Chp13
advertisement

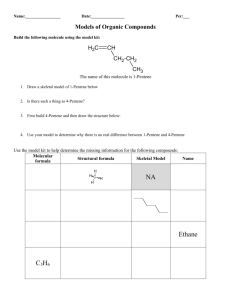
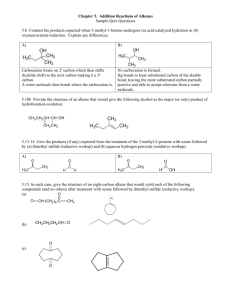
The Organic Chemistry of Enzyme-Catalyzed Reactions Chapter 13 Rearrangements Rearrangements Pericyclic Reactions - concerted reactions in which bonding changes occur via reorganization of electrons within a loop of interacting orbitals Sigmatropic Rearrangements [3,3] sigmatropic rearrangement General form of the Claisen rearrangement O O H Scheme 13.1 Chorismate Mutase-catalyzed Conversion of Chorismate to Prephenate Scheme 13.2 7 O 9 COO1 9 2 6 CH2 3 5 4 HO CH2 C COO- -OOC O 8 COO7 8 1 6 2 5 3 4 HO 13.1 13.2 chorismate prephenate A step in the biosynthesis of Tyr and Phe in bacteria, fungi, plants Conformation of Chorismate in Solution Required conformer for Claisen rearrangement (10-40% observed in solution from NMR spectrum) -O 2C O COO- OH 13.6 chair-like TS‡ Evidence for Chairlike Transition State Stereochemical outcome if chorismate mutase proceeds via chair and boat transition states, respectively, during reaction with (Z)-[9-3H]chorismate -OOC H 3H -OOC O COO- A COOchair OH 13.8 Z-13.7 O B pro-R H COOCOO- 3H COO- H O COO- boat OH OH Scheme 13.3 3H O OH 3H pro-S H Z-13.7 13.9 To Determine the Position of the 3H Chemoenzymatic degradation of the prephenate formed from the chorismate mutase-catalyzed conversion of (Z)-[93H]chorismate to determine the position of the tritium Scheme 13.4 COO- COOHS HR -OOC HS COOO HR pH < 6 - CO2 , -H2O HS O phenylpyruvate tautomerase -HR+ OH Z-[9- 3H]chorismate E-[9- 3H]chorismate 20% 3H release 67% 3H release Therefore, chair TS‡ OH COO- Five Hypothetical Stepwise Mechanisms for the Reaction Catalyzed by Chorismate Mutase COO- (1) + 4 B+ H COO- O rearrangement OH -OOC COOO H2O + -OOC Figure 13.1 COO- O (2) COO- -OOC +O + prephenate COO- O COO- COOB: B+ H O H O COO- COO- (3) prephenate + OH OH COOO COO-O COO- COO- (4) prephenate H O OH X H X H B+ :B 2° inverse isotope effect on C-4 (sp2 sp3); therefore not 1-3 (sp3 sp2) COOO COO-O COO- (5) COOprephenate B: H O B+ O H Both are substrates COO- COO- O COOO COO- OCH3 13.10 13.11 mechanism 5 excluded mechanisms 1, 2, 5 excluded 16 mutants made to show neither general acid-base catalysis (mechanisms 1-3, 5) nor nucleophilic catalysis (mechanism 4) is important Conclusion: pericyclic Function of the enzyme is to stabilize the chair transition state geometry Oxy-Cope Rearrangement General form of Cope (A) and oxy-Cope (B) reactions Cope A OH OH O oxy-Cope B Scheme 13.5 Neither observed yet by an enzyme, but a catalytic antibody has been raised Oxy-Cope Rearrangement Catalyzed by an Antibody COOH ‡ COOH COOH bond rotation OH OH HO 13.14 13.13 COOH COOH COOH bond rotation O 13.15 O Scheme 13.6 OH O N H O OH 13.12 hapten to raise the antibody NH2 [2,3] Sigmatropic Rearrangement Catalyzed by Cyclohexanone Oxygenase O Ph Se 13.16 enzyme NADPH O2 Se Ph [2,3] sigmatropic rearrangement PhSe O H • •O O H PhSe• PhSe H • 13.17 Scheme 13.7 [4+2] Cycloaddition (Diels-Alder) Reaction R ‡ H R R H R' H R' H R' H R" R" R" H 13.18 boat like TS‡ Scheme 