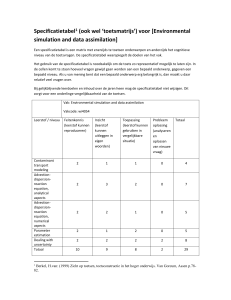
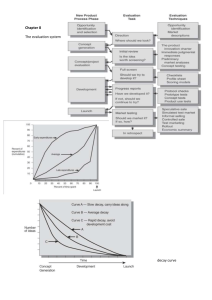
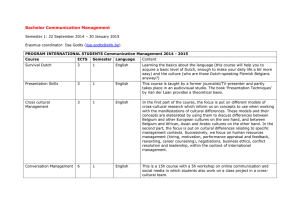
Seksloze wandelende takken gaan al een miljoen
advertisement

PHASMATODAE Stick Insect Phasmida Stick insects are among the best camouflaged of all creatures, with a body shape that mimics the branches of their home. Photograph by Robert Sisson http://animals.nationalgeographic.com/animals/bugs/stick-insect/ Seksloze wandelende takken gaan al een miljoen jaar mee Sommige soorten onder de wandelende takken bestaan al meer dan één miljoen jaar zonder seksuele voortplanting, zo blijkt uit Canadees onderzoek. Het is bekend dat sommige soorten wandelende takken van het geslacht Timema zich voortplanten zonder enige vorm van seks. Deze insecten, die leven aan de Amerikaanse Westkust, produceren genetische klonen van zichzelf. Wetenschappers uit Canada probeerden aan de hand van de mutatiesnelheid van de genen te achterhalen hoe lang deze soorten reeds een seksloos bestaan leiden. Uit dat onderzoek is nu gebleken dat met name de Timema tahoe en de Timema genevieva al minstens één miljoen jaar overleven zonder seks. "Ons onderzoek levert nieuw bewijs dat aseksualiteit niet noodzakelijk resulteert in het snelle verdwijnen van soorten", zo verklaarde vorser Tanja Schwander van de Simon Fraser University in Canada aan de BBC. In het verleden ging men ervan uit dat soorten met een aseksuele reproductie geen lang leven was beschoren. Ze planten zich weliswaar sneller voort, maar zouden evolutionair op een dood spoor zitten. Ze kunnen zich immers minder aanpassen aan veranderende leefomstandigheden. De Timema zijn de oudste insecten waarvan bewijzen bestaan dat ze voor zeer lange periodes seksloos kunnen overleven. Hoe ze dat precies flikken, ondanks die darwinistische onheilstijdingen, moet volgens Schwander verder onderzocht worden. (belga/jv) 20/07/11 http://en.wikipedia.org/wiki/Timema http://bugguide.net/node/view/14905 Order Phasmatodea Family Timematidae Genus Timema Species unknown. Meestervermommer uit ver verleden http://www.geo.uu.nl/ngv/geonieuws/geonieuwsart.php?artikelnr=772 In Eocene afzettingen is een prachtig bewaard gebleven meestervermommer gevonden. Het betreft een fossiel insect dat tot de wandelende bladeren (Phyliinae) moet worden gerekend. Deze groep behoort samen met de wandelende takken tot de orde van de Phasmatodea, waarvan recent zo’n 3000 soorten bekend zijn, vooral uit de tropen en subtropen. Wandelende bladeren beschermen zich tegen hun vijanden door hun gelijkenis met bladeren. Niet voor niets heeft een van de bekendste recente geslachten dan ook de naam Phyllium (= blad). Het nu gevonden exemplaar van 47 miljoen jaar oud heeft met dit geslacht betrekkelijk grote gelijkenis, en is daarom Eophyllium (= vroeg blad) genoemd. Zijn soortnaam, messelensis, is gegeven naar de vindplaats, de groeve Messel in de nabijheid van de Duitse plaats Darmstadt. Eophyllium Photo (A) and line drawing (B) of holotype of fossil leaf insect E. messelensis gen. et sp. nov. from the Eocene Messel Pit, Germany (MeI 12560). a3–a10, abdominal segments 3–10; ant, antennae; cer, cerci; fl, foreleg; fw, forewing; hl, hindleg; hw, hindwing; int, intestinal tract; ml, midleg; vom, vomer. E. messelensis gen. et sp. nov. in evolutionary and biogeographical context. (A) Simplified cladogram with a partial geochronologic scale showing the phylogenetic position of E. messelensis and the temporal sequence of character evolution. Oldest fossil records of determined adult representatives of Timematodea and Euphasmatodea are depicted. M, Messel fossil site; B, Baltic Amber. Dating of splitting events of crown-group Phasmatodea is unknown. Euphasmatodea represent an unknown number of lineages. Figures are not to scale. (B) Distribution of extant and fossil leaf insects. http://scienceblogs.com/pharyngula/2007/01/eophyllium_messelensis.php Het fossiele exemplaar van Eophyllium messelensis (foto Georg Oleschinski). Phyllium celebicum, een recente verwant van het fossiele wandelende blad (foto Georg Oleschinski) Het fossiel dat nu beschreven is, en dat in 2005 werd gevonden, is zeer goed bewaarde gebleven. Het is, samen met soortgelijke fossielen die eerder in dezelfde groeve werden gevonden, veruit het oudste fossiel van een dier dat door zogeheten mimicry (de aanname in kleur en vorm van de natuurlijke omgeving) zijn overlevingskansen vergrootte. Dat was kennelijk een uitstekende tactiek, gezien het feit dat nazaten van tientallen miljoenen later deze tactiek nog steeds (en met succes) toepassen. Tot de tactiek behoort uiteraard ook dat deze dieren zich niet door hun beweging onderscheiden van hun omgeving. Overdag blijven ze daarom bewegingsloos zitten; pas ‘s nachts worden ze actief. Wanneer ze overdag of “s nachts worden gestoord, bewegen ze zich op een manier die op de beweging van een blad in de wind lijkt. Ook dat maakt (en maakte) het moeilijk voor hun belagers om hen als prooi te herkennen. De belagers van Eophyllium messelensis moeten, zoals uit de fossiele fauna daar blijkt, vooral hebben bestaan uit vogels, primitieve primaten en vleermuizen. Op z’n minst een belangrijk deel van die jagers moet bij de jacht op zijn ogen hebben vertrouwd; anders had mimicry immers geen zin. De omgeving van groeve Messel nabij Darmstadt Bij de recente wandelende bladeren zien mannetjes en vrouwtjes er heel verschillend uit, waarbij de mimicry bij de vrouwtjes nog beter is ontwikkeld dan bij de mannetjes. Daaruit zou kunnen worden opgemaakt dat het nu beschreven exemplaar, ondanks zijn duidelijk verbrede lijf, een mannetje moet zijn. De onderzoekers sluiten overigens niet uit dat er in het verleden ook vrouwtjesexemplaren zijn gevonden, die door de vinders als fossiele bladeren zijn beschouwd. De vondst heeft ook paleobiogeografische consequenties, want het verbreidingsgebied van de wandelende bladeren moet in het Eoceen veel groter zijn geweest dan nu. Van wandelende bladeren zijn recent 37 soorten bekend, die alle in zuidoost-Aziê en de direct aangrenzende gebieden leven. Referenties: Wedmann, S., Bradler, S. & Rust, J., 2007. The first fossil leaf insect: 47 million years of specialized cryptic morphology and behavior. Proceedings of the National Academy of Sciences of the United States of America 104, p. 565-569. http://fossilinsects.net/pdfs/wedmann_etal_2007_PNAS_FirstLeafInsect47My.pdf Foto’s welwillend ter beschikking gesteld door Sonja Wedmann, Institut für Paläontologie, Rheinische Friedrich-Wilhelms-Universität, Bonn (Duitsland). http://www.physorg.com/news90157844.html http://discovermagazine.com/2008/jan/first-fossil-of-a-leaf-insectfound/?searchterm=2007%20paleontology http://www.wandelendetakken.be/specials/Eophyllium_messelensis.htm http://en.wikipedia.org/wiki/Eophyllium_messelensis Eophyllium messelensis Een wandelend blad van 47 miljoen jaar oud. Fossielen van wandelende bladeren en wandelende takken zijn extreem zeldzaam. In de Messel groeve nabij de Duitse stad Darmstadt heeft men bij opgravingen in 2005 een perfect bewaard fossiel gevonden van de Eophyllium messelensis. Eophyllium messelensis dankt zijn naam aan de tijd waarin het gevonden is (Eoceen), de genus (Phyllium) en aan de vindplaats (Messel). Mogelijk heeft men in het verleden wel eens fossielen van wandelende bladeren gemist omdat men veronderstelde dat het fossiele bladeren waren. Eerdere fossiele vondsten van de wandelende takken zoals de Bacilllus en de Leptynia worden gedateerd op 23 miljoen jaar oud waarmee de huidige vondst werkelijk een missing link is voor deze Phyllium groep. Mogelijk zijn de eerste wandelende bladeren ontstaan zo’n 125-90 miljoen jaar geleden wanneer de roofdieren hun visuele kenmerken begonnen te gebruiken om te jagen. Het gevonden fossiel betreft een mannelijk wandelend blad dat opmerkelijke vergelijkingen vertoont met de nog levende wandelende bladeren zoals de Phyllium celebicum. Deze vondst toont aan dat de zogeheten mimicry (de aanpassing in kleur en vorm van de natuurlijke omgeving) zijn overlevingskansen vergrootte. Deze tactiek blijkt zeer doeltreffend te zijn gezien het feit dat de nu levende wandelende bladeren, zo’n 47 miljoen jaar later, deze tactiek nog steeds toepassen. De belagers van Eophyllium messelensis moeten, zoals uit de fossiele fauna die ook in de buurt gevonden zijn, vooral hebben bestaan uit vogels, primitieve primaten (oa apen) en vleermuizen, die voornamelijk jagen met hun zicht waardoor de mimicry tot zijn recht komt. De tactiek past in het hele plaatje: zowel kleur als vorm zijn aangepast aan de omgeving, maar ook de dieren zelf blijven overdag onbeweeglijk en wachten op de nacht en wat wind om zich te verplaatsen en te eten. Uit deze vondst blijkt ook dat het verspreidingsgebied van de Phylliinae veel groter was in het eoceen dan nu. Dit komt voornamelijk omdat de temperatuur miljoenen jaren geleden veel hoger was dan nu en de voedselplanten voor de wandelende bladeren ook hier te vinden waren. De meeste wandelende bladeren zijn nu enkel te vinden in zuidoost-Azië en de aangrenzende gebieden. Eerdere fossiele vondsten van de wandelende takken zoals de Bacilllus en de Leptynia Het gevonden fossiel bevindt zich nu in het Senckenberg museum in Frankfurt-am-Main in Duitsland. Foto’s ter beschikking gesteld door Sonja Wedmann, Institut für Paläontologie, Rheinische Friedrich-WilhelmsUniversität, Bonn Bron: www.pnas.org en het artikel dat door Sonja Wedmann ter beschikking gesteld werd. cladogram. Phasmatodea ; ;het fossiel is Europees ;er zijn geen extante inheemse Europese phasmida (bekend ) Wandelende tak kreeg vleugels terug/Weg is niet helemaal weg Jacqueline de Vree http://noorderlicht.vpro.nl/artikelen/10268384/ zie ook --> When Parsimony Goes Wrong: The Wings of Stick Insects The stick insect Sipyloidea sipylus opening its wings. Photo by Drägüs. Timema dorotheae, a member of the basalmost genus of Phasmatodea. Photo by David Maddison. One of the most useful features in characterising insect wings is their venation. A generalised diagram of insect wing venation is given above, but different orders of insects have significantly different wing venation, enough so that relationships can be recognised for fossil insects known from wings alone. Comparisons between wing venation of different orders can also be very useful in establishing their relationships. http://catalogue-of-organisms.blogspot.com/2008/12/when-parsimony-goes-wrong-wings-of.html Links Webpagina's van het laboratorium van Michael Whiting, Brigham Young Universiteit in Provo, Utah Webpagina's over wandelende takken en bladeren van het Australian Museum Online Takkezooi: Nederlandstalige website over het houden en verzorgen van wandelende takken. Mogelijkheid om eitjes te bestellen van verschillende soorten. Tenminste vier keer in de evolutie hebben wandelende takken het vermogen om te vliegen verworven. En voor een groot deel weer verloren, overigens. Een dergelijke herhaling van de evolutie was geheel in strijd met sommige 20 eeuwse opvattingen binnen de evolutietheorie.(1) Weg is weg. Eenmaal uit de stamlijn verdwenen eigenschappen keren nooit meer terug. Dit principe staat bekend als WET VAN DOLLO , begin vorige eeuw opgesteld door de Belgische bioloog en dinosaurusjager Louis Dollo. Zo krijgen kippen nooit meer de tanden terug die ze in hun evolutie van dinosaurus tot pluimvee hebben verloren, en krijgen mensen nooit meer een staart, ons rudimentaire staartbeentje ten spijt. Michael Whiting met een van de 37 verschillende soorten wandelende takken ,gebruikt in zijn onderzoek FIGURE 1. Examples of wing features in stick insects, a, Example of a fully winged (macropterous) female phasmid (Phasma gigas) with enlarged hindwings and thickened forewings. b, Wing venation of male Phyllium celebicum with major veins labelled, demonstrating homology with other insect wing veins. A, anal vein; C, costa vein; Cu, cubitus vein; M, medial vein; R, radius vein; Rs, radial sector vein; Sc, subcosta vein. c, Example of a partially winged (brachypterous) female phasmid (Extatosoma popa) with reduced hindwings. d, Example of a wingless (apterous) female phasmid (Leprocaulinus sp.) with wings entirely absent. Onderzoek van de Amerikaanse bioloog/entomoloog Michael Whiting, verbonden aan de Brigham Young Universiteit in Provo, Utah, tart deze ‘use it or lose it’-(2)opvatting in de hedendaagse biologie. Tenminste vier keer in de evolutie hebben wandelende takken terug functionele vleugels ontwikkeld , zo schrijft hij in een artikel in het tijdschrift Nature. <="" a=""> http://www.newscientist.com/news/news.jsp?id=ns99993269 The graphic is only phasmid (ingroup) taxa. Since Whiting's hypothesis is that the basal phasmid was wingless the basal node carries the wingless state. But the ancestor to the Phasmida is winged (not depicted in the graphic) so the phasmids lost flight (synapomorphy for phasmida on Whiting tree), then regained it four times(red =wing gain ), with two reversals(green=wing loss .) This partial graphic only shows four separate origins of wings and two subsequent losses but no regaining of wings after they were lost Here is are the complete graphics from the article = Character mapping of wing types on phasmid phylogeny Parsimony optimization (ACCTRAN) of winged (blue) and wingless (red) states for male phasmids on the optimization alignment topology. This reconstruction requires seven steps with four wing gains and three losses; DELTRAN optimization requires five wing gains and two losses. Maximum likelihood reconstruction produces similar results (see Supplementary Information). Phylogeny of Phasmatodea on the basis of molecular data. Shown is the single optimization alignment tree based on 18S rDNA, 28S rDNA and histone 3. Nonparametric bootstrap supports are given above each node and partitioned Bremer supports are below each node in the order 18S/28S/H3. This topology is congruent with the maximum likelihood and bayesian topology (see Supplementary Information). Boxes at the end of nodes represent wing character states for males and females, respectively. n. sp., new species; spec. indet., undetermined species. Supplementary Figure 1: This is the ingroup portion of the Bayesian tree, with posterior probabilities given above nodes, and the probability that the ancestral state was wingless given below the node. This reconstruction requires 5 independent wing gains and 2 wing losses. - Download PDF file (61 KB) Supplementary Information: This file provides additional details concerning taxon selection, optimization alignment methodology, incongruence length difference metrics, parsimony tree reconstruction, likelihood tree reconstruction, Bayesian analysis, congruence of molecular results with known morphological characters, parsimony character mapping, and likelihood character mapping. - Download Word file (168 KB) Nooit eerder is een dergelijk staaltje re-evolutie aangetoond, reden voor het tijdschrift om Whitings onderzoek op de cover te zetten. Whiting was er helemaal niet op uit de evolutietheorie de duimschroeven aan te draaien. Hij wilde een moleculaire phylogenetische stamboom maken van de Phasmida ( Phasmatodea / Phasmatodea )gebaseerd op vergelijkend DNA