cranial nerves
advertisement

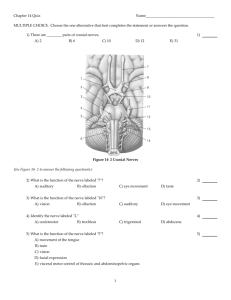
Important developments of the Vertebrates: brain and sense organs The ancestors of vertebrates switched from filter feeding to more active feeding, which required movement and the ability to sense the environment in detail. Important developments of the Vertebrates: brain and sense organs The need to gather and analyze information led to the development of multiple sense organs among the vertebrates. These include complex eyes, pressure receptors, taste and smell receptors, lateral line receptors for detecting water vibrations, and electroreceptors that detect electrical currents. Important developments of the Vertebrates: brain and sense organs The development of sensory structures and increased mobility generated the need for a control center to process information. The anterior end of the nerve cord consequently became enlarged into a brain. Important developments of the Vertebrates: brain and sense organs The vertebrate brain in fact developed into a tripartite brain (with a forebrain, midbrain, and hindbrain) that was enclosed within a protective cranium of bone or cartilage. Human Brain Brain structure In the most primitive forms of brains the forebrain is associated with the sense of smell, the midbrain with vision and the hindbrain with balance and hearing. From this primitive condition the size and complexity of the brain has greatly increased. Hindbrain The Hindbrain has two portions Most posterior portion is the Medulla oblongata. It operates primarily at the reflex level. Reflex centers for respiration, heartbeat, and intestinal movement are found in the medulla oblongata. The medulla oblongata also relays signals from the inner ear and is a major pathway through which signals pass to and from higher areas of the brain. Damage to the medulla, not surprisingly, is lifethreatening. Hindbrain: cerebellum The anterior portion of the hindbrain includes the cerebellum (present only in jawed vertebrates), which is highly folded and convoluted. The cerebellum integrates sensory information (touch, vision, positional, hearing) with motor input to maintain the organism’s equilibrium (its position and equilibrium in relation to gravity). The cerebellum also coordinates motor movements, both reflex movements and directed movements. Hindbrain: cerebellum If the cerebellum is removed an organism’s movements become uncoordinated and uneven. In humans damage to the cerebellum causes a condition called dysmetria in which someone reaching for a target with their hands (or feet) overshoots or undershoots it. Cerebellum The size of the cerebellum is proportional to its role. In fish, the cerebellum is proportionally enlarged in part because it must process lots of input from the lateral line system, but also because fish must orient themselves in three dimensions and equilibrium and balance are thus very important. In bottom-dwelling fish and those that are not active swimmers the cerebellum is relatively small. Midbrain The midbrain develops in step with the eyes and is the part of the brain that receives visual information. The roof of the midbrain (the tectum) is the part that receives visual information (and also lateral line and auditory input). The floor of the midbrain (the tegmentum) initiates motor output based on the input received. The midbrain is often the most prominent portion of the brain in fish and amphibians Frog brain model academic.emporia.edu/sievertl/verstruc/fbrain.htm Forebrain: diencephalon and telencephalon The forebrain has two major parts the posterior diencephalon and the anterior telencephalon. The diencephalon includes the pineal gland, pituitary gland, thalamus and the hypothalamus. Diencephalon: Hypothalamus The hypothalamus plays a major role in homeostasis, the regulation of the body’s internal physiological balance including such aspects as temperature, water balance, appetite, blood pressure and sexual behavior. It achieves this by the release of hormones either produced in the hypothalamus itself or stimulating the release of hormones from the pituitary gland. Diencephalon: Pituitary Gland The posterior pituitary gland stores and releases two hormones produced by the hypothalamus: oxytocin and antidiuretic hormone (ADH). In humans oxytocin stimulates uterine contractions in childbirth and also milk production. ADH acts on the kidneys to increase water retention and reduce urine volume. Diencephalon: Pituitary Gland The anterior pituitary (AP) produces many hormones that in turn control the activity and hormone release from other endocrine glands including the gonads, adrenal glands, and thyroid. Growth hormone is produced in the AP and has a direct effect on growth. Excess production can lead to gigantism and deficient