13.9 (d,l) An Intramolecular Diels-Alder Reaction Catalyzed by Alternaria solani OCH3 OHC OCH3 OHC O O O O exo endo OHC OCH3 O OHC H O OCH3 O H O H3 C H H3 C H Scheme 13.10 13.19a enzymatic exo : endo is 53 : 47 13.19b solanopyrones in aqueous solution exo : endo is 3 : 97 (nonenzymatic) An Antibody-Catalyzed Diels-Alder Reaction H ‡ O N NHAc -OOC O NH COO- O O Scheme 13.11 O H NH H O N O NHAc N O + O NHAc H O O NH O COO- Hapten used O -OOC H O NH H O 13.20 N O NHAc This hapten gives an antibody that makes only endo product HN O CONMe2 O This hapten gives an antibody that makes only exo product CONMe2 HN O O O O 13.23 N O O 13.24 O N O Rearrangements via a Carbenium Ion An acid-catalyzed acyloin-type rearrangement acid-catalyzed + H2O H :O: R C OH OH C R' R" 13.31 acyloins Scheme 13.14 R C : OH C OH R' R" [1,2] alkyl migration R C R" OH2 + O H C R' R OH O C C R" R' Reactions Catalyzed by Acetohydroxy Acid Isomeroreductase O CH3 C HO OH C COO- + NADP3H CH3 C OH C COO- + NADP+ 13.33 13.32 CH3 C C 14CH 3H 3 14CH 3 O OH COO- + 14CH CH 2 3 13.34 Scheme 13.15 NADP3H CH3 OH OH C C3H COO- 14CH CH 2 3 13.35 + NADP+ OH O CH3 C C COO- CH3 13.36 substrate Kinetically-competent intermediate Proposed Acyloin-type Mechanism for Acetohydroxy Acid Isomeroreductase Scheme 13.16 concerted B H O O C C CH3 :B H COO- CH3 C R + OH OH : O : OH C COO- CH3 C C B: C COO- CH3 C R R NADPH + H+ NADP+ OH OH CH3 C C R H C + COO- C COO- R H HO OH O C CH3 R R CH3 OH OH COO- O+ C OH OH COO- OH OH CH3 intermediate + B H C COO- R stepwise CH3 C NADP+ NADPH + H+ OH O CH3 C R C + COO- C C R H COO- Cyclizations Sterol biosynthesis Conversion of squalene to lanosterol H 20 12 10 1 18O 2 2 NADPH 3 H18O 11 9 14 10 4 5 17 13 8 15 7 H 16 cholesterol 6 13.38 13.37 lanosterol squalene squalene 2,3-epoxidase NADPH O2, flavin, nonheme Fe2+ 2,3oxidosqualenelanosterol cyclase Scheme 13.17 O Initial Mechanism Proposed for 2,3-Oxidosqualene-lanosterol Cyclase 2,3-oxidosqualene-lanosterol cyclase not isolated B: B+ H :O Me 2 Me 1 HO X squalene 2,3-epoxidase Me 9 H 13 17 Me 14 8 H anti-Markovnikov (to get 6-membered ring) 20 Me X H 17 Me 13.40 protosterol Me Me Me HO Scheme 13.18 Me H 3 13.39 squalene Me H Me Me Me H H 7 stereogenic centers (128 possible isomers) Me 13.38 Isotope labeling shows the 4 migrations are intramolecular Covalent catalysis proposed to control stereochemistry lanosterol only isomer formed Me Evidence for 17 Configuration Use of 20-oxa-2,3-oxidosqualene to determine the stereochemistry at C-17 of lanosterol from the reaction catalyzed by 2,3-oxidosqualene-lanosterol cyclase O instead of CH2 B+ H O H Me Me O Me Me O HO 17 Me H 13.41 Me H H 13.42 Scheme 13.19 no covalent catalysis needed Me Me H Me Me O HO 17 Me H Me 13.43 isolated H H 17 Further Support for Structure of Protosterol Use of (20E)-20,21-dehydro-2,3-oxidosqualene to determine the stereochemistry at C-17 of lanosterol from the reaction catalyzed by 2,3-oxidosqualene-lanosterol cyclase B: H extra double bond OH H 3H oxidosqualene cyclase yeast HO O 13.44 Scheme 13.20 20 3H 13.45 20 3H Me Me 