onderzoek tussen de wandelende takken /en de daaraan verwante wandelende bladeren/ Phylliidae , want dat was nog niet eerder gedaan. Voor zijn onderzoek reisde hij de hele wereld af op zoek naar nieuwe exemplaren. Op basis van hun DNA stelde Whiting een evolutionaire stamboom op van 37( voornamelijk soorten uit Nieuw guinea ) verschillende extante soorten Phasmiden. Uit het DNA-onderzoek concludeerde Whiting dat de eerste wandelende tak -Timema knulli - 275 miljoen jaar geleden is ontstaan( volgens de berekeningen met de moleculaire klok ) . Deze " oudste/primitiefste " wandelende tak had tot zijn verbazing geen vleugels. Veertig procent van de wandelende takken gaat gevleugeld door het leven, en de verwachting was dat de oer-tak dus ook vleugels zou moeten hebben. (3) Vijftig tot honderd miljoen jaar later doken er in Whitings vertakkende stamboom wandelende takken op met vleugels. En enige tijd later weer. ..En weer... En weer. Kan niet, volgens de evolutionaire opvattingen. Als een complexe eigenschap eenmaal uit een soort is verdwenen, muteren de erbij horende genen in rap tempo, (4) zodat die eigenschap nooit meer terug kan komen. Weg is weg. Kan wél, zeg Whiting. “Deze insekten zijn miljoenen jaren vleugelloos door het leven gegaan. Maar de genetische blauwdruk ervoor bleef behouden.” (A) Whiting opperde dat de genetische instructies voor het maken van vleugels ook nauw betrokken zijn aan die voor het aanmaken van ledematen zoals de poten . Iets dergelijks is bijvoorbeeld aangetoond bij het fruitvliegje – Drosophila melanogaster. Het is daarom niet verwonderlijk dat de samenwerkende set genen (5)grotendeels ongeschonden (6)miljoenen jaren evolutie heeft doorstaan, schrijft Whiting in Nature. De terugkeer van een verloren(7) gewaande eigenschap zou wel eens een grotere rol kunnen spelen in de evolutie dan tot nu toe werd aangenomen. Whiting vermoed dat iets dergelijks ook het geval is bij kakkerlakken, en bij halfvleugeligen zoals wantsen en schaatsenrijders. Michael F. Whiting et. al.: Loss and recovery of wings in stick insects. In: Nature, vol. 421, p. 264 (16 januari 2003). http://www.tungate.com/whiting.htm http://www.nature.com/nature/journal/v421/n6920/extref/nature01313-s2.doc http://www.ncbi.nlm.nih.gov/pubmed/1252964 (A) Poten regeneratie maakt kreupele vleugels http://www.pubmedcentral.nih.gov/articlerender.fcgi?artid=1634783 http://dbs.umt.edu/research_labs/emlenlab/Lab/Maginnis/Maginnis%20Royal%20Society%202006.pdf Overigens verliezen veel insekten tijdens hun levensloop( of als vertegenwoordigers van een kaste ) hun ( dure ) vleugels --->mieren , bladluizen De( HOX ? ) genen die de vleugelontwikkeling aansturen zijn waarschijnlijk gemakkelijk "aan "of "uitzetbaar " ( sociohormonen ? honger , vitamine-tekorten voedingstoffen /spoorelementen schaarste e.d . Links : classificatie http://tol.tolweb.org/Euphasmida http://zipcodezoo.com/Key/Phasmida_Order.asp http://www.newscientist.com/news/news.jsp?id=ns99993269 http://phasmida.speciesfile.org/. Noten (1)dat werd en wordt , dankbaar aangegrepen door allerlei creationisten http://www.evcforum.net/dm.php?control=msg&t=9438 Ondertussen heeft EVC deze ( ondertussen volledig aanvaarde ) kennis over de evolutie van wandelende takken -vleugels opgenomen in haar archief en geplaats in een bredere evolutiecontext in een afzonderlijk - educatief / documentatie- artikel http://razd.evcforum.net/WingWalkingstick.htm Zie verder over de spartelingen van die GUTOB creationist ----> http://www.volkskrantblog.nl/blog/5720 ( zie de reacties van PeterMudde ) http://www.volkskrantblog.nl/bericht/243296 (creationist ) PB: "De geinformeerde bioloog weet dat re-evolutie van vleugels in phasmieden (wandelden takken) Darwin's theorie volledig falsifieerde." Peter mudde Een( oudbollig ) stukje (door Darwin-volgelingen ) = " Darwin's theorie"(?) of bedoelt is misschien veeleer het idee dat als een complex ontwerp verdwijnt, het niet weer( weggelaten door de meeste creationisten = de moderne nuancering = op dezelfde manier ) verschijnt ? .(= De wet van Dollo ) De idee "eenmaal ( een bepaalde eigenschap ) weg , altijd weg " dateert van voordat de kennis van genetica het huidige nivo bereikte. Nu weten we, dat de aanzet tot de ontwikkeling van iets als vleugels uitgeschakeld kan worden, maar dat de rest van het programma niettemin aanwezig kan blijven. Als het op de een of andere manier weer aangezet kan worden krijg je de situatie zoals bij de Phasmida. Daar is niks antidarwinistisch aan... ATAVISME < (2) ....het principe is afkomstig van JB de Lamarck (3)Volgens de gebruikelijke consensus onder entomologen is het bezit van vleugels een basale en noodzakelijke eigenschap voor het fittere / gemakkelijkere overleven van de orthoptera ... Het stelt deze dieren in staat gemakkelijker te ontsnappen aan predatoren , partners op te zoekn en nieuwe (tijdelijke)voedselbronnen(--> bladluizen levenscyclus/ galwespen ) aan te boren De voorvaderlijke aftakkingen die leide tot de phasmida wordt verondersteld een gevleugeld insect te zijn .... Uitgaande van Whiting & all kan men dus nog steeds de mogelijkheid open houden dat het eerste voorouderlijke lid van de phasmida( niet noodzakelijk T.Knulli ) inderdaad vleugels bezat ... Waarbij T.knulli de meest primitieve extante soort is ( die de vleugels al heeft kwijt gespeeld ) zie ook( A ) The Phylogeny of the Extant Hexapod Orders http://research.amnh.org/scicomp/pdfs/wheeler/Wheeler_etal2001.pdf (4) Genetische erosie : genen waar geen selectiedruk op staat verdwijnen uit de genenpool van de populatie (en de vertikale verspreiding binnen de stamlijn ) (5) HOX genen /HOX GENES LINKS < (6) staat er geen selectie druk(meer) op de vleugels ( =ze ontbrekenof zijn vestigaal ) dan blijft er nog wel conserverende en stabiliserende druk bestaan op poten -bezit (7) " uitgeschakeld "( bijvoorbeeld door een back mutation mechanisme) is niet hetzelfde als daadwerkelijk verlies (= degeneratie ) aan genetische info UPDATES : * University of California - San Diego (2006, January 24). UCSD Biologists Find New Evidence For One-way Evolution. ScienceDaily. Retrieved January 4, 2010, from http://www.sciencedaily.com/releases/2006/01/060123172341.htm Adapted from materials provided by University of California - San Diego. Neutrale mutaties beletten dat evolutie in eiwit omkeert September 2009 Gedane zaken nemen geen keer. Dat geldt ook voor de geschiedenis van het leven op aarde. Voor het eerst is op moleculair niveau bewijs gevonden dat het evolutieproces (op moleculair niveau ) onomkeerbaar is. Het onderzoek staat vandaag in Nature. Evolutie zorgt ervoor dat de biologische diversiteit voortdurend toeneemt. Naarmate de miljoenen jaren verstrijken, ontstaan er nieuwe soorten en gaan organismen steeds minder op elkaar lijken. Die trend gaat er onder meer vanuit dat de tijd niet kan worden omgedraaid. Maar geldt dit ook voor het evolutieproces? Kan bij sommige organismen de evolutie omkeren – door een nieuwe ijstijd, bijvoorbeeld – waardoor een dier of plant opeens weer in een vroeger evolutiestadium terechtkomt? Amerikaanse wetenschappers van de University of Oregon hebben ontdekt dat de(moleculaire) basis voor zo’n omgekeerde evolutie(minstens in dit speciale geval/onderzoek ) ontbreekt. Ze vonden dat mutaties aan een eiwit een welbepaalde functie geven, niet zomaar kunnen worden ‘uitgezet’. Daarvoor zitten andere mutaties, die ontsnappen aan natuurlijke selectie, onvermijdelijk in de weg. De onderzoekers keken naar het receptoreiwit GR dat in de cellen van gewervelde dieren het hormoon cortisol bindt. Dit eiwit stamt af van een oereiwit dat 400 miljoen jaar geleden behalve cortisol ook het hormoon aldosteron kon binden. Het team kon eerder al de zeven mutaties identificeren die samen voor de cortisolvoorkeur hebben gezorgd. Door deze zeven uit te schakelen, wilden ze GR weer in haar vroegere evolutionaire stadium brengen. Het resultaat was verrassend. In de plaats van een overactief eiwit dat zowel cortisol als aldosteron kon binden, zagen ze een inactief, dood eiwit. Blijkbaar hadden andere, nog onbekende en oudere mutaties een succesvolle omkering van de evolutie tegengehouden. Ondanks genetische manipulatie kon de oerfunctie van GR niet meer worden aangezet. De wetenschappers bekeken de atoomstructuur van de gemanipuleerde oereiwitten met behulp van de röntgenstralen van een deeltjesversneller in Chicago. Ze vonden vijf willekeurige mutaties die op een of andere manier samen als een ratelmechanisme werken, zodat het evolutieproces niet kan worden opgekeerd. ‘Stel dat je je bed verzet, en op de vrijgekomen plaats zet je vervolgens je kleerkast. Als je dan je bed terug zou willen zetten, zal je dus eerst ook je kast weer moeten verzetten’, legt Joe Thornton, hoofdauteur van de studie, metaforisch uit. ‘Pas wanneer we deze vijf blokkerende mutaties hadden uitgeschakeld, kon de evolutie worden omgedraaid en kon GR behalve cortisol ook aldosteron binden.’ Maar omdat de vijf ontdekte mutaties geen of weinig effect op de werking van het eiwit hebben, is dit in de natuur uitgesloten. ‘De natuurlijke selectie heeft er geen vat op. Als de oude eiwitfunctie – mét aldosteronbinding – door veranderende omstandigheden opnieuw de meest optimale zou worden, dan nog kan de natuur niet om deze vijf mutaties heen.’ *Een uitgebreide heldere uiteenzetting komt zoals gewoonlijk van http://www.nytimes.com/2009/09/29/science/29evol.html?_r=1 Carl Zimmer Van belang is 1.- Het begin van het artikel: "The Belgian biologist Louis Dollo was the first scientist to ponder reverse evolution. “An organism never returns to its former state,” he declared in 1905, a statement later dubbed Dollo’s law. To see if he was right, biologists have reconstructed evolutionary history. In 2003, for example, a team of scientists studied wings (on the many species )of stick insects. They found that the insects’ common ancestor had wings, but some of its descendants lost them. Later, some of those flightless insects evolved wings again. Yet this study did not necessarily refute Dollo’s law. The stick insects may indeed have evolved a new set of wings, but it is not clear whether this change appeared as reverse evolution at the MOLECULAR level. = Did the insects go back to the EXACT original biochemistry for building wings, or find a NEW route, essentially evolving new proteins? Daarbij moet dus worden onderstreept dat het in het onderzoek van Thornton e.all ,gaat om "teruggedraaide "evolutie op moleculair niveau van een ontwikkelings- stamlijn van een welbepaaldde functie(s)van een eiwit Daaruit sensationele "conclusies " trekken zoals de "mogelijke " evolutie van , bijvoorbeeld , "walvissen die terug kieuwen gaan ontwikkelen" is eigenlijk journalisten -onzin die kan leiden tot stroman-constructies die gemakkelijk passen in de trukendoos van creationisten ... 2.- Maar belangrijker nog is de laatste paragraaf : "....For now it is an open question whether other proteins have an equally hard time evolving backward. But Dr. Thornton suspects they do. “I would never say evolution is never reversible,” Dr. Thornton said. But he thinks it can only go backward when the evolution of the trait is simple, like when a single mutation is involved. When new traits are produced by several mutations that influence one another, he argues, that complexity shuts off reverse evolution. “We know that kind of complexity is very common,” he said.(*zie noot1) If this molecular Dollo’s law holds up, Dr. Thornton believes it says something important about the course of evolutionary history. Natural selection can achieve many things, but it is hemmed in. Even harmless, random mutations can block its path. “The biology (= evolutionary outcome) we ended up with ( in this study )was not inevitable,” he said. “It was just one roll of the evolutionary dice.” Noot 1 ; Bijvoorbeeld : Ook de E.Colli stam in de experimenten van Lenski, die uiteindelijk citraat+ kon doorlaten door zijn moleculaire membraamporieen,gebruikte een opeenvolgende reeks mutaties om die nieuwe eigenschap te verwerven (Pierra) Het is algemeen bekend dat evolutie voortkomt uit meerdere (opeenvolgende) mutaties. In het lab worden binnen kort tijdsbestek voortdurend mutanten gekweekt van virussen, bacterien of celkweken die vaak erg afwijken van het wild-type wat betreft hun DNA-sequentie en eventuele celfuncties. ***Als men het over (on)omkeerbare evolutie wilt hebben kan je eigenlijk niet buiten S.J. GOULD( de rol van de contigency ) en zijn britse tegenpool CONWAY MORRISS ( over "onvermijdelijke "convergente evolutie en de mogelijke teleologische ( en dus ooit voorspelbare ) richting van de evolutieprocessen ? ) Lees vooral het blog van Zimmer en de commentaren die daar ongetwijfeld zullen komen ...: blogs.discovermagazine.com/loo... Belangrijke update : Joe Thornton dient ID-coryfee M. Behe van antwoord : Het is een bijna even doorwegend antwoord als dat van Lenski aan die kerel van conservapedia .... blogs.discovermagazine.com/loo... (Carl Zimmer) en vergeet de website van Thornton niet, met alle relevante pdf documenten www.uoregon.edu/~joet/pubs.htm waaronder het Nature artikel www.uoregon.edu/~joet/PDF/brid... En er is ook nog de website van biochemicus Larry Moran die het ook al over dit onderzoek van Thornton( en over Carl Ziummer ) heeft ... sandwalk.blogspot.com/2009/09/... PIERRA http://www.vkblog.nl/bericht/279265/Terugwaartse_evolutie Terugwaartse evolutie vrijdag 25 september 2009 Een onderzoeksteam aan de Universiteit van Oregon laat zien dat moleculaire evolutie ( in de natuur ) niet teruggedraaid kan worden. Ze onderzochten twee receptors voor cortisol; een voorouderlijke versie van zo’n 400 miljoen jaar geleden en een modernere versie die gedurende de daaropvolgende 40 miljoen jaar evolueerde. De mutaties, die de huidige versie zijn specifieke taak verschaften, werden gevolgd door ‘permissieve’ mutaties, die van geen belang zijn voor de functie van de receptor, maar die wel verhinderden om de huidige receptor zijn voorouderlijke functie terug te geven. Er bestaat veel discussie rond de vraag of evolutie wel dan niet omkeerbaar zou zijn. Er wordt bijvoorbeeld aangenomen dat walvissen en dolfijnen niet meer terug kunnen( op natuurlijke wijze ) naar het gebruik van kiewen; hun evolutionaire weg is te lang geweest om dit mogelijk te maken. Tot nu toe is het nooit mogelijk geweest de (on)omkeerbarheid van de evolutie te toetsen. De onderzoekers bestudeerden daartoe de structuur van een enkel eiwit: de glucocorticoidreceptor (GR) die zich bindt aan het hormoon cortisol, een stresshormoon dat o.a. belangrijk is voor het functioneren van het afweersysteem. GR is een eiwit en dus opgebouwd uit aminozuren. Door de volgorde van deze aminozuren te vergelijken in verschillende huidige diersoorten, zijn de onderzoekers erin geslaagd een ‘stamboom’ van dit eiwit te maken en de voorouderlijke vorm te reconstrueren. Het blijkt dat de GR zo’n 450 miljoen jaar geleden (toen de kraakbeenvissen zoals haaien zich afsplitsten van de beenvissen) zowel affiniteit had voor cortisol als aldosterone, een ander hormoon. Zo’n 40 miljoen jaar later (toen de eerste viervoeters aan land gingen), werd deze receptor specifiek voor cortisol alleen. Gedurende die 40 miljoen jaar zijn er 37 aminozuren veranderd (door mutatie). Slechts twee veranderingen (de X mutaties) waren nodig om de functie te veranderen ofwel om specifiek te worden voor cortisol: de één zorgde dat de proteïne (het eiwit GR) zich vouwde, waardoor het zijn affiniteit voor beide hormonen verloor, en de tweede zorgde ervoor dat de receptor specifiek werd voor cortisol alleen. ( noot A) De onderzoekers vroegen zich af of het mogelijk was de latere vorm van GR terug te brengen in de voorouderlijke vorm waarin het zowel cortisol als aldosterone kan herkennen, door de mutaties van deze twee aminozuren terug te draaien. Ze melden dat het niet mogelijk was de ancestrale functie terug te krijgen en dat bovendien de receptor geen enkel hormoon meer herkende. Het blijkt nu dat er van de 37 mutaties vijf zijn die zich na de twee eerder genoemde (X-mutaties) voordeden; deze waren willekeurig en hadden geen effect op de verandering in de moleculaire functie van de proteïne. Zodra de onderzoekers probeerden de twee hoofdmutaties terug te draaien, bleken deze vijf willekeurige mutaties verantwoordelijk voor het in elkaar storten van de structuur van de proteïne. Van het internet: protein structure Eerst moesten dus de( andere dan de hormoonproducerende ) effecten van deze vijf ( historisch plaats gevonden ) mutaties omgekeerd worden(= back mutations ); Dat kan misschien wel artificieel bewerkstelligd/opgelost worden,; Maar , in de toekomstige natuurlijke loop van de evolutieprocessen (= in de werkelijke(natuurlijke ) wereld )bestaat er geen selectieve druk om deze mutaties terug te draaien; Deze " back- mutations " hebben immers geen (direct) voordeel- effect op welk hormoon de proteïne herkent ....