Community Ecology - Plattsburgh State Faculty and Research Web
advertisement

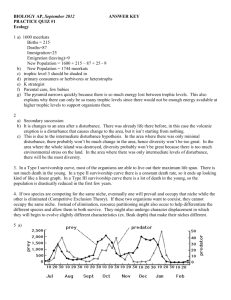
Community Ecology Community is the assemblage of populations of different organisms living in an area and potentially interacting. E.g. pond community, community of decomposers in a rotten log, forest community. Interspecific interactions Interactions between organisms of different species are referred to as interspecific interactions. These include competition, predation, parasitism, herbivory, mutualism and disease. Some interactions benefit only one participant (e.g. predation, herbivory) but some benefit both participants (mutualism). In other cases neither participant benefits (competition). Competition has been extensively studied by ecologists. Competition Interspecific competition occurs when different species compete for a resource that is in limited supply. Some resources such as air are usually not limiting so there is no competition for them. Plants compete for water, nutrients and light. Mussels and barnacles compete for space to settle on rocks in the intertidal zone. Owls, foxes and weasels compete for small mammal prey. Competitive Exclusion If competition between two species is very strong, one species may outcompete the other and competitively exclude it. Competitive exclusion first well documented by Gause in 1934. Competitive Exclusion Gause studied two species of Paramecium: P. aurelia and P. caudatum. When cultured in separate containers each species thrived and population leveled off at carrying capacity of test tube. Competitive Exclusion However, when P. aurelia and P. caudatum were grown together P. caudatum became extinct. Gause concluded P. aurelia had a competitive advantage at competing for food and outcompeted P. caudatum. Competitive Exclusion Gause developed from this and other experiments his Competitive Exclusion Principle: Two species competing for the same limiting resources cannot coexist in the same place. If the inferior competitor cannot escape the competition it will be driven to extinction. Ecological Niches The sum total uses that a species makes of the biotic and abiotic resources in its environment is referred to as its niche. (Pronounced to rhyme with “itch” in the U.S., but elsewhere to rhyme with “sheesh.”). Ecological Niches An organism’s ecological niche is analgous to its “profession,” what it does for a living. Niche includes many components: the food the organism eats, the places it occupies, the time of day it is active, the temperature range it can tolerate, etc. Ecological Niches The concept of niches can be used to restate the competitive exclusion principle: Two species cannot coexist if their niches are identical. Ecologically similar species, however, can coexist if there are significant differences in their niches. Ecological Niches As a result of competition a species fundamental niche, the niche potentially occupied by that species may be different from its realized niche the niche it actually occupies in a particular environment. Connell’s work on barnacles Joseph Connell demonstrated the effects of competition on niche occupation in barnacles. He studied Balanus balanoides and Chthamalus stellatus in the Scottish intertidal zone. These species have a stratified distribution on intertidal rocks. Connell’s work on barnacles Chthamalus occurs higher than Balanus in the intertidal zone. Connell carried out experiments in which he excluded one or other species from rocks and observed what happened. Connell’s work on barnacles When Chthamalus was excluded Balanus did not spread higher up the rocks because it apparently cannot tolerate the stress of drying out for long periods. Balanus’s realized niche is thus similar to its fundamental niche. Connell’s work on barnacles In contrast when Balanus was excluded, Chthamalus spread down the rock. The realized niche of Chthamalus in the presence of Balanus is much smaller than its fundamental niche. Connell’s work on barnacles Connell’s observations showed that although Chthamalus settled in the lower zone that Balanus smothered or crushed Chthamalus and that the most mortality occurred during the period of most rapid Balanus growth. Resource partitioning Competition between species can result in natural selection causing niches to differentiate to escape the effects of competition. Anolis lizards in the Caribbean all feed on similar prey and are similar in size, but they make use of different foraging perches. Character displacement Natural selection can also result in morphological changes in species that reduce competitive effects. Many competitors have populations that occur sympatrically (overlap their competitor) and allopatrically (geographically separate). Character displacement Often in sympatric populations the two species