17 Me H HO H OH Me H 17 Me H 17 H 13.46 Me Model Study for Stereospecificity and Importance of 17 Configuration Chemical model for the conversion of protosterol to lanosterol Scheme 13.21 BF3 OH H Me B: H H Me H BzO 17 H Me Me Me 13.47 H BF3 H CH2Cl2 -90°C 3 min 90% H Me Me BzO H 13.48 With the 17 isomer a mixture of C-20 epimers is formed 17 Evidence that the Cyclization Is Not Concerted Mechanism proposed for the formation of the minor product isolated in the 2,3oxidosqualene cyclase-catalyzed reaction with 20-oxa-2,3-oxidosqualene O O H X H enzyme + H H X=O HO HO 13.43 40% H+ Markovnikov addition not when X=CH2 X H X=O a b 13.49 3% H H HO HO H O + a H H H 13.41 O H does not come from a concerted reaction H H 13.50 13.51 X = CH2 or O ring expansion b X X + + X=O H HO H HO H 13.52 13.43 H 13.42 Scheme 13.22 Evidence for Carbocation Intermediate R 6 7 O B H 13.53 no reaction without methyls suggests initial epoxide opening Vmax/Km for R = CH3, H, Cl 138, 9.4, 21.9 pmol g-1h-1M-1 correlates with carbocation stabilization (CH3 > Cl >H) Squalene Biosynthesis Squalene synthase-catalyzed conversion of farnesyl diphosphate to squalene via presqualene diphosphate R R H 3-O P O 6 2 -H+ R -PPi R 3-O P O 6 2 NADPH 3-O P O 6 2 H 13.54 farnesyl diphosphate NADP+, PPi R 13.55 presqualene diphosphate R= Scheme 13.23 R 13.37 squalene Rearrangement of Presqualene Diphosphate to Squalene Mechanism proposed for the conversion of presqualene to squalene by squalene synthase R R R H R H H H2C H 3-O P O 6 2 NADP H 13.55 R R= Scheme 13.24 R 13.56 R 13.57 NADP+ H R squalene In the Absence of NADPH there is a Slow Hydrolysis Evidence for 13.56 and 13.57 Mechanisms proposed for the squalene synthasecatalyzed hydrolysis of presqualene diphosphate to several different products in the absence of NADPH H c,d H b H H2C 3-O P O 6 2 b a R 13.56 a H R 13.57 OH d c B: a R= OH B: R 13.55 d c H R R R R R R R R H R HO R OH Scheme 13.25 13.58 14% R 13.59 58% 13.60 24% Support for Intermediate 13.57 Use of dihydro-NADPH to provide evidence for the formation of intermediate 13.57 in the reaction catalyzed by squalene synthase dihydro-NADPH R H H H H B: HO N R R 13.57 R= H NH2 OH R O 13.61 unreactive NADPH to mimic bound NADPH R 13.62 Scheme 13.26 Rearrangements Via Radical Intermediates DNA Photolyase UV light causes DNA damage Reactions catalyzed by DNA photolyase and (6-4) photolyase O O O DNA photolyase HN O NH N N O HN NH h (visible) O P 13.63 cyclobutane pyrimidine dimer h (uv) O N visible h used as a substrate for photoreactivation N O P O O HN O OH N O (6-4) photolyase N O HN h (visible) NH H N P 13.64 (6-4) photoproduct h (uv) both types carcinogenic, mutagenic O N N O P Scheme 13.27 NH2 Other Cofactors Used by Photolyases O CH2O O O- P N O OHO N N O CH2 CH2 N N O P (CHOH)3 N OH O NH N H O H2N N H N 13.65 reduced FADH- H HN N O O N 13.66 N5,N10-methenyl H4PteGlun C CH2OH O H N CHCH2CH2 C COO- (CHOH)3 OH n CH2 HO These act as photoantennae to absorb blue light and transmit to the FADH- N N O NH 13.67 O 8-OH-7,8-didemethyl-5-deazariboflavin Mechanism