( natuurlijk wél op de "leefbaarheid" / de structuur van het eiwit ) Dergelijke "neutrale" mutaties ( = eventueel aanwezige mutaties waar geen (selectie)druk wordt op uitgeoefend wat betreft de moleculaire hoofdfunctie van het eiwit )spelen geen rol in adaptationistische (schema's aangaande die functies) ... Maar ze " verdwijnen" daarom niet ... *1.- Ze lijken slechts "neutraal "want ze spelen wel degelijk een rol : ze bepalen namelijk of het eiwit structureel nog "levend" is Het gaat er in dit onderoek om dat de 5 neutrale mutaties geen betere of slechtere receptor maken. Ze zijn dus neutraal.( wat dat betreft ) (Pierra ) ....De vijf permissieve ofwel neutrale mutaties (die geen effect hebben op de functie van de receptor) deden zich voor NA de twee essentiele mutaties die de receptor specifiek maakten voor cortisol. -Deze neutrale mutaties zijn niet onderhevig aan de wetten van het uitsluitend adaptationistische evolutiemodel , ofwel het zogenaamde natuurlijke selectie mechanisme op uitsluitend adaptionalistische leest , en vormen daarom een blokkeerpen waardoor het rad van de evolutie niet teruggedraaid kan worden. De selectie vindt dus ook niet plaats op die laatste mutatie(s)!! Dat is nu juist het probleem. wat verder onderzoek stimuleerde Je zou kunnen stellen ( in de trant van een theistische evolutionist)dat deze neutrale, onnuttige mutaties de functie hebben evolutie 'vooruit'te laten gaan en die zouden daarom als teleologisch 'nuttig' gezien kunnen worden Natuurlijke selectie heeft er geen vat op neutrale mutaties : het omdraaien, al was het maar van één van deze vijf mutaties, levert eveneens geen voordeel of nadeel op. Dus mocht dit toevallig toch gebeuren (zie hieronder kansberekening), iets wat heel onwaarschijnlijk is, dan zijn er nog steeds 4 van dat soort mutaties die teruggedraaid moeten worden. Voor elk van deze 4 is de kans opnieuw minimaal dat ze terugmuteren( op natuurlijke wijze ) en dan ook nog eens in de populatie blijven bestaan. * Kansberekening :De kans om twee maal zes te gooien is 1/6x1/6. De kans om een zes en daarna een willekeurig ander getal te gooien is 1/6x5/6. Als je dit doorvoert naar het genoom (stel 1 miljard basen), en je wilt twee keer dezelfde base muteren, dan kom je op ongeveer 1 (de kans om een willekeurige base te muteren) x 1 miljardste om diezelfde base weer te muteren (x1/4 om dezelfde base ervoor terug te krijgen), tegenover 1 miljard -1miljardste / 1 miljard voor een willekeurige mutatie elders in het genoom. Dit natuurlijk afgezien van de muatiesnelheid in het algemeen en dus van de kans op mutatie zelf. Gezien de kans zo laag is kun je je erover verbazen dat een dergelijk onderzoek als dat van Joe Thorton gedaan wordt oftewel nodig is. Dus al met al is de natuurlijke omkering van de vijf permissieve mutaties een zeer onwaarschiinlijke / quasi onmogelijke gebeurtenis. Daarom worden deze 5 mutaties gezien als een pal die voorkomt dat het (natuurlijke ) evolutierad terugdraait. *2.- (Neutrale en andere/zelfs(bijvoorbeeld) homozygotisch schadelijke ) mutaties kunnen evengoed door historisch toeval ( genetische drift /contingency ) non-adaptationistisch worden behouden .... Over adaptationisme en genetische drift ; http://bioinfo.med.utoronto.ca/Evolution_by_Accident/Evolution_by_Accident.html Evolution by Accident v1.43 ©2006 Laurence A. Moran http://bioinfo.med.utoronto.ca/Evolution_by_Accident/Evolution_by_Accident.h tml Modern concepts of evolutionary change are frequently attacked by those who find the notions of randomness, chance, and accident to be highly distasteful. Some of these critics are intelligent design creationists. Their objections have been refuted elsewhere. In this essay I'm more concerned about my fellow evolutionists who go to great lengths to eliminate chance and accident from all discussions about the fundamental causes of evolution. This is my attempt to convince them that evolution is not as predictable as they claim. I was originally stimulated to put my ideas down on paper when I read essays by John Wilkins [Evolution and Chance] and Loren Haarsma [Chance from a Theistic Perspective] on the TalkOrigins Archive. The main conclusion of this essay is that a large part of ongoing evolution is determined by stochastic events that might as well be called "chance" or "random." Furthermore, a good deal of the past history of life on Earth was the product of chance events, or accidents, that could not have been predicted. When I say "evolution by accident" I'm referring to all these events. This phrase is intended solely to distinguish "accidental" evolution from that which is determined by non-random natural selection. I will argue that evolution is fundamentally a random process, although this should not be interpreted to mean that all of evolution is entirely due to chance or accident. The end result of evolution by accident is modern species that do not look designed. Random Genetic Drift Most of us are familiar with natural selection as one of the mechanisms of evolution. Another wellknown mechanism is random genetic drift. This is a mechanism that results in fixation (or loss) of alleles by purely random processes. The topic is thoroughly covered in all the major textbooks—it is not controversial. Unless you deny the existence of random genetic drift, you must agree that some of evolution is entirely due to chance events. The controversy is over how much of evolution is due to drift and how much is due to natural selection. Excellent arguments have been advanced to prove that most of evolution is due to random genetic drift and that's the position I take. Thus, in a discussion about the role of chance and accident in evolution I would say that most of evolution is accidental because of the frequency of drift vs. selection. Note that this says nothing about the perceived importance of these mechanisms. That's a value judgement. Some evolutionists think that adaptation, or evolution by natural selection, is the only interesting part of evolution. These evolutionists don't deny that random genetic drift occurs; instead, they simply relegate it to the category of uninteresting phenomena. Others, like me, think that random genetic drift is far more interesting than natural selection because drift is responsible for junk DNA, molecular phylogenies, molecular clocks, and DNA fingerprinting. Whenever you hear someone denying the role of chance in evolution you can be certain they are ignoring random genetic drift. In some cases this is because they don't even know about drift. Those people are easy to spot because they usually reveal their ignorance of evolution in other ways. In other cases the chance-deniers are well aware of the existence of random genetic drift but they choose to define it out of evolution. Sometimes they specifically say that evolution by drift isn't really evolution. More often, they will use terms like "Darwinism" to describe evolution. Technically, Darwinism can be construed to mean only evolution by natural selection so this is an acceptable way of avoiding the topic of drift. However, if you read closely, you'll see that these writers are often very sloppy about using ""Darwinism" to describe their interests. The term often fills in for all of evolution in a sort of rhetorical sleight of hand. Thus, this group of chance-deniers tends to eliminate chance from evolution by re-defining evolution so that it only applies to natural selection. As you might expect, those who choose to eliminate chance by redefinition are usually the same people that are only interested in natural selection (see above). If adaptationists were being really honest, they would take the time to make their point very clear. I think they should say something like, "If we ignore random genetic drift, for the reasons that we have just given, then evolution is not a random process." Or perhaps they could say, "Much of evolution is truly random but I'm only interested in the part that isn't." The Meaning of Chance Before continuing, we have to address another semantic issue. Philosophers will argue that there is no such thing as chance, randomness, and accident. They will point out, quite correctly, that almost everything has a cause. For example, if the allele for O-type blood became fixed in some native North American populations by random genetic drift—as it did—then this is not really a "random" event. Each and every step in the process had a cause even though it may have been as subtle as a tribe that had a favorable corn crop or a single tuberculosis bacterium that killed off a small child. The net result of millions of caused events was that the A and B alleles were eliminated from the population and everyone has O-type blood. The point about every event having an ultimate cause is true. Taken to its logical conclusion, this is the line of argument that leads to the denial of free will and the triumph of determinism. While it's fun to argue these points over a few beers—and I don't deny that I've engaged in those arguments—it doesn't really impinge on the real world that we are dealing with here. After all, if we are going to deny that anything is random then we have to stop talking about the outcomes of coin flips and the spin of the roulette wheel. But that would be silly. We all know what we mean when we talk about chance events or accidents. We mean that such events are not predictable by any means at our disposal. We are contrasing such events with those, such as natural selection, that have an obvious cause and a (mostly) predictable outcome. Accidents and Contingency Evolution is more than just a change in the frequency of alleles in a population [What Is Evolution?]. It's also the history of life on Earth from the earliest beginnings almost four billion years ago to the present day. When I say that evolution is by accident I'm referring as much to this historical event as to short-term changes within a population. I assume that the opponents of chance are excluding randomness from history as well as from population dyamics but this isn't always clear. There are many random events that took place in the past and these had a profound influence on the outcome of evolution. One of the easiest to understand is mass extinction, although there are many more including some very mundane things like ice ages and continental drift. We don't need to draw up a list in order to make the point. Here's how Freeman and Heron put it in their book "Evolutionary Analysis." Because they are responsible for such thorough turnover in dominant life forms, mass extinctions are an important force in explaining the diversification of life through time. It is doubtful whether many of the major changes observed over the past 250 million years would have taken place had mass extinctions not occurred. ... It is important to recognize, though, that these defining moments were largely random events. Just as mutation and drift introduce a strong random component into the process of adaptation, mass extinctions introduce chance into the process of diversification. This is because mass extinctions are a sampling process analogous to genetic drift. Instead of sampling allele frequencies, mass extinctions sample species and lineages. ... The punchline? Chance plays a large role in the processes responsible for adaptation and diversity. Scott Freeman and Jon C. Herron (1998) p. 520 I'm not sure how the chance-deniers deal with mass extinctions and all the other things that happened. Perhaps they think mass extinctions are really non-random events but they just forget to tell us why? Perhaps they don't think this is part of evolution so they can deny that chance plays a role in evolution by ignoring history? Most likely, the chance-deniers just never think about the significance of the fossil record. The word "contingency" often comes up in these discussions. Contingency is not the same as chance or randomness. Contingency refers to historical events where each step is dependent upon, or contingent upon, preceeding events. The final result of any long historical process is the product of a large number of distinct circumstances any one of which might have been different. If all these circumstances were severely constrained, then the end result might be highly predictable, like the outcome of a computer algorithm. On the other hand, if most of the circumstances were the result of random events, like lucky accidents, then the end result would be unpredictable. When I refer to evolution by accident I'm referring, in part, to a history of evolution that includes a huge number of chance and random events all of which are contingent upon everything that preceeded them. Thus, modern species have taken an unpredicatble path through time. Chance and Necessity Natural selection is an important mechanism of evolution. In negative selection, an unfavorable allele is eliminated from a population because it confers a phenotype that is detrimental to the individual. Individuals carrying this unfavorable allele will tend to have fewer offspring than those with a more beneficial allele. In positive selection, the individuals carrying the beneficial allele will prosper and eventually the beneficial allele will become fixed in the population so that everyone enjoys its phenotypic benefits. New alleles arise because of mutation. If the new allele is detrimental it stands an excellent chance of being eliminated—this is negative, or