production to dwarfism. Diencephalon: pineal gland and thalamus The pineal gland affects skin pigmentation by affecting melanocytes. It also plays a role in regulating biological rhythms. The thalamus is the major coordinating center for incoming sensory impulses from all over the body and it relays the information to the cerebral cortex. Telencephalon The telencephalon (or cerebrum) includes two expanded lobes, the cerebral hemispheres (which in many mammals are greatly folded) and the olfactory bulbs. The receipt of olfactory information is a major role of the telencephalon and in species in which olfactory information is important the olfactory bulbs are greatly enlarged. Cerebral hemispheres The cerebral hemispheres in reptiles and especially in birds and mammals are enlarged 5 to 20-fold over those of nonamniotes of comparable size. The enlarged size of the cerebral hemispheres allows more and faster processing of sensory information and thus greater intelligence. Cranial nerves Not all nervous inputs and outputs to and from the brain travel via the spinal cord. A series of cranial nerves connect directly into the brain. Most have Roman numerals for names and many of them directly connect to sensory structures and other structures in the head. Cranial nerves The cranial nerves are: Cranial nerve 0: Nervus terminalis runs to blood vessels of the olfactory epithelium. Cranial nerve I: Olfactory nerve: connects with olfactory cells in the mucous membranes of the olfactory sac. Cranial nerve II: Optic nerve: connects to the eyes. Cranial nerves III and IV and VI: connect to extrinsic eye muscles. Cranial nerve V: Trigeminal nerve: branches into three nerves that connect to the eye, jaws and the skin of the head. Cranial nerve VII also innervates the face as well as the taste buds. Cranial nerves Cranial nerve VIII Auditory nerve connects to the inner ear. Cranial nerve IX: Glossopharyngeal nerve connects to taste buds and parts of the throat. Cranial nerve X: Vagus nerve: serves areas of the mouth, pharynx and most of the viscera. Cranial nerve XI: supplies some jaw muscles and the trapezius. Cranial nerve XII: Hypoglossal nerve innervates tongue muscles Cranial nerves arise from both neural crest cells and from ectodermal placodes in the embryo Neural crest cells Neural crest cells are groups of special cells derived from the embryonic dorsal tubular nerve cord. Early in development these cells separate from the neural tube before it closes. They assemble into cords above the neural tube and migrate along distinct pathways to various permanent locations where they differentiate into a variety of structures. Image Source: http://www.niaaa.nih.gov/publications/arh25-3/175-184.htm Neural crest cells Neural crest cells give rise to among other structures: Schwann cells Some components of the peripheral nervous system Odontoblasts (give rise to dentin) Dermis of facial region (from which many skull bones are produced) Beak of birds Some chromatophore cells Connective tissue of the heart Parts of the meninges Ectodermal Placodes Ectodermal placodes (with some exceptions in fish) are thickenings of the surface ectoderm that sink inwards and develop into various sensory structures. Paired olfactory placodes that form at the tip of the head develop into odor receptors that connect to the brain. Paired optic placodes produce the lens of the eye. Ectodermal Placodes: Vestibular apparatus Some dorsolateral placodes (in fish) give rise to the lateral line system. The otic placode (one of the dorsolateral placodes) forms the vestibular apparatus in the inner ear. The vestibular apparatus plays a major role in both balance and hearing. Ectodermal Placodes: Vestibular apparatus There are three semicircular canals (arranged at roughly 90 degree angles to each other) and two connecting structures (the sacculus and utriculus) in the vestibular apparatus The canals are fluid filled and respond to rotation when the head is tilted. The information about orientation and motion is then delivered to the brain for interpretation. http://goodrich.med.harvard.edu/pictures/BRODEL34smaller.bmp Ectodermal Placodes: Vestibular apparatus In some fishes and in reptiles, birds and mammals a section of the vestibular apparatus (the lagena) is specialized for sound reception. In terrestrial vertebrates the lagena is usually elongated and in most mammals it becomes coiled forming the cochlea. Ectodermal Placodes: Vestibular apparatus In mammals sound vibrations are transferred from the eardrum via the inner ear bones (malleus, incus and stapes) to the cochlea. The vibrations cause hair cells in the fluid filled cochlea to move and this movement is converted into nerve signals that are then transmitted to the brain where they are interpreted as