Lecture 4
advertisement

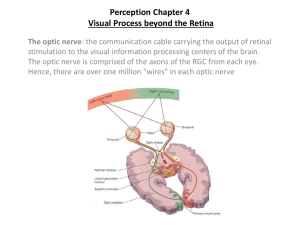
Visual System: Photons to memory -- Each sensory system responds with some specificity to a stimulus and each employs specialized cells - the peripheral receptors - to translate the stimulus into a signal that all neurons can use. -- Precision requires that labor be divided among neurons so that not only different stimulus energies (light vs sound vs mechanical deformation of skin or hair) but also different stimulus qualities (e.g., color vs motion) are analyzed by separate groups of neurons. -- Organization along labeled lines, comparison of events that occur simultaneously at different receptors serves recognition of stimulus strength or contrast. Perception of a sensory experience can change even though the input remains the same. Receptors are specific for a narrow range of input. Sensory transduction - all receptors transduce the energy to which they are sensitive into a change in membrane voltage. Neuronal signaling is accomplished by a combination of rate and temporal codes. Center/surround organizations brought about by lateral inhibition serve to sharpen responses over that which would be achieved by excitation alone. Sekuler, R., and Blake, R., Perception, 2nd edition. New York: McGraw-Hill, Inc., 1990. © 1990 by McGraw-Hill, Inc. Adapted by permission of McGraw-Hill, Inc. W. W. Norton W. W. Norton Visual Field: Monocular and Binocular Zones • • • • • Upper half of visual field projects onto inferior (ventral) half of retina and lower half of visual field projects onto superior (dorsal) half of retina Left visual hemifield projects to nasal hemiretina of left eye and temporal hemiretina of right eye Ganglion cells’ axons from temporal hemiretinas do not cross. Thus nasal retina axons innervate contralateral LGN and temporal retinal axons innervate ipsilateral LGN. Thus the left visual field is processed by right LGN and V1; and right visual field is processed by left LGN and left V1. W. W. Norton Left LGN The LGN from monkey 30 LGN • The LGN consists of six layers. • Each layer receive input from one eye. • Magnocelluar – inner two layers ( 1, 2 layer) – input from M ganglion cells • Pavocelluar – outer four layers ( 3, 4, 5, 6 layer) – input from P ganglion cells • Visual system – divided into two or more streams of information, – different aspects of visual perceptions are processed separately • The division is first evident at the ganglion cells – M class, P class • P cells (80%) - P pathway – selective for wavelength & high spatial frequencies – slow sustained responses • M cells (10 %) - M pathway – sensitive to low spatial frequencies – transient responses LGN • • • • • • P-retinal ganglion cells (abundant in retina; small receptive fields; sensitive to color contrast; and high sensitivity to spatial frequency; thus their pathway will encode form and color) project to the Parvocellular Layers (3, 4, 5, 6) of the LGN. M-retinal ganglion cells (fewer in retina; larger receptive fields; not sensitive to color contrast; lower spatial but higher temporal frequency sensitivity; thus their pathway will encode gross features and movement) project to the Magnocellular Layers (1, 2) of the LGN. Reflective of their inputs, Parvocellular (parvo=small) layers have smaller cells, smaller RFs, are slow to respond, and are color sensitive. Thus form and color are encoded by these layers. Magnocellular Layers (magno=large) have larger cells, large RFs, respond faster, and are not sensitive to color. Thus these layers encode gross features and movement. LGN RFs: Similar concentric RFs to those of ganglion cells, but with stronger (larger) surround. This translates to more sensitivity to contrast than ganglion cells. If you were to lower an electrode straight dorsoventrally from Layer 6 until 1, what would you see? Retinotopy is preserved, so you’d see one part of the retina present in all layers and the orientations in each layer would be in register. If you were to close the right eye and just allow light to hit the one eye, then you would be able to glean which layers are ipsilateral (Layers 2, 3, 5) and contralateral (Layers 1, 4, and 6). Hubel and Wiesel (1977) These recordings are from LGN. LGN neurons have concentric receptive fields with either an on center-off surround or off center-on surround organization. This on centeroff surround cell fires rapidly when the light encompasses the center region, and