purifying, selection. If the new allele is beneficial it has a signifcant chance of becoming fixed in the population by positive natural selection. The probablitiy depends on just how beneficial the allele is, or on how fit the individual carrying it is relative to the rest of the population. The probabilty also depends on the size of the population. The numbers have been worked out by population geneticists. It turns out that for large populations (>1000) the size of the population can be ignored and the probability of fixation of any beneficial allele is P = 2s. This means that if a new mutation happens to produce an allele that confers a 5% (s = 0.05) advantage, then there's a 10% (2s = 2 × 0.05 = 0.10) chance that it will be fixed in the population by natural selection. This probability is much higher than the probability that a neutral allele (s = 0) will be fixed by random genetic drift. This is why natural selection is not random. Natural selection is a non-random process because there is a preferred outcome but mutation is, to all intents and purposes, random. This is what Jacques Monod means when he refers to evolution as a combination of chance and necessity. The "chance" is the randomness of mutation and mutations supply the raw material for evolution. The "necesssity" is the non-random process of natural selection. Monod points out that the underlying cause of evolution depends entirely on chance mutations This means that evolution is fundamentally random. In the following passage from his book "Chance and Necessity" he begins by describing the various classes of mutation; substitutions, deletions/insertions, and "scrambling" of chromosomes. He goes on to state clearly what this means for an understanding of evolution. We call these events accidental; we say that they are random occurrences. And since they constitute the only possible source of modifications in the genetic text, itself the sole repository of the organism's hereditary structures, it necessarily follows that chance alone is at the source of every innovation, of all creation in the biosphere. Pure chance, absolutely free but blind, at the very root of the stupendious edifice of evolution: this central concept of modern biology is no longer one among other possible or even conceivable hypotheses. It is today the sole conceivable hypothesis, the only one that squares with observed and tested fact. Jacques Monod (1971) As you can see, Jacques Monod is a strong supporter of evolution by accident. Note that Monod is not even referring to random genetic drift or to accidental events during the history of life. He has focused entirely on the relationship between mutation and natural selection and comes down on the side of "pure chance, absolutely free but blind." He's not the only one who takes this position; the mutationists also favor a prime role for random mutation (see below). The Attack of the Adaptationists Richard Dawkins disagrees with Monod. According to Dawkins, natural selection is the exact opposite of chance. Recall that under positive selection there is a signifcant probabilty that a beneficial allele will become fixed in the population by natural selection. This probabilty (e.g., 10%) is much higher than the probability of fixing a neutral allele by random genetic drift (e.g., 0.001%). It follows that luck and chance are ruled out if you focus your attention on adaptation. The emphasis in all of Dawkins' writing is on evolution by natural selection, or Darwinian theory, or Darwinism. The whole rationale of Darwin's theory was, and is, that adaptive complexity comes about by slow and gradual degrees, step by step, no single step making too large a demand on blind chance as explanation. The Darwinian theory, by rationing chance to the small steps needed to supply variation for selection, provides the only realistic escape from sheer luck as the explanation for life. Richard Dawkins (2004) p. 465 This is the adaptationist perspective with a peculiar Dawkins flavor. Dawkins believes that all living things look designed and the illusion of "design" is due to natural selection. According to Dawkins, evolution cannot be haphazard, random, or accidental, else it could never achieve the design attributed to it. I don't think anyone disagrees with the fact that natural selection is non-random. The disagreement is about whether the observed result is due more to the particular mutations that arose or to the process by which they become fixed in the population. (There is also disagreement about the importance of other mechanisms, such as random genetic drift, but Dawkins ignores that part because he focuses on Darwinism in these discussions. The point he wants to make is that adaptation is not random. It's the only part of evolution that he cares about.) What about Monod's argument that evolution is pure chance because mutations are random? Doesn't this mean that the end result of evolution is largely due to those mutations that just happened to occur? How does Dawkins deal with that? Well, for one thing, he focuses on the particular pathway that was followed and not on all possible pathways that could have been followed. Here's how Dawkins explains it in "The Blind Watchmaker." We have seen that living things are too improbable and too beautifully "designed" to have come into existence by chance. How, then, did they come into existence? The answer, Darwin's answer, is by gradual, step-by-step transformations from simple beginnings, from primordial entities sufficiently simple to have come into existence by chance. Each successive change in the gradual evolutionary process was simple enough, relative to its precursor, to have arisen by chance. But the whole sequence of cumulative steps constitutes anything but a chance process, when you consider the complexity of the final end-product relative to the original starting point. The cumulative process is directed by nonrandom survival. Richard Dawkins (1986) p. 43 One could hardly disagree with this statement as long as you keep your eye on the process and ignore the mutations. Of course natural selection is non-random. That's so trivial that intelligent people wonder why Dawkins spends so much time on it. (It's because some of his readers are Creationists who need to hear it over and over again.) But, does the fact that natural selection is non-random mean that there's no signifcant random component to evolution? No, it doesn't. Dawkins is interested in explaining the "design" he sees in nature. In order to get here from there he reminds us that nonrandom natural selection plays an important role. Fair enough, but what other possible pathways could have been followed? If the actual pathway is only one of several million possibilities then why was that one particular design selected? Is there no possibility that it could have been accidental and selected? Isn't it possible that the actual end-product is as much due to the random mutations that occurred as it is due to natural selection? Dawkins seems to ignore this possibility when he draws attention to the non-randomness of evolution. I think that's a mistake. We can easily agree that natural selection is non-random but it doesn't necessarily follow that evolution now becomes predictable. But perhaps Dawkins does favor some form of predictability? As a matter of fact, he does. He frequently argues that species will become ideally adapted to their environment implying that there is only one way to achieve this goal. He frequently uses metaphors like "Climbing Mt. Improbable" that suggest a single goal for a species. If there's only one mountain in the adaptive landscape, and if the goal is simply to get to the top of that mountain, and if all possible mutations that help achieve that goal are bound to happen, then we can essentially ignore the role of mutation and concentrate on natural selection. Perhaps that's what he's thinking. I dunno. Mutationism Mutationism is one of Dawkins' most feared bogeymen. The term isn't widely used these days because it's tainted by association with saltationist ideas from the middle of the last century. It's time to put aside that bias and think about the modern view of mutationism. Mutationists put even more emphasis on the role of mutation than Jacque Monod did. In this sense they are about as far from the Dawkins position as you can possibly be and this is why modern mutationists are feared by adaptationists. The idea behind mutationism is not difficult to grasp. It's not even radical. The basic concept is that the path of evolution is steered by mutation pressure. One of the modern proponents of this concept is Masatoshi Nei who writes in his book "Molecular Evolutionary Genetics" ... At the DNA level, most new genes seem to have been produced by gene duplication and subsequent nucleotide changes. In these cases, the mutational change of DNA (duplication and nucleotide substitution) is clearly responsible for creating a new gene or character. Natural selection plays no such role. The role of natural selection is to eliminate less fit genotypes and save a beneficial one when there are many different genotypes in the same environment. Therefore, it seems clear that at the molecular level evolution occurs primarily by mutation pressure, though positive selection certainly speeds up gene substition in populations. Masatoshi Nei (1987) p. 413 Mutations are random. If you believe Nei then much of evolutionary biology must be due to chance events. The mutationists claim that mutation is the rate-limiting event in evolution and it's the source of novelty. This contrasts dramatically with those who think that all required mutations will occur when needed and the species will always get to the top of the fitness peak. They don't believe that mutations limit evolution. In addition to their emphasis on the importance of mutation, mutationists are also tolerant of other nonrandom events in evolution. In this book, I have examined various aspects of molecular evolution and concluded that mutation is the driving force of evolution at the molecular level. I have also extended this view to the level of phenotypic evolution and speciation, though I do not deny the importance of natural selection in evolution. I have challenged the prevailing view that a population or organisms contains virtually all sorts of variation and that the only force necessary for a particular character to evolve is natural selection. I have also emphasized the unpredictability of the evolutionary fate of organisms caused by uncontrollable external factors such as rapid climactic changes, geological catastrophes, or even asteroid impacts. Masatoshi Nei (1987) p. 431 You don't need to agree with Nei in order to get the point. The point is that one can't dismiss the role of mutation out-of-hand as the chance-deniers usually do. Those who deny the role of chance need to make their case. It's not good enough to simply declare that natural selection trumps mutation. The Case Against Mutation So, what is the case against chance mutation as a major influence on the evolution of species? I alluded to it earlier. The argument is that mutations do not seriously limit the rate of evolution and, more importantly, they do not direct the course of evolution. In other words, mutations are so common that they will always be available whenever they are needed. The only thing required is that natural selection act on the pool of mutations and this is why natural selection is the most important force in evolution. A corollary to this idea is that modern species are perfectly adapted to their environment and the only thing that will cause further evolution is a change in the environment. This is a necessary corollary since over a period of time all mutations must have been available and must have become fixed in the population. Is this really what Dawkins and his colleagues are saying? Well, not exactly. Mostly they avoid stating flat out that this is their belief but it's pretty clear that it is. How else can they ignore the importance of chance mutations? Here's an example of belief in the perfection of adaptation from "The Blind Watchmaker." If the conditions in which a lineage of animals lives remain constant; say it is dry and hot and has been so without a break for 100 generations, evolution in that lineage is likely to come to a halt, at least as far as adaptations to temperature and humidity are concerned. The animals will become as well fitted as they can to the local conditions. This doesn't mean that they couldn't be completely redesigned to be even better. It does mean that they can't improve themselves by any small (and therefore likely) evolutionary step .... Evolution will come to a standstill until something in the conditions changes: the onset of an ice age, a change in the average rainfall of the area, a shift in the prevailing wind. Richard Dawkins (1986) p.178-179 Do you get the point? Mutations aren't important. When new alleles are required they will be present in the population. That's why species can become perfectly adapted to their local environment over a short period of time. That's why natural selection is the important force and mutations only incidentally are required to supply the raw material for evolution. What Dawkins only implies, Ernst Mayr states explicitly. According to their evolutionary significance, three kinds of mutations can be distinguished: beneficial, neutral, or deleterious. Individuals with genotypes that contain a beneficial new mutation will be faored by natural selection. However, since almost all conceivable beneficial mutations of a population in a stable environment have already been selected in the recent past, the occurence of new beneficial mutations is rather rare. ... Before the role of selection was fully understood, it was believed by many evolutionists that some evolutionary changes were due to "mutation pressure." This is a misconception. The frequency of a gene in a population is in the long run determined by natural selection and stochastic processes, and not by the frequency of mutation. [my emphasis] Ernst Mayr (2001) p.98 With drastic selection taking place in every generation, it is legitimate to ask why evolution is normally so slow. The major reason is that owing to hundreds or thousands of generations that have undegone preceeding selection, a natural population will be close to the optimal genotype. ... All the mutations of which this geneotype is capable and that could lead to an improvement of this standard phenotype have already been incorporated in previous generations. [my emphasis] Ernst Mayr (2001) p.135 If you believe that most species are perfectly adapted to their environment then you will believe that the probablity of a given mutation does not play a major role in evolution by natural selection. Thus, the randomness of mutation isn't a significant feature of evolution but the non-randomness of natural selection is. I don't buy it. I don't see perfection around me and, furthermore, I don't believe that adaptation is all there is to evolution. I think that the path of evolution is influenced by the frequency and randomness of mutations. What