sounds. http://www.tchain.com/otoneurology/images/master-ear.jpg Significance of neural crest cells and ectodermal placodes The vertebrate head is mostly a collection of parts that are derived from neural crest or ectodermal placode tissue. These unique tissues and their mode of embryonic production distinguish vertebrates from all other chordates. The role of hox genes in the evolution of the Vertebrates A factor that may have played a role in the evolution of the vertebrates is the duplication of the Hox gene complex. Hox (short for hemeobox) genes are master control genes that regulate the expression of a hierarchy of other genes during development. Hox genes Because a single hox gene influences the expression of many other structural genes a change in when and where a hox gene is turned on may lead to major morphological changes in the phenotype such as the addition or loss of legs, arms, antennae and other structures. http://evolution.berkeley.edu/evolibrary/images/mutantfly.jpg Induced ectopic eyes In Drosopila (arrowed) From Induction of Ectopic Eyes by Targeted Expression of the eyeless Gene in Drosophila Georg Halder,* Patrick Callaerts,* Walter J. Gehring. Science. Vol. 267 24 March 1995 Hox genes Invertebrates and amphioxus have only one set of hox genes, the living jawless vertebrates have two sets, but all jawed vertebrates have four sets. Hox genes The duplication of the Hox genes appears to have occurred around the time vertebrates originated and it may be that this gene duplication freed up copies of these genes, which control development, to generate more complex animals. Hox genes One group of animals in whose evolution hox genes are hypothesized to have played a major role is snakes. It’s suggested that the how genes controlling the expression of the chest region in lizard ancestors of snakes expanded their zone of control in the developing embryo. Hox genes As the hox genes for thoracic development increased their influence, limb development was suppressed at the same time giving the limbless condition we wee in snakes today. Geological Time Scale Precambrian 4,500-542mya Paleozoic: 542-200 mya Mesozoic: 200-65 mya: Age of Dinosaurs Cenozoic: 65mya to present: Age of Mammals Paleozoic Cambrian: 542-488 mya. first appearance of chordates Ordovician: 488-444 mya: Silurian: 444-416 mya Devonian: 416-359 mya Carboniferous: 359-299 mya Permian: 299-251 mya Triassic: 251-200 mya Mesozoic Jurassic: 200-146 mya Cretaceous: 146-65 mya “Camels Often Sit Down Carefully, Perhaps Their Joints Creak” Early vertebrate ancestors Fossils of early chordates are scarce, but a few are known including Pikaia from the Burgess Shale (approx 505 mya) that appears to be an early cephalochordate and has a notochord and segmented muscles. Unlike living cephalochordates it has a pair of sensory tentacles. It was small, about 5cm long. Figure 23.10 15.8 Pikaia Pikaia http://proto5.thinkquest.nl/~jre0294/pikaia%20plaatje.jpg Early vertebrate ancestors Another fossil from China is Haikouella lanceolata about 525mya, which places it in time at the base of the vertebrate radiation and a likely vertebrate ancestor. It was about an inch long (<3cm). Haikouella possesses all the chordate characters and also a suite of vertebrate characters: Dorsal nerve cord with a relatively large brain Gills Head with possible eyes Pharyngeal muscles and gills Myomeres Haikouella lanceolata Haikouella Haikouichthys and Myllokunmingia Two other Chinese fossils from the early Cambrian are clearly early vertebrates. These are Haikouichthys and the very similar (perhaps identical) Myllokunmingia. As in the case of Haikouella, both of these animals were also small (<3cm). Haikouichthys and Myllokunmingia Both Haikouichthys and Myllokunmingia lacked bone and cranial elements, but both possessed Gill bars and gills, V-shaped myomeres, A head A heart Large eyes An ear Possible vertebrae Myellokunmingia Conodonts For almost 150 years tiny, tooth-like microfossils have been important index fossils in geological studies. These conodont elements are extremely common in rocks from the late Cambrian through the end of the Triassic. It was unclear what organism they belonged to until the early 1980’s. Conodont elements http://content.answers.com/main/content/img/McGrawHill/Encyclopedia /images/CE157400FG0010.gif http://www.toyen.uio.no/palmus/galleri/montre/mic01.jpg Conodont (Manticolepis subrecta) elements composed of calcium-phosphate, and are tiny (0.1-0.2mm), toothlike structures from the Devonian Conodonts In the 1980’s the discovery of Carboniferous era conodont fossils in Scotland and later in South Africa solved the mystery. These fossils were of a soft-bodied, slender, laterally compressed animal with a complete set of conodont elements in its