Proposed for DNA Photolyase H2N N H N h (300-500 nm) H2N H HN O O HN O N HN N R N R 13.66* * R' O N P 13.63 O N O 13.66 NH N H N O N * H N N N H O 13.65* O R' O N NH N H N O NH O 13.65 R' HN NH N O N N O N H P O N NH O 13.68 O HN O O O HN NH N N O O O O HN NH N N O O O NH N N P P P 13.69 13.70 13.71 EPR evidence O Scheme 13.28 Proposed Mechanism for the Formation of the (6-4) Photoproduct O O O H HN O O N H N O N P HN h (uv) [2+2] O HN N O N H P 13.72 Scheme 13.29 H OH O N N O N H P 13.64 O N O HN O O N H :B N O H Mechanism Proposed for (6-4) Photolyase H2N N H N N h (300-500 nm) H HN P 13.64 N R O HN O N H N N O N NH N H O N O NH O 13.65 R' HN O N R' 13.65* O O N N H P 13.72 N R * R' O Scheme 13.30 N O 13.66* 13.66 H O N N H HN N O H2N * H N H N H N O N O N H N NH O P O O HN O O N H N HN O O O H O N N H H N O N O HN O NH N N P P P 13.71 O Coenzyme B12 Rearrangements adenosylcobalamin O H2NC CONH2 H2NC A O N R R CoIII H D H2NC B N N CONH2 OH OH N C a O O CH2 HN N O CONH2 N H O b O O P O H 13.73 N N N NH2 HO -O (coenzyme B12) N OH H2O (vitamin B12) R CH2 cobalamin ring 5-deoxyadenosyl Co 13.74 abbreviation for coenzyme B12 Conversion of Vitamin B12 to Coenzyme B12 Bioynthesis of coenzyme B12 H2O CoIII NADH/FAD NADH/FAD CoII cob(III)alamin reductase cob(II)alamin reductase B12r O O =O P O P O P O 3 OCoI 13.75 Scheme 13.31 B12s O- CH2 O Ad 2nd known reaction at C-5 of ATP -P3O10-5 OH OH adenosylating enzyme Mg2+ R CH2 Co Light Sensitivity of the Co-C Bond of Coenzyme B12 R CH2 Co RCH2 is 5'-deoxyadenosyl Scheme 13.32 h CoII 13.76 R + CH2 13.77 Table 13.1. CoenzymeB12 -Dependent Enzyme-CatalyzedReactions Enzyme Reaction Catalyzed CARBON SKELETAL REARRANGEMENTS CH 3 Methylmalonyl-CoA mutase HOOC HOOCCH 2CH 2 COSCoA CH COSCoA CH 3 2-Methyleneglutarate mutase HOOC HOOCCH 2CH 2 C COOH CH C COOH CH 2 CH 2 CH 3 Glutamate mutase HOOC HOOCCH 2CH 2 CH COOH CH CH COOH NH2 NH2 Isobutyryl-CoA mutase CH 3 H3C CH 3CH 2CH 2 COSCoA CH COSCoA ELIMINATIONS Diol dehydratase R CH CH 2OH OH Glycerol dehydratase RCH 2CHO R = CH 3 or H HOCH 2 CH CH 2OH HOCH 2 CH 2CHO OH Ethanolamine ammonia lyase CH 3CHO CH 2 CH 2OH NH2 ISOMERIZATIONS L-b-Lysine-5 ,6-aminomutase H2C CH 2 CH 2 CHCH 2 COOH NH2 D-Ornithine-4 ,5-aminomutase H3C CH CH 2 CHCH 2 COOH NH2 NH2 H2C CH 2 CH 2 CH COOH NH2 H3C CH NH2 CH 2 CH COOH NH2 NH2 NH2 REDUCTION Ribonucleotide reductase 4- O9 P3O O OH N 4-O P O 9 3 reductant OH O OH N General Form of Coenzyme B12-Dependent Rearrangements H C1 C2 H Y X C1 C2 Y X X is alkyl, acyl, or electronegative group Scheme 13.33 Three Examples of Coenzyme B12 Rearrangements Figure 13.2 A HOOC glutamate mutase H CH2 C CH H H COOH H CH2 C HOOC NH2 CH3 CH C OH H OH diol dehydratase CH2 C NH2 H OH CH3 CH C H O OH -H2O H H C CH NH2 H B COOH CH2 CHCOONH3+ D-ornithine 4,5aminomutase H CH2 CHCH2 CHCOONH2 NH3+ CH3CH2C H Mechanism for Diol Dehydratase and Ethanolamine Ammonia-Lyase Stereospecific conversion of (1R,2R)-[1-2H]-[1-14C]propanediol to (2S)-[2-2H]-[1-14C]propionaldehyde catalyzed by diol dehydratase CH3 HO HO C 14 R H R C CH3 diol dehydratase H