Causes Speciation? Speciation plays an important role in evolution. Let's consider branching speciation where a single species splits into two species. This form of speciation usually begins when a small population becomes isolated from the rest of the species. Over time, the two populations diverge because there is no exchange of genes between them. After many generations of isolation they have diverged to the point where they can no longer interbreed when they come back into contact. Two species, or two taxa, have evolved. The events that lead to an isolated population can be entirely accidental. For example, a small flock of birds is blown to the Galapagos by a typhoon or the rising Isthmus of Panama splits a fish species into two groups on the east and west sides. The genetic changes leading to speciation can also be random because they result from fixation of neutral alleles by random genetic drift—this is thought to be the main mechanism of speciation. Thus, speciation—one of the most important events in evolution—is largely by accident. Ernst Mayr, who was one of the world's leading experts on speciation, puts it in plain, simple language. If evolutionists have learned anything from a detailed analysis of evolution, it is the lesson that the origin of new taxa is largely a chance event. Ninety-nine out of 100 newly arising species probably became extinct without giving rise to descendant taxa. And the characteristic of any new taxon is to a large extent determined by such chance factors as the genetic composition of the founding population, the special internal structure of its genotype, and the physical as well as biotic environment that supplies the selection forces of the new species population. Ernst Mayr (1985) Replaying the Tape of Life A the risk of forcing an analogy, it's useful to think of evolution as a tinkerer and not a designer. This is the view originally espressed by François Jacob, winner of the Nobel Prize in 1965 along with Jacques Monod. Jacob points out that the tinkerer takes whatever parts and pieces are lying around and cobbles together something that works. That's what evolution does, whether the mechanism is natural selection, random genetic drift, or anything else. The implications are profound. It means that the modern product of all this tinkering comes from a long line of Rube-Goldberg-like ancestors with no discernable plan or goal. Here's how Jacob puts it, It is hard to realize that the living world as we know it is just one among many possibilities; that its actual structure results from the history of the earth. Yet living organisms are historical structures: literally creations of history. They represent, not a perfect product of engineering, but a patchwork of odd sets pieced together when and where opportunities arose. For the opportunism of natural selection is not simply a matter of indifference to the structure and operation of its products. It reflects the very nature of a historical process, full of contingency. François Jacob (1977) p. 1166 This reminds us that historical processes are contingent and the individual events that occur are often haphazard and unpredictable. The end result—in this case a modern species—is unpredictable because at each step there were many different paths that could have been followed. This is evolution by accident. Stephen Jay Gould wrote a book about the role of chance in evolution. He called it "Wonderful Life." On the surface it's a book about the Burgess Shale and the Cambrian explosion but there's a powerful message as well. Gould is interested in why some species survive while others go extinct. Are the survivors better adapted than the losers of is it a matter of luck? We could answer this question if we could carry out an experiment. I call this experiment "replaying life's tape." You press the rewind button and, making sure you thoroughly erase everything that actually happens, go back to any time and place in the past—say, to the seas of the Burgess Shale. Then let the tape run again and see if the repetition looks at all like the original. If each replay strongly resembles life's actual pathway, then we must conclude that what really happened pretty much had to occur. But suppose that the experimental versions all yield sensible results strikingly different from the actual history of life? What could we then say about the predictability of sefl-conscious intelligence? or of mammals? or of vertebrates? or of life on land? or simply of multicellular persistence for 600 million years? Stephen Jay Gould (1989) pp. 48-50 I'm one of those who think that the tape will be different every time it's replayed because there are so many accidents and contingencies. This is Gould's point as well, recognizing of course that the experiment can never be performed. Others disagree. Simon Conway Morris (2003), for example, claims that something similar to humans will evolve whenever the tape is replayed. And Richard Dawkins (2004) is, as you might imagine, sympathetic to the idea that many similar things will reappear each time. Chance Restored I've tried to summarize all of the random and accidental things that can happen during evolution. Mutations are chance events. Random genetic drift is, of course, random. Accidents and contingency abound in the history of life. All this means that the tape of life will never replay the same way. Chance events affect speciation. All these things seem obvious. So, what's the problem? The "problem" is that writers like Richard Dawkins have made such a big deal about the nonrandomness of natural selection that they risk throwing out the baby with the bathwater. A superficial reading of any Dawkins' book would lead you to the conclusion that evolution is an algorithmic process and that chance and accident have been banished. That's not exactly what he says but it sure is the dominant impression you take away from his work. I said at the beginning of this essay that I was inspired to write it by reading John Wilkins' essay on "Evolution and Chance." Wilkins denies, as does Dawkins, that there is any "deep improbability" in evolution. In fact, his main conclusion is that "evolution is not fundamentally a random process." I beg to differ. I think there's a lot of randomness and improbability in evolution and I hope I've convinced you. I think the term "evolution by accident" is an accurate description of how evolution occurs. http://sandwalk.blogspot.com/2009/09/naked-adaptationism.html http://sandwalk.blogspot.com/2009/09/adaptationist-in-piazza-san-marco.html Gould, S.J. and Lewontin, R.C. (1979) The Spandrels of San Marco and the Panglossian paradigm: a critique of the adaptationist programme. Proc. R. Soc. Lond. B 205:581-598. http://ethomas.web.wesleyan.edu/wescourses/2004s/ees227/01/spa ndrels.html The Spandrels of San Marco and the Panglossian Paradigm: A Critique of the Adaptationist Programme Stephen Jay Gould and Richard C. Lewontin Republished from the original with the kind permission of The Royal Society of London: Gould, S. J. And Lewontin, R. C., "The Spandrels of San Marco and the Panglossian Paradigm: A Critique Of The Adaptationist Programme," Proceedings Of The Royal Society of London, Series B, Vol. 205, No. 1161 (1979), Pp. 581-598. An adaptationist programme has dominated evolutionary thought in England and the United States during the past forty years. It is based on faith in the power of natural selection as an optimizing agent. It proceeds by breaking an organism into unitary "traits" and proposing an adaptive story for each considered separately. Trade-offs among competing selective demands exert the only brake upon perfection; non-optimality is thereby rendered as a result of adaptation as well. We criticize this approach and attempt to reassert a competing notion (long popular in continental Europe) that organisms must be analyzed as integrated wholes, with Baupläne so constrained by phyletic heritage, pathways of development, and general architecture that the constraints themselves become more interesting and more important in delimiting pathways of change than the selective force that may mediate change when it occurs. We fault the adaptationist programme for its failure to distinguish current utility from reasons for origin (male tyrannosaurs may have used their diminutive front legs to titillate female partners, but this will not explain why they got so small); for its unwillingness to consider alternatives to adaptive stories; for its reliance upon plausibility alone as a criterion for accepting speculative tales; and for its failure to consider adequately such competing themes as random fixation of alleles, production of non-adaptive structures by developmental correlation with selected features (allometry, pleiotropy, material compensation, mechanically forced correlation), the separability of adaptation and selection, multiple adaptive peaks, and current utility as an epiphenomenon of nonadaptive structures. We support darwin's own pluralistic approach to identifying the agents of evolutionary change. 1. Introduction The great central dome of St. Mark's Cathedral in Venice presents in its mosaic design a detailed iconography expressing the mainstays of Christian faith. Three circles of figures radiate out from a central image of Christ: angels, disciples, and virtues. Each circle is divided into quadrants, even though the dome itself is radially symmetrical in structure. Each quadrant meets one of the four spandrels in the arches below the dome. Spandrels-the tapering triangular spaces formed by the intersection of two rounded arches at right angles are necessary architectural byproducts of mounting a dome on rounded arches. Each spandrel contains a design admirably fitted into its tapering space. An evangelist sits in the upper part flanked by the heavenly cities. Below, a man representing one of the four biblical rivers (Tigris, Euphrates, Indus, and Nile) pours water from a pitcher in the narrowing space below his feet. The design is so elaborate, harmonious, and purposeful that we are tempted to view it as the starting point of any analysis, as the cause in some sense of the surrounding architecture. But this would invert the proper path of analysis. The system begins with an architectural constraint: the necessary four spandrels and their tapering triangular form. They provide a space in which the mosaicists worked; they set the quadripartite symmetry of the dome above. Such architectural constraints abound, and we find them easy to understand because we do not impose our biological biases upon them. Every fan-vaulted ceiling must have a series of open spaces along the midline of the vault, where the sides of the fans intersect between the pillars. Since the spaces must exist, they are often used for ingenious ornamental effect. In King's College Chapel in Cambridge, for example, the spaces contain bosses alternately embellished with the Tudor rose and portcullis. In a sense, this design represents an "adaptation," but the architectural constraint is clearly primary. The spaces arise as a necessary by-product of fan vaulting; their appropriate use is a secondary effect. Anyone who tried to argue that the structure exists be-cause the alternation of rose and portcullis makes so much sense in a Tudor chapel would be inviting the same ridicule that Voltaire heaped on Dr. Pangloss: "Things cannot be other than they are... Everything is made for the best purpose. Our noses were made to carry spectacles, so we have spectacles. Legs were clearly intended for breeches, and we wear them." Yet evolutionary biologists, in their tendency to focus exclusively on immediate adaptation to local conditions, do tend to ignore architectural constraints and perform just such an inversion of explanation. As a closer example, recently featured in some important biological literature on adaptation, anthropologist Michael Harner has proposed (1977) that Aztec human sacrifice arose as a solution to chronic shortage of meat (limbs of victims were often consumed, but only by people of high status). E. O. Wilson (1978) has used this explanation as a primary illustration of an adaptive, genetic predisposition for carnivory in humans. Harner and Wilson ask us to view an elaborate social system and a complex set of explicit justifications involving myth, symbol, and tradition as mere epiphenomena generated by the Aztecs as an unconscious rationalization masking the "real" reason for it all: need for protein. But Sahlins (1978) has argued that human sacrifice represented just one part of an elaborate cultural fabric that, in its entirety, not only represented the material expression of Aztec cosmology, but also performed such utilitarian functions‚as the maintenance of social ranks‚and systems of tribute among cities. We strongly suspect that Aztec cannibalism was an "adaptation" much like evangelists and rivers in spandrels, or ornamented bosses in ceiling spaces: a secondary epiphnomenon representing a fruitful use of available parts, not a cause of the entire system. To put it crudely: a system developed for other reasons generated an increasing number of fresh bodies; use might as well be made of them. Why invert the whole system in such a curious fashion and view an entire culture as the epi-phenomenon of an unusual way to beef up the meat supply. Spandrels do not exist to house the evangelists. Moreover, as Sahlins argues, it is not even clear that human sacrifice was an adaptation at all. Human cultural practices can be orthogenetic and drive toward extinction in ways that Darwinian processes, based on genetic selection, cannot. Since each new monarch had to outdo his predecessor in even more elaborate and copious sacrifice, the practice was beginning to stretch resources to the breaking point. It would not have been the first time that a human culture did itself in. And, finally, many experts doubt Harner's premise in the first place (Ortiz de Montellano, 1978). They argue that other sources of protein were not in short supply, and that a practice awarding meat only to privileged people who had enough anyway, and who used bodies so inefficiently (only the limbs were consumed, and partially at that) represents a mighty poor way to run a butchery. We deliberately chose non-biological examples in a sequence running from remote to more familiar: architecture to anthropology. We did this because the primacy of architectural constraint and the epiphenomenal nature of adaptation are not obscured by our biological prejudices in these examples. But we trust that the message for biologists will not go unheeded: if these had been biological systems, would we not, by force of habit, have regarded the epiphenomenal adaptation as primary and tried to build the whole structural system from it? 2. The adaptationist programme We wish to question a deeply engrained habit of thinking among students of evolution. We call it the adaptationist programme, or the Panglossian paradigm. It is rooted in a notion popularized by A.R. Wallace and A. Weismann, (but not, as we shall see, by Darwin) toward the end of the nineteenth century: the near omnipotence of natural