pharynx. http://www.le.ac.uk/gl/map2/abstractsetc/conanimals.jpg Conodonts The fossils showed clear evidence that conodonts were vertebrates. There were Vshaped myomeres, a notochord, caudal fin rays, and what appeared to be a postanal tail and a dorsal nerve cord. In addition, histological examination of conodont elements showed they contained a variety of mineralized vertebrate dental tissues: cellular bone, calcium phosphate crystals, calcified cartilage, enamel and dentin. Conodonts Because dentin is laid down by odontoblasts, the presence of dentin in conodont elements is indirect evidence of neural crest tissue, which is a uniquely vertebrate characteristic. Condonts animals were mostly 3-10cm long although some may have been as big as 30cm. Conodont Conodonts There is evidence of wear on conodont elements which suggests they were used to crush and slice food. Recent fossil evidence also shows the conodont elements were attached to tongue-like or cartilaginous plates that could be moved in and out of the mouth presumably to impale and catch food items. This and the animal’s large eyes suggests that conodonts actively selected larger food items and likely were predators. Ostracoderms: Jawless early vertebrates A wide variety of armored jawless fishes collectively referred to as ostracoderms (from the Greek ostrac a shell and derm skin) are known from the very late Cambrian and early Ordovician (488-444 mya) up to near the end of the Devonian period (359 mya). Ostracoderms First vertebrates to possess bone and also the first to possess an intricate lateral line system and an inner ear with two semicircular canals. Ostracoderms were encased in bony plates (with skin in between the plates so they could flex). The bony plates of the head in many cases were large and often fused into a head shield They did not have a well developed endoskeleton and it was usually of cartilage. Given the lack of bony vertebrae in fossils, presumably the body was stiffened by a notochord. Silurian marine fish fauna. Mostly agnathans, but also (#10) a gnathostome; an acanthodian called Nostolepis . www.palaeos.com . Ostracoderms Most ostracoderms were small (10-35 cm in length) and most lacked paired fins so they probably were not precision swimmers. The ostracoderms were jawless with narrow, fixed mouths. They appear to have been mainly filter feeders that used their pharyngeal muscles to pump water. Ostracoderms Because most ostracoderms were small, filter feeders, many were dorsoventrally flattened and most lacked fins it is likely that they were poor swimmers and almost certainly were bottom dwellers that extracted food from sediments. Ostracoderms The phylogenetic relationships of the various ostracoderm groups are still being figured out. Major groups include the Pteraspidomorphs, Osteostracans and Anaspids. Pteraspidomorphs Most Pteraspidomorphs had head shields formed by the fusion of large bony plates. The rest of the body behind the head is covered with small plates and scales. None possessed paired fins, but some had spines that projected from the head shield. Pteraspidomorphs occur from the Ordovician to the late Devonian. They possessed paired nasal openings and a vestibular apparatus with two semicircular canals. (A Pteraspidomorph) (An Anaspid) Anaspids Appear late in the Silurian and possess much more flexible body armour made up of small plates and a hypocercal tail (with an extended ventral lobe) which suggest a trend towards more open-water swimming. Figure 23.14 A Pteraspidomorph 15.10 Ostracoderms (it should read anaspid not anapsid in the caption on the right). Osteostracans Osteostracans were also heavily armoured and possessed a large head shield. Unlike pteraspidomorphs, there were in some species anterior lobes that projected from the head shield (and are now believed to be homologous to the pectoral fins of gnathostomes). These would have enhanced stability in swimming. Osteostracans The Osteostracans are considered to be the closest known relatives of the gnathostomes. Shared derived characters linking them to the gnathostomes include cellular dermal bone, pectoral fins with a narrow base, large orbits and calcified cartilage. Figure 23.14 A Pteraspidomorph 15.10 Ostracoderms (it should read anaspid not anapsid in the caption on the right). http://www.nature.com/nature/journal/v443/n7114/images/443921a-f1.0.jpg Silurian marine fish fauna. Mostly agnathans, but also (#10) a gnathostome; an acanthodian called Nostolepis . www.palaeos.com . Ostracoderms Ultimately, the ostracoderms were outcompeted by fish that possessed the next big evolutionary development: jaws. By the end of the Devonian the ostracoderms had become extinct. The conodonts survived into the Jurassic and a few other agnathans have survived to today.