C D D 14 C H H O 13.78 13.79 (1R, 2R) (2S) Scheme 13.34 No incorporation of solvent protons; therefore no elimination of water (enol would form) Stereospecific [1,2] migration of the pro-R H with inversion kH/kD = 10-12 Stereospecific Conversion of (1R,2S)-[1-2H]-[1-14C]propanediol to [1-2H]-[1-14C]propionaldehyde Catalyzed by Diol Dehydratase CH3 S H HO C 14 R C CH3 OH diol dehydratase H C H 14 D (1R, 2S) C D O H 13.80 13.81 Scheme 13.35 With the (1R, 2S) epimer, the pro-S H migrates; therefore stereochemistry at C-2 determines which C-1 H migrates Stereospecificity of Elimination of Water Diol dehydratase-catalyzed conversion of (2S)-[1-18O]propanediol to [18O]propionaldehyde (A) and of (2R)-[1-18O]propanediol to propionaldehyde (B) CH3 CH3 H C OH H18O C HR A HS H C H migrates HO C H pro-R HS H18O CH3 - H2O (pro-R hydroxyl group loss) H C C 18O pro-S 13.82 H H 13.83 (2S)-[1-18O] CH3 HO C CH3 H B H18O C HR HS HR migrates H H18O pro-R C C OH - H CH3 H218O (pro-R hydroxyl group loss) H pro-S H C H C O H 13.84 (2R)-[1-18O] Scheme 13.36 The same OH is eliminated (pro-R) regardless of which C-1 H migrates Therefore the C-1 H and the C-2 OH migrate from opposite sides giving inversion at both C-1 and C-2 Crossover Experiment to Show that Diol Dehydratase Catalyzes an Intermolecular Transfer of a Hydrogen from C-1 to C-2 CH3 H HO C C OH H H2C + OH CH3 diol dehydratase H C H 3H + H C 3H HO CH2 3H 13.85 C O C H O Scheme 13.37 Therefore, hydrogen transfer is intermolecular H 1 2 i n C o e n yz m e B R a d io a ct vi it y Time Course for Incorporation of Tritium from [1-3H]propanediol into the Cobalamin of Diol Dehydratase 0 Figure 13.3 30 Time (sec) 60 Determination of the Site of Incorporation of 3H into Coenzyme B12 Aerobic and anaerobic photolytic degradation of coenzyme B12 to locate the position of the tritium incorporated from [1-3H]propanediol in a reaction catalyzed by diol dehydratase OH OH aerobic OH OH h 1/2 3H lost OH O + Co O no 3H here O CH2 N N C N H N 13.86 3H here Co N N NH2 N OH OH NH2 OH anaerobic Scheme 13.38 N all 3H retained O + Co h H2C no 3H here 3H here N N 13.87 N N NH2 Reconstitution of the isolated [3H] coenzyme B12 into apoenzyme with propanediol gives [2-3H]propionaldehyde. All 3H transferred from [3H] coenzyme B12 Synthesized (R,S)-[5-3H] Coenzyme B12 Transfers All 3H to the Product Randomly OH OH O CH3 N N 13.88 N N NH2 possible intermediate to equilibrate the C-5 protons 13.88 isolated with substrates that cannot rearrange Coenzyme B12 is the hydrogen transfer agent. Proposed Rationalization for EPR Spectrum of Co(II) + Carbon Radicals Formation of 5-deoxyadenosine, cob(II)alamin, and substrate radicals during coenzyme B12-dependent reactions R Substrate-H + H C H CoIII Scheme 13.39 R H C H + H CoII Substrate Product Mechanism(s) Proposed for Diol Dehydratase The part shown in the dashed box is even more speculative than the rest of the mechanism Not clear if important Scheme 13.40 OH CH3 R R H CH2 CH2 Co CH CH R OH OH CH3 CH3 13.88 CH CH OH 13.89 OH CH3 R CH3 Co Co CH CH OH Co 13.92 O H C CH2CH3 