selection in forging organic design and fashioning the best among possible worlds. This programme regards natural selection as so powerful and the constraints upon it so few that direct production of adaptation through its operation becomes the primary cause of nearly all organic form, function, and behavior. Constraints upon the pervasive power of natural selection are recognized of course (phyletic inertia primarily among them, although immediate architectural constraints, as discussed in the last section, are rarely acknowledged). But they are usually dismissed as unimportant or else, and more frustratingly, simply acknowledged and then not taken to heart and invoked. Studies under the adaptationist programme generally proceed in two steps: (1) An organism is atomized into "traits" and these traits are explained as structures optimally designed by natural selection for their functions. For lack of space, we must omit an extended discussion of the vital issue "What is a trait?" Some evolutionists may regard this as a trivial, or merely a semantic problem. It is not. Organisms are integrated entities, not collections of discrete objects. Evolutionists have often been led astray by inappropriate atomization, as D'Arcy Thompson (1942) loved to point out. Our favorite example involves the human chin (Gould, 1977, pp. 381-382; Lewontin, 1978). If we regard the chin as a "thing," rather than as a product of interaction between two growth fields (alveolar and mandibular), then we are led to an interpretation of its origin (recapitulatory) exactly opposite to the one now generally favored (neotenic). (2) After the failure of part-by-part optimization, interaction is acknowledged via the dictum that an organism cannot optimize each part without imposing expenses on others. The notion of "trade-off' is introduced, and organisms are interpreted as best compromises among competing demands. Thus interaction among parts is retained completely within the adaptationist programme. Any suboptimality of a part is explained as its contribution to the best possible design for the whole. The notion that sub-optimality might represent anything other than the immediate work of natural selection is usually not entertained. As Dr. Pangloss said in explaining to Candide why he suffered from venereal disease: "It is indispensable in this best of worlds. For if Columbus, when visiting the West Indies, had not caught this disease, which poisons the source of generation, which frequently even hinders generation, and is clearly opposed to the great end of Nature, we should have neither chocolate nor cochineal." The adaptationist programme is truly Panglossian. Our world may not be good in an abstract sense, but it is the very best we could have. Each trait plays its part and must be as it is. At this point, some evolutionists will protest that we are caricaturing their view of adaptation. After all, do they not admit genetic drift, allometry, and a variety of reasons for non-adaptive evolution? They do, to be sure, but we make a different point. In natural history, all possible things happen sometimes; you generally do not support your favored phenomenon by declaring rivals impossible in theory. Rather, you acknowledge the rival but circumscribe its domain of action so narrowly that it cannot have any importance in the affairs of nature. Then, you often congratulate yourself for being such an undogmatic and ecumenical chap. We maintain that alternatives to selection for best overall design have generally been relegated to unimportance by this mode of argument. Have we not all heard the catechism about genetic drift: it can only be important in populations so small that they are likely to become extinct before playing any sustained evolutionary role (but see Lande, 1976). The admission of alternatives in principle does not imply their serious consideration in daily practice. We all say that not everything is adaptive; yet, faced with an organism, we tend to break it into parts and tell adaptive stories as if trade-offs among competing, well designed parts were the only constraint upon perfection for each trait. It is an old habit. As Romanes complained about A.R. Wallace in 1900: "Mr. Wallace does not expressly maintain the abstract impossibility of laws and causes other than those of utility and natural selection... Nevertheless, as he nowhere recognizes any other law or cause... he practically concludes that, on inductive or empirical grounds, there is no such other law or cause to be entertained. The adaptationist programme can be traced through common styles of argument. We illustrate just a few; we trust they will be recognized by all: (1) If one adaptive argument fails, try another. Zig-zag commissures of clams and brachiopods, once widely regarded as devices for strengthening the shell, become sieves for restricting particles above a given size (Rudwick, 1964). A suite of external structures (horns, antlers, tusks) once viewed as weapons against predators, become symbols of intra-specific competition among males (Davitashvili, 1961). The eskimo face, once depicted as "cold engineered" (Coon, et al., 1950), becomes an adaptation to generate and withstand large masticatory forces (Shea, 1977). We do not attack these newer interpretations; they may all be right. We do wonder, though, whether the failure of one adaptive explanation should always simply inspire a search for another of the same general form, rather than a consideration of alternatives to the proposition that each part is "for" some specific purpose. (2) If one adaptive argument fails, assume that another must exist; a weaker version of the first argument. Costa & Bisol (1978), for example, hoped to find a correlation between genetic polymorphism and stability of environment in the deep sea, but they failed. They conclude (1978, pp. 132, 133): "The degree of genetic polymorphism found would seem to indicate absence of correlation with the particular environmental factors which characterize the sampled area. The results suggest that the adaptive strategies of organisms belonging to different phyla are different." (3) In the absence of a good adaptive argument in the first place, attribute failure to imperfect understanding of where an organism lives and what it does. This is again an old argument. Consider Wallace on why all details of color and form in land snails must be adaptive, even if different animals seem to inhabit the same environment (1899, p. 148): "The exact proportions of the various species of plants, the numbers of each kind of insect or of bird, the peculiarities of more or less exposure to sunshine or to wind at certain critical epochs, and other slight differences which to us are absolutely immaterial and unrecognizable, may be of the highest significance to these humble creatures, and be quite sufficient to require some slight adjustments of size, form, or color, which natural selection will bring about." (4) Emphasize immediate utility and exclude other attributes of form. Fully half the explanatory information accompanying the full-scale Fiberglass Tyrannosaurus at Boston's Museum of Science reads: "Front legs a puzzle: how Tyrannosaurus used its tiny front legs is a scientific puzzle; they were too short even to reach the mouth. They may have been used to help the animal rise from a lying position." (We purposely choose an example based on public impact of science to show how widely habits of the adaptationist programme extend. We are not using glass beasts as straw men; similar arguments and relative emphases, framed in different words, appear regularly in the professional literature.) We don't doubt that Tyrannosaurus used its diminutive front legs for something. If they had arisen de novo, we would encourage the search for some immediate adaptive reason. But they are, after all, the reduced product of conventionally functional homologues in ancestors (longer limbs of allosaurs, for example). As such, we do not need an explicitly adaptive explanation for the reduction itself. It is likely to be a developmental correlate of allometric fields for relative increase in head and hindlimb size. This non-adaptive hypothesis can be tested by conventional allometric methods (Gould, 1974, in general; Lande, 1978, on limb reduction) and seems to us both more interesting and fruitful than untestable speculations based on secondary utility in the best of possible worlds. One must not confuse the fact that a structure is used in some way (consider again the spandrels, ceiling spaces, and Aztec bodies) with the primary evolutionary reason for its existence and conformation. 3. Telling Stories ‘All this is a manifestation of the rightness of things, since if there is a volcano at Lisbon it could not be anywhere else, for it is impossible for things not to be where they are, because everything is for the best’. [Dr. Pangloss on the great Lisbon earthquake of 1755, in which up to 50,000 people lost their lives]. We would not object so strenuously to the adaptationist programme if its invocation, in any particular case, could lead in principle to its rejection for want of evidence. We might still view it as restrictive and object to its status as an argument of first choice. But if it could be dismissed after failing some explicit test, then alternatives would get their chance. Unfortunately, a common procedure among evolutionists does not allow such definable rejection for two reasons. First, the rejection of one adaptive story usually leads to its replacement by another, rather than to a suspicion that a different kind of explanation might be required. Since the range of adaptive stories is as wide as our minds are fertile, new stones can always be postulated. And if a story is not immediately available, one can always plead temporary ignorance and trust that it will be forthcoming, as did Costa & Bisol (1978), cited above. Secondly, the criteria for acceptance of a story are so loose that many pass without proper confirmation. Often, evolutionists use consistency with natural selection as the sole criterion and consider their work done when they concoct a plausible story. But plausible stories can always be told. The key to historical research lies in devising criteria to identify proper explanations among the substantial set of plausible pathways to any modern result. We have, for example (Gould, 1978) criticized Barash's (1976) work on aggression in mountain bluebirds for this reason. Barash mounted a stuffed male near the nests of two pairs of bluebirds while the male was out foraging. He did this at the same nests on three occasions at ten-day intervals: the first before eggs were laid, the last two afterwards. He then counted aggressive approaches of the returning male toward the model and the female. At time one, aggression was high toward the model and lower toward females but substantial in both nests. Aggression toward the model declined steadily for times two and three and plummeted to near zero toward females. Barash reasoned that this made evolutionary sense, since males would be more sensitive to intruders before eggs were laid than afterward (when they can have some confidence that their genes are inside). Having devised this plausible story, he considered his work as completed (1976, pp. 1099, 1100): The results are consistent with the expectations of evolutionary theory. Thus aggression toward an intruding male (the model) would clearly be especially advantageous early in the breeding season, when territories and nests are normally defended... The initial aggressive response to the mated female is also adaptive in that, given a situation suggesting a high probability of adultery (i.e., the presence of the model near the female) and assuming that replacement females are available, obtaining a new mate would enhance the fitness of males... The decline in malefemale aggressiveness during incubation and fledgling stages could be attributed to the impossibility of being cuckolded after the eggs have been laid... The results are consistent with an evolutionary interpretation. They are indeed consistent, but what about an obvious alternative, dismissed without test by Barash? Male returns at times two and three, approaches the model, tests it a bit, recognizes it as the same phony he saw before, and doesn't bother his female. Why not at least perform the obvious test for this alternative to a conventional adaptive story: expose a male to the model for the first time after the eggs are laid? After we criticized Barash's work, Morton et al. (1978) repeated it, with some variations (including the introduction of a female model), in the closely related eastern bluebird Sialia sialis. "We hoped to confirm", they wrote, that Barash's conclusions represent "a widespread evolutionary reality, at least within the genus Sialia. Unfortunately, we were unable to do so." They found no "anti-cuckoldry" behavior at all: males never approached their females aggressively after testing the model at any nesting stage. Instead, females often approached the male model and, in any case, attacked female models more than males attacked male models. "This violent response resulted in the near destruction of the female model after presentations and its complete demise on the third, as a female flew off with the model's head early in the experiment to lose it for us in the brush" (1978, p. 969). Yet, instead of calling Barash's selected story into question, they merely devise one of their own to render both results in the adaptationist mode. Perhaps, they conjecture, replacement females are scarce in their species and abundant in Barash's. Since Barash's males can replace a potentially "unfaithful" female, they can afford to be choosy and possessive. Eastern bluebird males are stuck with uncommon mates and had best be respectful. They conclude: "If we did not support Barash's suggestion that male bluebirds show anticuckoldry adaptations, we suggest that both studies still had 'results that are consistent with the expectations of evolutionary theory' (Barash 1976, p. 1099), as we presume any careful study would." But what good is a theory that cannot fail in careful study (since by 'evolutionary theory', they clearly mean the action of natural selection applied to particular cases, rather than the fact of transmutation itself)? 