diverge physically. For example, among Geopsiza finches on the Galapagos Islands sympatric G. fortis and G. fuliginosa populations differ in beak depth, but measurements of allopatric populations overlap greatly. Character displacement Because beak dimensions affect the efficiency with which birds can consume different size seeds, the differentiation of bill depths in sympatry appears to reduce the intensity of competition between the two species. Other interactions Besides competition organisms engage in a wide variety of other interactions: Predation Herbivory Parasitism Disease Mutualism Predation Predation is another major interaction between organisms that shapes communities. Predators and prey both have extensive suites of adaptations designed to enable them to catch prey or avoid being caught. Adaptations Predators possess weaponry: teeth, claws poison, etc. They are also usually quick and stealthy. Prey generally flee or hide to avoid predation, but may also possess defensive structures (horns, armor, spines) or toxins (pufferfish, plants). Cryptic coloration Warning colors Many organisms have effective chemical defenses and signal them by using warning or aposematic colors. Examples include Monarch Butterfly, Coral Snake, and Poison arrow frog Coral Snake Poison arrow frog Mimicry Some organisms mimic the warning coloration of toxic organisms to gain protection. E.g. Viceroy Butterfly mimics pattern of Monarch Butterfly The mimicry of toxic organisms by non-toxic ones is called Batesian mimicry. Batesian Mimicry Monarch Butterfly Viceroy butterfly. Coral Snake and mimics. Which is the coral snake? Wasp, Hornet moth, Wasp beetle, Hoverfly Mullerian mimicry In Mullerian mimicry several toxic or dangerous species all display the same or similar warning colors. Result of convergent evolution. Mullerian mimics on left of red line Batesian on right of line Herbivory Plants are subject to grazing by many organisms and defend themselves with mechanical defenses (thorns, silica, hard shells) and by producing toxins (e.g. capascin [substance that makes chilies hot], strychnine, nicotine and tannins). Parasitism In parasitism the parasite derives its nourishment from its host. Parasites may be internal (endoparasites e.g. tapeworm, fluke) or external (ectoparasites e.g. tick, flea). In addition parasitoids lay eggs in or on prey and when the larvae hatch they consume the prey. Parasitism Many parasites have complex life cycles that include several host species (e.g. for malaria humans and mosquito are hosts). Parasites frequently modify behavior of host species so they are more vulnerable to predation by the next host species (e.g. acanthocephalan worms enter cockroach’s brain and cause it to wander about in the light where it can be caught and eaten by a rat the next host species). Disease Just like parasites pathogens which cause disease are harmful to their hosts. Pathogens include bacteria, viruses, fungi and protists (single-celled organisms). Generally microscopic. Disease Pathogens that gain access to populations that have not been previously exposed to the disease can have devastating effects. E.g. smallpox introduced to New World, Dutch Elm Disease. Mutualism Mutualistic interactions are interactions that benefit both species involved. Acacias and Pseudomyrmex ants. Tree produces hollow spines to house ants and food in form of sugar and protein –rich nodules. Ants deter herbivores, remove fungal spores and cut back competing vegetation. Acacias and Pseudomyrmex ants Mutualism Other mutualistic interactions include nitrogen fixing bacteria and legume roots, bacteria that live in the guts of grazing mammals, and mycorrhizae (association between fungi and plant roots). Trophic structure Community structure and dynamics strongly influenced by feeding relationships between organisms: the Trophic Structure. Transfer of energy up through trophic levels called the food chain. Energy travels from the primary producers (plants and other photosynthesizers) through herbivores and various carnivores. Food webs Charles Elton recognized in the 1920’s that food chains are linked together into food webs. Food webs summarize the feeding relationships in a whole community. Because many organisms feed at multiple trophic levels, food webs can be quite complex. Fig 53.13 Food webs However, webs can be simplified by grouping species with similar feeding habits into trophic groups (e.g. carnivorous plankton in the previous web). Food chain length Most food chains are relatively short, usually having no more than 5 links. There are two major hypotheses for why food chains are generally short. Food chain length Energetic hypothesis: because energy is inefficiently transferred from one trophic level to the next (only about 10% passes from one level to the next) there is