is inhibited when light is positioned over the surround. The stimulus produces little change when crossing center and surround. These neurons are ideal for signaling changes in illumination from edges. Flow of Visual Information: retina (pretectum; superior colliculus) LGN V1 V2 What/Where Pathways W. W. Norton Zeki, S., A Vision of the Brain. Oxford, UK: Blackwell Scientific, 1993. The primary visual cortex : V1 • The LGN neurons mainly project to the primary visual cortex. • V1 consists of six layers and several sub layers are arranged in bands parallel to the surface of the cortex. • The axons from the LGN terminate on cortical neurons in layer 4 of V1. W. W. Norton These neurons are from V1 and are simple cells. Neural Computation Simple Cells of V1 • • • • • • • For a cell that has separated and elongated on and off regions (simple RF), you need the following effective stimulus: It must excite the specific segment of the retina innervated by receptors in the excitatory zone (specific position on the retina and also a specific (excitatory) position in the RF); It should have the correct linear properties (either bar or edge); It should have a specific axis of orientation; Note: some simple cells are directionally sensitive; and diffuse light is not effective since the firing rate elicited in the on-region cancels that of the off-region. Its on region is made from input from many on-center LGN cells lining up at a particular orientation, and its off region is made from the input of off-center LGN cells lining up at a particular orientation. They tend to be segregated in layers 4 and 6 of V1. Complex Cells of V1 • • • • From a group of simple cells with same axis of orientation but with slightly offset RF positions (but overlapping), you can produce complex cells. Thus for a complex cell, the axis of orientation is still important (responds well to bars or edges), but you no longer are sensitive to the position of that stimulus in the RF (since you no longer have clear on or off regions). Note: some can be direction selective, and some can be end-inhibited or end-stopped. They tend to be all over (outside of layer 4). P layer and M layer in V1 • P layer – The P layer neurons send their axons to neurons in the sub-layer 4C • M layer – The M layer neurons send their information to neurons in sub layer 4C 4B V2 V5. – Cells in layer 4B • selective for the direction of movement • some of these neurons are binocular and sensitive to retinal disparity Splits of P pathway in V1 • The P pathway splits to produce two new path ways in the upper layers of V1. • P-B pathway (blobs) – primarily deal with color • P-I pathway (interblob region) – sensitive to the orientation of the stimulus Columnar Organization to Modules in V1 • • • • Ocular dominance and orientation columns Orientation columns: Identical axis of orientation in all layers Geometric vs. Pinwheel (more likely as measured by voltage-sensitive dyes that flouresce when particular orientation columns light up during presentation of a bar of light with a specific axis of orientation.) A hypercolumn/module (1mm by 1mm by 2mm) consists of an axis of all (0 to 180 degrees) orientations (orientation columns); alternating columns process separate inputs from each eye (ocular dominance columns); and blobs related to color. Visual area 2 • Thick stripes – sensitive to orientation and movement – sensitive to retinal disparity • Thin stripes – not orientation selective – color sensitive • Interstripes – orientation selective – not selective to direction and color Visual area 4 • Both subdivisions of the P pathway, the thick stripes(color) and the inter-stripes(form), project to V4. • Some V4 cells respond not to the wavelength of light but to its ‘color’(color constancy). • Damage to this area of V4 in humans impairs the ability to distinguish color(achromatopsia). • V4 is important for object discrimination. • V4 projects primarily to the temporal visual cortex Visual Areas 3 and 5 • The M pathway projects to V3 and V5. • Cells in V3 – orientation selective – concerned with processing dynamic form • Cells in V5(MT) – process information on motion and stereoscopic depth Fates of Different V1 Layers W. W. Norton Area MT W. W. Norton Zeki, S., A Vision f the Brain. Oxford, UK: Blackwell Scientific, 1993. Zeki, S., A Vision f the Brain. Oxford, UK: Blackwell Scientific, 1993. Different Areas Analyze Different Aspects of A Stimulus