4. The Master's Voice Re-examined Since Darwin has attained sainthood (if not divinity) among evolutionary biologists, and since all sides invoke God's allegiance, Darwin has often been depicted as a radical selectionist at heart who invoked other mechanisms only in retreat, and only as a result of his age's own lamented ignorance about the mechanisms of heredity. This view is false. Although Darwin regarded selection as the most important of evolutionary mechanisms (as do we), no argument from opponents angered him more than the common attempt to caricature and trivialize his theory by stating that it relied exclusively upon natural selection. In the last edition of the Origin, he wrote (1872, p. 395): As my conclusions have lately been much misrepresented, and it has been stated that I attribute the modification of species exclusively to natural selection, I may be permitted to remark that in the first edition of this work, and subsequently, I placed in a most conspicuous position-namely at the close of the introduction-the following words: "I am convinced that natural selection has been the main, but not the exclusive means of modification." This has been of no avail. Great is the power of steady misinterpretation. Romanes, whose once famous essay (1900) on Darwin's pluralism versus the panselectionism of Wallace and Weismann deserves a resurrection, noted of this passage (1900, p. 5): "In the whole range of Darwin's writings there cannot be found a passage so strongly worded as this: it presents the only note of bitterness in all the thousands of pages which he has published." Apparently, Romanes did not know the letter Darwin wrote to Nature in 1880, in which he castigated Sir Wyville Thomson for caricaturing his theory as panselectionist (1880, p. 32): I am sorry to find that Sir Wyville Thomson does not understand the principle of natural selection... If he had done so, he could not have written the following sentence in the Introduction to the Voyage of the Challenger: "The character of the abyssal fauna refuses to give the least support to the theory which refers the evolution of species to extreme variation guided only by natural selection." This is a standard of criticism not uncommonly reached by theologians and metaphysicians when they write on scientific subjects, but is something new as coming from a naturalist ... Can Sir Wyville Thomson name any one who has said that the evolution of species depends only on natural selection? As far as concerns myself, I believe that no one has brought forward so many observations on the effects of the use and disuse of parts, as I have done in my "Variation of Animals and Plants under Domestication"; and these observations were made for this special object. I have likewise there adduced a considerable body of facts, showing the direct action of external conditions on organisms. We do not now regard all of Darwin's subsidiary mechanisms as significant or even valid, though many, including direct modification and correlation of growth, are very important. But we should cherish his consistent attitude of pluralism in attempting to explain Nature's complexity. 5. A Partial Typology of Alternatives to the Adaptationist Programme In Darwin's pluralistic spirit, we present an incomplete hierarchy of alternatives to immediate adaptation for the explanation of form, function, and behavior. (1) No adaptation and no selection at all. At present, population geneticists are sharply divided on the question of how much genetic polymorphism within populations and how much of the genetic differences between species is, in fact, the result of natural selection as opposed to purely random factors. Populations are finite in size, and the isolated populations that form the first step in the speciation process are often founded by a very small number of individuals. As a result of this restriction in population size, frequencies of alleles change by genetic drift, a kind of random genetic sampling error. The stochastic process of change in gene frequency by random genetic drift, including the very strong sampling process that goes on when a new isolated population is formed from a few immigrants, has several important consequences. First, populations and species will become genetically differentiated, and even fixed for different alleles at a locus in the complete absence of any selective force at all. Secondly, alleles can become fixed in a population in spite of natural selection. Even if an allele is favored by natural selection, some proportion of populations, depending upon the product of population size N and selection intensity s, will become homozygous for the less fit allele because of genetic drift. If N*s is large, this random fixation for unfavorable alleles is a rare phenomenon, but if selection coefficients are on the order of the reciprocal of population size (N*s = 1) or smaller, fixation for deleterious alleles is common. if many genes are involved in influencing a metric character like shape, metabolism, or behavior, then the intensity of selection on each locus will be small and N*s per locus may be small. as a result, many of the loci may be fixed for non-optimal alleles. Thirdly, new mutations have a small chance of being incorporated into a population, even when selectively favored. Genetic drift causes the immediate loss of most new mutations after their introduction. With a selection intensity s, a new favorable mutation has a probability of only 2s of ever being incorporated. Thus one cannot claim that, eventually, a new mutation of just the right sort for some adaptive argument will occur and spread. "Eventually" becomes a very long time if only one in 1,000 or one in 10,000 of the "right" mutations that do occur ever get incorporated in a population. (2) No adaptation and no selection on the part at issue; form of the part is a correlated consequence of selection directed elsewhere. Under this important category, Darwin ranked his "mysterious" laws of the "correlation of growth." Today, we speak of pleiotropy, allometry, "material compensation"‚(Rensch, 1959, pp. 179-187) and‚ mechanically forced correlations in D'Arcy Thompson's sense (1942; Gould 1971). Here we come face to face with organisms as integrated wholes, fundamentally not decomposable into independent and separately optimized parts. Although allometric patterns are as subject to selection as static morphology itself (Gould, 1966), some regularities in relative growth are probably not under immediate adaptive control. For example, we do not doubt that the famous 0.66 interspecific allometry of brain size in all major vertebrate groups represents a selected "design criterion," though its significance remains elusive (Jerison, 1973). It is too repeatable across too wide a taxonomic range to represent much else than a series of creatures similarly well designed for their different sizes. But another common allometry, the 0.2 to 0.4 intraspecific scaling among homeothermic adults differing in body size, or among races within a species, probably does not require a selectionist story, though many, including one of us, have tried to provide one (Gould, 1974). R. Lande (personal communication) has used the experiments of Falconer (1973) to show that selection upon body size alone yields a brain-body slope across generations of 0.35 in mice. More compelling examples abound in the literature on selection for altering the timing of maturation (Gould, 1977). At least three times in the evolution of arthropods (mites, flies, and beetles), the same complex adaptation has evolved, apparently for rapid turnover of generations in strongly r-selected feeders on super-abundant but ephemeral fungal resources: females reproduce as larvae and grow the next generation within their bodies. Offspring eat their mother from inside and emerge from her hollow shell, only to be devoured a few days later by their own progeny. It would be foolish to seek adaptive significance in paedomorphic morphology per se; it is primarily a byproduct of selection for rapid cycling of generations. In more interesting cases, selection for small size (as in animals of the interstitial fauna) or rapid maturation (dwarf males of many crustaceans) has occurred by progenesis (Gould, 1977, pp. 324-336), and descendant adults contain a mixture of ancestral juvenile and adult features. Many biologists have been tempted to find primary adaptive meaning for the mixture, but it probably arises as a by-product of truncated maturation, leaving some features "behind" in the larval state, while allowing others, more strongly correlated with sexual maturation, to retain the adult configuration of ancestors. (3) The de-coupling of selection and adaptation. (i) Selection without adaptation. Lewontin (1979) has presented the following hypothetical example: "A mutation which doubles the fecundity of individuals will sweep through a population rapidly. If there has been no change in efficiency of resource utilization, the individuals will leave no more offspring than before, but simply lay twice as many eggs, the excess dying because of resource limitation. In what sense are the individuals or the population as a whole better adapted than before? Indeed, if a predator on immature stages is led to switch to the species now that immatures are more plentiful, the population size may actually decrease as a consequence, yet natural selection at all times will favour individuals with higher fecundity." (ii) Adaptation without selection. Many sedentary marine organisms, sponges and corals in particular, are well adapted to the flow regimes in which they live. A wide spectrum of "good design" may be purely phenotypic in origin, largely induced by the current itself. (We may be sure of this in numerous cases, when genetically identical individuals of a colony assume different shapes in different microhabitats.) Larger patterns of geographic variation are often adaptive and purely phenotypic as well. Sweeney and Vannote (1978), for example, showed that many hemi-metabolous aquatic insects reach smaller adult size with reduced fecundity when they grow at temperatures above and below their optima. Coherent, climatically correlated patterns in geographic distribution for these insects-so often taken as a priori signs of genetic adaptation may simply reflect this phenotypic plasticity. "Adaptation" -- the good fit of organisms to their environment -- can occur at three hierarchical levels with different causes. It is unfortunate that our language has focused on the common result and called all three phenomena "adaptation": the differences in process have been obscured, and evolutionists have often been misled to extend the Darwinian mode to the other two levels as well. First, we have what physiologists call "adaptation": the phenotypic plasticity that permits organisms to 'mold' their form to prevailing circumstances during ontogeny. Human "adaptations" to high altitude fall into this category (while others, like resistance of sickling heterozygotes to malaria, are genetic, and Darwinian). Physiological adaptations are not heritable, though the capacity to develop them presumably is. Secondly, we have a "heritable" form of non-Darwinian adaptation in humans (and, in rudimentary ways, in a few other advanced social species): cultural adaptation (with heritability imposed by learning). Much confused thinking in human sociobiology arises from a failure to distinguish this mode from Darwinian adaptation based on genetic variation. Finally, we have adaptation arising from the conventional Darwinian mechanism of selection upon genetic variation. The mere existence of a good fit between organism and environment is insufficient for inferring the action of natural selection. (4) Adaptation and selection but no selective basis for differences among adaptations. Species of related organisms, or sub-populations within a species, often develop different adaptations as solutions to the same problem. When "multiple adaptive peaks" are occupied, we usually have no basis for asserting that one solution is better than another. The solution followed in any spot is a result of history; the first steps went in one direction, though others would have led to adequate prosperity as well. Every naturalist has his favorite illustration. In the West Indian land snail Cerion, for example, populations living on rocky and windy coasts almost always develop white, thick, and relatively squat shells for conventional adaptive reasons. We can identify at least two different developmental pathways to whiteness from the mottling of early whorls in all Cerion, two paths of thickened shells and three styles of allometry leading to squat shells. All 12 combinations can be identified in Bahamian populations, but would it be fruitful to ask why -- in the sense of optimal design rather than historical contingency -- Cerion from eastern Long Island evolved one solution, and Cerion from Acklins Island another? (5) Adaptation and selection, but the adaptation is a secondary utilization of parts present for reasons of architecture, development or history. We have already discussed this neglected subject in the first section on spandrels, spaces, and cannibalism. If blushing turns out to be an adaptation affected by sexual selection in humans, it will not help us to understand why blood is red. The immediate utility of an organic structure often says nothing at all about the reason for its being. 6. Another, and Unfairly Maligned, Approach to Evolution In continental Europe, evolutionists have never been much attracted to the Anglo-American penchant for atomizing organisms into parts and trying to explain each as a direct adaptation. Their general alternative exists in both a strong and a weak form. In the strong form, as advocated by such major theorists as Schindewolf (1950), Remane (1971), and Grassé (1977), natural selection under the adaptationist programme can explain superficial modifications of the Bauplan that fit structure to environment: why moles are blind, giraffes have long necks, and ducks webbed feet, for example. But the important steps of evolution, the construction of the Bauplan itself and the transition between Baupläne, must involve some other unknown, and perhaps "internal," mechanism. We believe that English biologists have been right in rejecting this strong form as close to an appeal to mysticism. But the argument has a weaker -- and paradoxically powerful -- form that has not been appreciated, but deserves to be. It also acknowledges conventional selection for superficial modifications of the Bauplan. It also denies that the adaptationist programme (atomization plus optimizing selection on parts) can do much to explain Baupläne and the transitions between them. But it does not therefore resort to a fundamentally unknown process. It holds instead that the basic body plans of organisms are so integrated and so replete with constraints upon adaptation (categories 2 and 5 of our typology) that conventional styles of selective arguments can explain little of interest about them. It does not deny that change, when it occurs, may be mediated by natural selection, but it holds that constraints restrict possible paths and modes of change so strongly that the constraints themselves become much the most interesting aspect of evolution. Rupert Riedl, the Austrian zoologist who has tried to develop this thesis for English audiences (1977 and 1975, translated into English by R. Jeffries in 1978) writes: The living world happens to be crowded by universal patterns of organization which, most obviously, find no direct explanation through environmental conditions or adaptive radiation, but exist primarily through universal requirements which can only be expected under the systems conditions of complex organization itself... This is not self-evident, for the whole of the huge and profound thought collected in the field of morphology, from Goethe to Remane, has virtually