H2O OH CH3 R HO CH CH2 13.91 CH2 Co OH CH3 R HO CH CH 13.90 CH2 OH CH3 R HO CH CH CH3 Co H Co Radicals observed in EPR spectrum 13.93 Chemical Model Study for a Proposed Diol Dehydratase-catalyzed Rearrangement Involving a Co(III)-olefin -Complex The trapezoid represents the cobaloxime ligand CH2 13CH 2 Co N CH2 Co 13CH 2 OAc MeOH N 13CH CH2 Co N 2 13.96 MeO CH2 13CH 2 Co 13.94 Scheme 13.41 13.95 N 13.97 OMe A Cobalt Complex Is Not Necessary The Fenton reaction as a model for a proposed diol dehydratase-catalyzed free radical rearrangement HO CH2 CH2 Fe2+ + H2O2 HO CH CH2 X Fe3+ + X + + H2O2 + Fe2+ CH3CHO HO HO CH HO CH2 X H+ H CH2 XH -XH Fe+3 O CH HO CH CH3 Scheme 13.42 Fe+2 HO CH CH2 HO CH CH2 HO CH CH2 Another Chemical Model Study for a Proposed Diol Dehydratase-catalyzed Free Radical Rearrangement (the cobalt complex is just to initiate the reaction by radical generation) OH OH OH OH h Co OH OH or N 13.100 H 13.99 OH 13.98 OH O 13.101 OH -H2O H Scheme 13.43 OH H OH OH Carbon Skeletal Rearrangements Stepwise (a) versus concerted (b) mechanisms for the methylmalonyl-CoA mutase-catalyzed generation of 5-deoxyadenosine, cob(II)alamin, and substrate radical Scheme 13.44 H H O O SCoA R H CH2 H COO- R O H R SCoA H CH2 H Co Co Co N N N N H 610His DMB 610His N H DMB H H CH3 COO- b a DMB H N H SCoA H COO- 610His * b EPR confirms Co(II) + organic radical Crystal structures with and without substrates bound show the active site closes upon substrate binding - shields radical intermediates Co-C cleavage is 21 times faster with (CH3)MM-CoA than with (CD3)MM-CoA. Therefore, Co-C and C-H cleavage are concerted. O Six Possible Pathways for the Conversion of MethylmalonylCoA Radical to Succinyl-CoA Radical Catalyzed by Co(I) Methylmalonyl-CoA Mutase Co(II) H H SCoA H H COO13.104 COO- H 13.105 O SCoA O H H a SCoA O H SCoA H H COO- COO- H 13.107 13.106 Co(III) Co(II) Co(I) H Co(III) Co(II) b O H H Co(II) SCoA O H c SCoA H H COO13.102 d O H e H H SCoA COO13.103 H COO13.108 O f O Co(II) 13.110 SCoA H H H COO- SCoA Co(II) H H H COO- 13.109 O Ab initio calculations disfavor pathway e No concensus about the others O H H SCoA H COOCo 13.111 H SCoA H COO- H Co 13,112 Figure 13.4 Ribonucleotide Reductase Converts ribonucleotides to deoxyribonucleotides Results are different from other coenzyme B12 enzymes: • 0.01-0.1% of 3H from [3-3H]UTP is released • no 3H from [3-3H]UTP found in adenosylcobalamin • no crossover between [3-3H]UTP + ATP • [3-3H]UTP gives [3-3H]dUTP • 3H in [5-3H]adenosylcobalamin is washed out in the absence of substrate • adenosylcobalamin 5-deoxyadenosine + Co(II) By EPR formation of Co(II) corresponds to formation of 5-deoxyadenosine and the generation of a thiiyl radical (Cys-408) Mechanism Proposed for Coenzyme B12-dependent Ribonucleotide Reductase C408 S S H CH3Ado CH2Ado • CoII rates of formation are identical; therefore, concerted reaction Co Scheme 13.45 His 4-O P O 9 3 Ha S S S B O Hb Ha O 4-O P O 9 3 Hb • OH B- O OH SH SH H B- OH H S SH -H2O O BH S S S 4-O P O 9 3 Ha O Hb B Hb SH S Ha O Hb B 4-O P O 9 3 B 13.114 