. It occurs through active transport; ATP required. A proton pump is probably involved and K+ move through channels driven by a membrane potential. Osmotic pressure decreases and water moves into the guard cells. The increase turgidity of the cells causes a change in shape and the stoma opens. Most of the K+ are stored in the central vacuole. The tonoplast plays a role here. Regulation of aquaporins may also be involved by varying the permeability of the membranes to water. Opening of the stomata is most pronounced in blue light, and to a lesser extent in red light. Light proton pump moves H+ K+ transported into the cell water diffuses into out of the guard cell through specific K channels the guard cells guard cells change shape and open the stoma. The stoma may close by a reversal of the process when light decreases. Loss of turgidity closes the stoma. Stimuli to open and close the stomata: 1. Stomata are open or closed according to the physiological needs of the plant. Photosynthesis: depletion of CO2. A low concentration of CO2 in the leaf induces stomata to open even in the dark. Transpiration: loss of water causes loss of turgor throughout the plant. The hormone abscisic acid is produced in response to water deficiency and causes the guard cells to close. Increase in temperature increases cellular respiration and CO2 production. In mesophytes, the stomata are usually open during the day and closed at night. CAM plants do the reverse. 2. Light triggers the intake of K+ by the guard cells. There are blue-light receptors in the membrane of the guard cells that trigger ATPpowered proton pumps, which in turn promotes the uptake of K+ ions. 3. An internal clock causes the stomata to open and close periodically. Cycles of 24 hours are called circadian rhythms. Xerophytes have leaf adaptations that reduce the rate of transpiration. Small, thick leaves or reduced to spines. Thick cuticle Highly reflective leaves and hairy leaves that trap a boundary of water. Stomata are concentrated on the lower, shady leaf surface, in pits: sunken stomata. CAM pathway of photosynthesis. The stomata open during the night to incorporate CO2 into organic acids. Effects of transpiration on wilting and leaf temperature A leaf may lose more than its weight in water every day. Water may move in the xylem as fast as 75 cm/min, about the speed of a second hand moving around the clock. Water loss is a trade-off for allowing CO2 to enter the leaf. Transpiration also results in evaporative cooling, which can lower the temperature of a leaf by as much as 10-15ºC compared to the surrounding air. The transpiration-to-photosynthesis ratio evaluates how efficiently a plant uses water. It is the amount of water lost per gram of CO2 assimilated into organic material by photosynthesis. For many plants species this ratio is 600:1 or 600 g of water are lost for each gram of CO2 incorporated into carbohydrate. For C4 plants, the ratio is 300:1 Transpiration also brings mineral to all parts of the plant and helps in cooling the plant. Adaptations that reduce evaporative water loss Plants adapted to arid regions are called xerophytes. Completing the life cycle during the brief rainy season. Leaves become very reduced, e.g. spines, to prevent excessive loss of water. Leaf adaptations: thick cuticle, stomata in cavities or crypts, and presence of trichomes. Stem becomes photosynthetic. Fleshy stems and leaves store water. CAM metabolism. SUGARS ARE TRANSPORTED TO PLACES OF STORAGE AND USE The transport of the products of photosynthesis is called translocation. Phloem sap is the aqueous solution that flows through the sieve tubes, and it differs markedly from xylem sap. TRANSLOCATION OF PHLOEM SAP Sucrose is the main sugar translocated in the phloem. Sugars move from the source where it is being produced, to the sink, where the sugars are being utilized or stored. Sucrose manufactured in mesophyll cells can travel via the symplast to sieve-tube members. In some species, sucrose leaves the symplast and travels through the apoplast and is actively incorporated into the sieve-tube members or by the companion cells that then pass the sucrose to the sieve tubes through plasmodesmata. In some plants, companion cells have many ingrowths of their walls, enhancing transfer of solutes between apoplast and symplast. These cells are called transfer cells. In maize and many other plants, phloem loading requires active transport because sucrose concentrations in sieve-tube members are two to three times higher than in mesophyll. The loading is done through proton pumps and cotransport mechanisms. The process of unloading at the sink end varies between the species and organs of the plant. BULK FLOW BY POSITIVE PRESSURE. Pressure flow theory. Phloem sap flows from source to sink at rates as great as 1 m/hr, much too fast to be accounted for by either diffusion or cytoplasmic streaming. This theory postulates that sugar moves in the phloem by means of a pressure gradient that exists between the source, where sugar is loaded into the sieve tube members, and the sink, where sugar is removed from the phloem. 1. Sucrose and other carbohydrates are actively loaded into the sieve tubes at the source by a chemiosmotic mechanism. 2. It requires ATP. ATP supplies energy to pump protons out of the sieve tube members into the apoplast. Creates proton gradient. The gradient drives the uptake of sucrose into the symplast through channels by the cotransport of protons back into the sieve tube members. 3. As a result water moves into the sieve tubes by osmosis increasing the hydrostatic pressure in the sieve tubes that forces water to flow in the sieve tubes. 4. Sugar is actively or passively unloaded from the sieve tube into tissues at the sink. 5. As a result water leaves the sieve tubes at the sink decreasing the hydrostatic pressure inside the sieve tubes. 6. A gradient is created between the source and sinks which drives the flow within the sieve tubes. Other substances transported in the phloem are hormones, ATP, amino acids, inorganic ions, viruses and complex organic molecules like sugar-alcohol compounds. THE SYMPLAST IS HIGHLY DYNAMIC The transport needs of a plant cell change during its development. A developing leaf is a sink but spends most of its life as a source. Water stress may alter the membrane transport proteins. The symplast is living tissue. PLASMODESMATA: CONTINUOUSLY CHANGING STRUCTURES Plasmodesmata can change rapidly in permeability and number. They can open or close rapidly in response to changes in turgor pressure, cytoplasmic calcium levels or cytoplasmic pH. As a leaf matures from sink to a source, its plasmodesmata either close or are eliminated, causing phloem unloading to cease. Plant cells regulate plasmodesmata and cause them to dilate or contract. Viral movement proteins mimic the cell’s regulators of plasmodesmata. When dilated, the plasmodesmata can provide a passageway for macromolecules such RNS and proteins. Certain groups of cells and tissues have many of these plasmodesmata connection. These groups of cells and tissues are called symplastic domains. Electrical signaling in the phloem The phloem can conduct electrical signals similar to those of the animal nervous system through the symplast. These signals help to integrate the functions of the entire plant. The electrical signal triggered in one part of the plant travels in the phloem and elicit a response in other part of the plant. The response can be a change in gene transcription, respiration, photosynthesis, phloem uploading, etc. Phloem: an information superhighway Systemic changes are those that affect the entire body of the plant. Some macromolecules that travel in phloem are proteins, RNA, secondary metabolites, and hormones