been cut off from modern biology. It is not taught in most American universities. Even the teachers who could teach it have disappeared. Constraints upon evolutionary change may be ordered into at least two categories. All evolutionists are familiar with phyletic constraints, as embodied in Gregory's classic distinction (1936) between habitus and heritage. We acknowledge a kind of phyletic inertia in recognizing, for example, that humans are not optimally designed for upright posture because so much of our Bauplan evolved for quadrupedal life. We also invoke phyletic constraint in explaining why no mollusks fly in air and no insects are as large as elephants. Developmental constraints, a subcategory of phyletic restrictions, may hold the most powerful rein of all over possible evolutionary pathways. In complex organisms, early stages of ontogeny are remarkably refractory to evolutionary change, presumably because the differentiation of organ systems and their integration into a functioning body is such a delicate process so easily derailed by early errors with accumulating effects. Von Baer's fundamental embryological laws (1828) represent little more than a recognition that early stages are both highly conservative and strongly restrictive of later development. Haeckel's biogenetic law, the primary subject of late nineteenth century evolutionary biology, rested upon a misreading of the same data (Gould, 1977). If development occurs in integrated packages and cannot be pulled apart piece by piece in evolution, then the adaptationist programme cannot explain the alteration of developmental programmes underlying nearly all changes of Bauplan. The German paleontologist A. Seilacher, whose work deserves far more attention than it has received, has emphasized what he calls "bautechnischer," or architectural constraints (Seilacher, 1970). These arise not from former adaptations retained in a new ecological setting (phyletic constraints as usually understood), but as architectural restrictions that never were adaptations but rather were the necessary consequences of materials and designs selected to build basic Baupläne. We devoted the first section of this paper to nonbiological examples in this category. Spandrels must exist once a blueprint specifies that a dome shall rest on rounded arches. Architectural constraints can exert a farranging influence upon organisms as well. The subject is full of potential insight because it has rarely been acknowledged at all. In a fascinating example, Seilacher (1972) has shown that the divaricate form of architecture occurs again and again in all groups of mollusks, and in brachiopods as well. This basic form expresses itself in a wide variety of structures: raised ornamental lines (not growth lines because they do not conform to the mantle margin at any time), patterns of coloration, internal structures in the mineralization of calcite and incised grooves. He does not know what generates this pattern and feels that traditional and nearly exclusive focus on the adaptive value of each manifestation has diverted attention from questions of its genesis in growth and also prevented its recognition as a general phenomenon. It must arise from some characteristic pattern of inhomogeneity in the growing mantle, probably from the generation of interference patterns around regularly spaced centers; simple computer simulations can generate the form in this manner (Waddington and Cowe, 1969). The general pattern may not be a direct adaptation at all. Seilacher then argues that most manifestations of the pattern are probably non-adaptive. His reasons vary but seem generally sound to us. Some are based on field observations: color patterns that remain invisible because clams possessing them either live buried in sediments or remain covered with a periostracum so thick that the colors cannot be seen. Others rely on more general principles: presence only in odd and pathological individuals, rarity as a developmental anomaly, excessive variability compared with much reduced variability when the same general structure assumes a form judged functional on engineering grounds. In a distinct minority of cases, the divaricate pattern becomes functional in each of the four categories. Divaricate ribs may act as scoops and anchors in burrowing (Stanley, 1970), but they are not properly arranged for such function in most clams. The color chevrons are mimetic in one species (Pteria zebra) that lives on hydrozoan branches; here the variability is strongly reduced. The mineralization chevrons are probably adaptive in only one remarkable creature, the peculiar bivalve Corculura cardissa (in other species they either appear in odd specimens or only as post-mortem products of shell erosion). This clam is uniquely flattened in an anterio-posterior direction. It lies on the substrate, posterior up. Distributed over its rear end are divaricate triangles of mineralization. They are translucent, while the rest of the shell is opaque. Under these windows dwell endosymbiotic algae! All previous literature on divaricate structure has focused on its adaptive significance (and failed to find any in most cases). But Seilacher is probably right in representing this case as the spandrels, ceiling holes, and sacrificed bodies of our first section. The divaricate pattern is a fundamental architectural constraint. Occasionally, since it is there, it is used to beneficial effect. But we cannot understand the pattern or its evolutionary meaning by viewing these infrequent and secondary adaptations as a reason for the pattern itself. Galton (1909, p. 257) contrasted the adaptationist programme with a focus on constraints and modes of development by citing a telling anecdote about Herbert Spencer's fingerprints: Much has been written, but the last word has not been said, on the rationale of these curious papillary ridges; why in one man and in one finger they form whorls and in another loops. I may mention a characteristic anecdote of Herbert Spencer in connection with this. He asked me to show him my Laboratory and to take his prints, which I did. Then I spoke of the failure to discover the origin of these patterns, and how the fingers of unborn children had been dissected to ascertain their earliest stages, and so forth. Spencer remarked that this was beginning in the wrong way; that I ought to consider the purpose the ridges had to fulfill, and to work backwards. Here, he said, it was obvious that the delicate mouths of the sudorific glands required the protection given to them by the ridges on either side of them, and therefrom he elaborated a consistent and ingenious hypothesis at great length. I replied that his arguments were beautiful and deserved to be true, but it happened that the mouths of the ducts did not run in the valleys between the crests, but along the crests of the ridges themselves. We feel that the potential rewards of abandoning exclusive focus on the adaptationist programme are very great indeed. We do not offer a council of despair, as adaptationists have charged; for nonadaptive does not mean non-intelligible. We welcome the richness that a pluralistic approach, so akin to Darwin's spirit, can provide. Under the adaptationist programme, the great historic themes of developmental morphology and Bauplan were largely abandoned: for if selection can break any correlation and optimize parts separately, then an organism's integration counts for little. Too often, the adaptationist programme gave us an evolutionary biology of parts and genes, but not of organisms. It assumed that all transitions could occur step by step and underrated the importance of integrated developmental blocks and pervasive constraints of history and architecture. A pluralistic view could put organisms, with all their recalcitrant yet intelligible complexity, back into evolutionary theory. References Baer, K. E. von, 1828, Entwicklungsgeschichte der Tiere, Konigsberg: Borntrager. Barash, D. P., 1976, Male response to apparent female adultery in the mountain bluebird: an evolutionary interpretation, Am. Nat. 110: 1097-1101. Coon, C.S., Garn, S.M., and Birdsell, J.B., 1950, Races, Springfield Oh., C. Thomas. Costa, R., and Bisol, P. M., 1978, Genetic Variability In Deep-Sea Organisms, Biol. Bull., 155: 125- 133. Darwin, C. 1872, The Origin Of Species, London, John Murray. ______, 1880, Sir Wyville Thomson and Natural Selection, Nature, London, 23: 32. Davitashvili, L.S. Teoriya Polovogo Otbora [Theory Of Sexual Selection], Moscow, Akademii Nauk. Falconer, D.S., 1973, Replicated Selection For Body Weight In Mice, Genet. Res., 22: 291-321. Galton, F, 1909, Memories of my Life, London, Methuen. Gould, S. J., 1966, Allometry and Size in Ontogeny and Phylogeny, Biol. Rev., 41: 587-640. _____, 1971, D'Arcy Thompson and The Science Of Form, New Literary Hist., 2, No. 2, 229-258. _____, 1974, Allometry in Primates, with Emphasis on Scaling and the Evolution of the Brain. In Approaches To Primate Paleobiology, Contrib. Primatol, 5: 244-292. _____, 1977, Ontogeny And Phylogeny, Cambridge, Ma., Belknap Press. _____, 1978, Sociobiology: The Art of Storytelling, New Scient., 80: 530-533. Grasse, P.P., 1977, Evolution of Living Organisms, New York, Academic Press. Gregory, W. K. 1936, Habitus Factors In The Skeleton Fossil And Recent Mammals, Proc. Am. Phil. Soc., 76: 429-444. Hamer, M., 1977, The Ecological Basis For Aztec Sacrifice. Am. Ethnologist, 4: 117-135. Jenson, H.J., 1973, Evolution Of The Brain And Intelligence, New York, Academic Press. Lande, R., 1976, Natural Selection And Random Genetic Drift In Phenotypic Evolution, Evolution, 30: 314-334. _____, 1978, Evolutionary Mechanisms Of Limb Loss In Tetrapods, Evolution, 32: 73-92. Lewontin, R.C., 1978, Adaptation, Scient. Am., 239 (3): 156-169. _______, 1979, Sociobiology As An Adaptationist Program, Behav. Sci., 24: 5-14. Morton, E.S., Geitgey, M.S., And Mcgrath, S., 1978, On Bluebird 'Responses To Apparent Female Adultery'. Am. Nat., 112: 968-971. Ortiz De Montellano, B.R., 1978, Aztec Cannibalism: An Ecological Necessity? Science, 200: 611- 617. Remane, A., 1971, Die Grundlagen Des Naturlichen Systems Der Vergleichenden Anatomie Und Der Phylogenetik. Konigstein-Taunus: Koeltz. Rensch, B., 1959, Evolution Above The Species Level. New York, Columbia University Press. Riedl, R., 1975, Die Ordnung Des Lebendigen, Hamburg, Paul Parey, Tr. R.P.S. Jefferies, Order In Living Systems: A Systems Analysis Of Evolution, New York, Wiley, 1978. _____, 1977, A Systems-Analytical Approach To Macro-Evolutionary Phenomena, Q. Rev. Biol., 52: 351-370. Romanes, G. 1., 1900, The Darwinism Of Darwin And Of The Post-Darwinian Schools. In Darwin, And After Darwin, Vol. 2, New Edn., London, Longmans, Green And Co. Rudwiek, M.J.S., 1964, The Function Of Zig-Zag Deflections In The Commissures Of Fossil Brachiopods, Palaeontology, 7: 135-171. Sahlins, M., 1978, Culture As Protein And Profit, New York Review Of Books, 23: Nov., Pp. 45-53. Schindewolf, O. H., 1950, Grundfragen der Palaontologie, Stuttgart, Schweizerbart. Seilacher, A., 1970, Arbeitskonzept zur Konstruktionsmorphologie, Lethaia, 3: 393-396. _______, 1972, Divaricate Patterns In Pelecypod Shells, Lethaig, 5: 325-343. Shea, B. T., 1977, Eskimo Craniofacial Morphology, Cold Stress And The Maxillary Sinus, Am. J. Phys. Anthrop., 47: 289-300. Stanley, S.M., 1970, Relation Of Shell Form To Life Habits In The Bivalvia (Mollusca). Mem. Geol. Soc. Am., No. 125, 296 Pp. Sweeney, B.W., and Vannote, R.L., 1978, Size Variation And The Distribution Of Hemimetabolous Aquatic Insects: Two Thermal Equilibrium Hypotheses. Science, 200:444-446. Thompson, D. W., 1942, Growth And Form, New York, Macmillan. Waddington, C.H., And Cowe, J.R., 1969, Computer Simulation Of A Molluscan Pigmentation Pattern, J. Theor. Biol., 25: 219-225. Wallace, A.R., 1899, Darwinism, London, Macmillan. Wilson, E. 0., 1978, On Human Nature, Cambridge, Ma., Harvard University Press. http://evilutionarybiologist.blogspot.be/2007/10/this-weeks-citation-classic.html Lewens, T. (2009) Seven types of adaptionism. Biol. Philos. 24:161–182. [doi: 10.1007/s10539008-9145-7] (Noot ) Als de GR nu opeens niet meer zou werken zou dat absoluut problemen opleveren in het lichaam. Het klopt dat cortisol ook kan binden aan de MR, maar die affiniteit is lager, de MR zit op andere plaatsen in het lichaam en bovendien zet de MR intracellulair een andere pathway ingang. Kort gezegd, de functie van de MR is anders dan die van de GR. Wat nou direct het grootste probleem zou opleveren bij het niet werken van de GR zou ik niet precies weten. Er zijn dus nog steeds in ons gestel receptoren aanwezig die én cortisol én aldosteron herkentnen : de in het artikel beschreven cortisol-receptor stamt van een dergelijke maar dan wel oudere versie af. De mogelijkheid om aldosteron te binden werd ook niet verloren: de oude stamreceptor-drager van beide , bracht twee nieuwe soorten receptors voort. Later zag ik(=Pierra) dat deze auteur (Joe Thornton) ook de stamboom van deze andere receptor gepubliceerd heeft, ik meen ook in Nature... Bronnen:(Van Pierra ) New Scientist Nu.nl ScienceDaily LORD HOWE STICK INSECT http://whyevolutionistrue.wordpress.com/2012/03/01/a-giant-insect-saved-from-extinction/ http://vimeo.com/14413689 http://vimeo.com/6260138 http://en.wikipedia.org/wiki/ Dryococelus_australis http://www.nhm.ac.uk/nature-online/species-of-the-day/biodiversity/endangered-species/dryococelusaustralis/index.html http://www.arkive.org/lord-howe-island-stick-insect/dryococelus-australis/ http://www.npr.org/blogs/krulwich/2012/02/24/147367644/six-legged-giant-finds-secret-hideawayhides-for-80-years Eurycantha calcarata Lucas. http://bunyipco.blogspot.com/2010/09/interesting-dilemma.html http://www.youtube.com/embed/Cs1Xs3Eheag http://www.youtube.com/embed/5apWTc23mHU Fossil GLADIATOR http://lemondedesphasmes.free.fr/spip.php?article69 Raptophasma Kernegerri Zompro 2001 Baltic amber Misverstanden rond "levende" fossielen http://sandwalk.blogspot.com/2012/04/myth-of-living-fossils.html The general public has been told time and time again that there exist among us certain species that have not evolved for millions of years. These so-called "living fossils" have somehow managed to avoid any changes in the frequencies of alleles in their evolving populations. This is, of course, impossible by any reasonable definition of evolution, a conclusion that was promoted on talk.origins two decades ago [Claim CB930:]. Yet the myth persists. It persists for three reasons: It plays into the popular misconception that natural selection is synonymous with evolution. If a species isn't adapting by obvious changes over time then it isn't evolving. Another way of saying this is that some species can be so perfectly adapted to their environment that all changes are selected against and negative selection prevents evolution. External morphological changes are the only evidence of evolution.? The so-called "living fossils" show no evidence of morphological change over millions of years when, in fact, all of the popular examples show plenty of evidence of such change. In other words, the facts are misrepresented