13.113 C119 C419 Ha O 4-O P O 9 3 B Ha 4-O P O 9 3 O Hb • HO B- H S 13.117 S B- HO H S S 13.116 H B O H S 13.115 S B Mechanism Proposed for Reducing and Reestablishing the Active Site of Coenzyme B12-dependent Ribonucleotide Reductase C408 C408 S Scheme 13.46 S H H CH2Ado CH2Ado • CoII Co His B- electrons are transferred to active-site disulfide S S C736 SH SH C119 C419 13.117 SH B- SH C731 S C736 regenerates active site for next cycle S C731 reduced by thioredoxin The function of the cobalamin in this enzyme is to initiate the radical reaction by abstraction of H• from Cys-408 Other Ribonucleotide Reductases Use Other Radicals to Abstract a H• from an Active Site Cys Cofactors for class I (13.118), class III (13.119), and class IV (13.120) ribonucleotide reductases O 122Tyr O O 84Asp O H2O H2O O Fe O H3N 238Glu O O O 115Glu 241His S Fe O Fe S Fe S 204Glu H3C S Fe O Mn 13.120 Ade OH OH 13.119 Tyr O S 13.118 Figure 13.5 H NH Fe O 118His CO2 O Mn Reaction Catalyzed by Lysine 2,3-Aminomutase pro-R H H3N NH3+ COO- Ha Hb 13.121 L--Lys SAM [4Fe-4S]+2 PLP Hb COO- H3N H3N pro-R H Ha 13.122 L--Lys Scheme 13.47 Requires PLP, SAM, [4Fe-4S], and a reducing agent Transfers 3-pro-R H of L--Lys to 2-pro-R of L--Lys with migration of 2-amino of L--Lys to C-3 of L--Lys No exchange with solvent With (S)-[5-3H]adenosylmethionine, 3H ends up in both L-Lys and L--Lys One equivalent of Met and 5-deoxyadenosine are formed with L--[3-3H]Lys. 1-6% of 3H ends up in SAM C-S bond is stable, unlike C-Co bond In the presence of a reducing agent, [4Fe-4S]+ is observed in the EPR, which reduces SAM to Met and 5-deoxyadenosyl radical It appears that SAM is functioning like coenzyme B12 Mechanism Proposed for Lysine 2,3-Aminomutase O COOH + H3N =O OH 3PO [4Fe-4S]2+ e- =O H S NH3+ 3PO N H 13.123 Ado Ado CH2 H H H3N H N H H3 N =O N H 13.126 H Scheme 13.48 H COO- NH2 13.122 + PLP + SAM + -OOC [4Fe-4S]+ CH3 H N H N H CH3 OH 3PO OH 3PO CH3 Met [4Fe-4S]2+ H2 O H3N CH3 =O =O 13.127 H N OH 3PO N H 13.124 H3 N COOH CH3 Ado H COO- OH 3PO not observed in EPR CH2 Met [4Fe-4S]2+ CH2 =O OH COO- [4Fe-4S]+ Ado N HR HS CH3 CH2 COOH CH3 H3 N N N H Ado Ado H H3N NH2 13.121 COO- 13.125 N H CH3 unique function for PLP EPR detects organic radicals; 13C label shows product radical 13.126 in EPR spectrum Model Study for New Function of PLP Chemical model study to test the proposed rearrangement mechanism for lysine 2,3aminomutase CH3 CH3 Br Bu3Sn CO2Et N Bu3SnH AIBN CO2Et N Ph Ph - Bu3Sn CH3 CH3 CO2Et N Ph Scheme 13.49 CH3 CO2Et CO2Et N N Ph H Ph SnBu3 To Get Evidence for Substrate Radical (13.124) stabilize -radical COOS H3N 13.128 H NH2 Evidence for Substrate Radical Formation Lysine 2,3-aminomutase-catalyzed rearrangement of 4-thialysine to generate a more stable substrate radical COOH S H3N H S H3 N N HR HS S H3N N H3N H 13.129 Pyr Pyr Pyr 13.130 + S– NH3 EPR detected Ado–CH2 Ado–CH2 COO- PLP + O + H 13.131 COO- HO NH4+ H2 O N Ado–CH3 Ado–CH3 Pyr Pyr = pyridine ring of PLP COO- S N Ado–CH3 Ado–CH2 COO- COO- COO- H HO N COOH OH + N N Pyr B: Pyr Pyr isolated Scheme 13.50