too little energy left to support “superpredators.” Explains why there are no tiger-eating birds for example. Very large organisms generally feed at the bottom of the food chain (e.g. whales) Food chain length Dynamic stability hypothesis: Suggests that long food chains are less stable than short food chains and that population fluctuations at the bottom of such chains would result in extinction of top predators. Food chain length Most available data support the energetic hypothesis. In studies of tree hole communities manipulating the amount of litter deposited in the tree holes affected the length of food chains as predicted by the energetic hypothesis Food chain length Energetic hypothesis predicts longer chains when the supply of food at the base of the chain is increased and shorter chains when food supply is decreased. Australian researchers found such a pattern in a series of manipulative experiments where they adjusted leaf litter levels (the food supply) for tree hole communities. 53.15 Effects of single species on community structure Certain species can have a large impact on community structure because of their abundance or because they alter conditions for other species. Dominant species In some communities one species predominates and exerts a strong influence on the other species in the community (e.g. sugar maples shade out other plants and their shallow roots consume most available water). Dominant species Dominant species may be dominant because they are better competitors or because they have few predators or pathogens (often the case with invasive species). Removing the dominant species may or may not have a large impact on community structure. Dominant species For example, the near extinction of the American Chestnut (once the dominant tree in eastern forests before 1910 forming 40% of the large trees) had little effect on community structure. Other trees became more common, but birds and mammals were apparently almost unaffected. Keystone species Some species may not necessarily be abundant but can strongly affect community structure. These species are referred to as keystone species. Removal experiments can show the effect of keystone species. Keystone species Robert Paine removed the sea star Pisaster from a section of rocky shoreline in Washington. Pisaster preys on mussels and when Pisaster was removed the mussels increased in abundance and crowded out other invertebrates and algae. Keystone species When Pisaster was present 15 to 20 species of red algae occurred, but when it was removed the number of red algae species fell to fewer than 5. Foundation species (“ecosystem engineers” Some organisms have a profound effect on the environment and influence community structure in that way. Classic example is the beaver which through tree felling and dam building substantially modifies the environment producing ponds and wetlands used by many species Effects of disturbance on species diversity and composition Historically ecologists thought that communities were usually in equilibrium and that stability was maintained by interspecific competition. However, disturbance is now recognized as playing a major role in shaping communities. Disturbance Disturbances are events that change communities by removing species or altering resource availability. Common disturbances include fires, storms, floods, droughts and human disturbance. Disturbance Disturbance disrupts the competitive advantage of strong competitors and provides opportunities for other species by creating environmental patchiness. Disturbance Greatest diversity in communities is generally produced by intermediate levels of disturbance. Low levels allow superior competitors to predominate. High levels of disturbance exclude many species that cannot tolerate the associated stresses and also species that only colonize disturbed habitats slowly. Ecological Succession After a major disturbance a site is recolonized in a predictable sequence. Early colonizers that disperse quickly into disturbed sites are replaced by later-arriving but superior competitors. Ecological Succession McBride Glacier in Glacier Bay, Alaska has retreated steadily since 1760. Pattern of vegetation development in wake of retreating glacier follows clear pattern. First, in pioneer stage exposed ground colonized by mosses, liverworts, fireweed and Dryas (a mat forming shrub), willows and cottonwoods. After about 30 years Dryas predominates Next alder trees invade and form dense thickets. The alder stands are next invaded by Sitka Spruce which overgrow them forming a thick forest after about 100 years. Finally hemlocks invade the spruce forests and after another 100 years spruce-hemlock forest forms. Biogeographic effects on community diversity Communities differ greatly in the number of species they contain. A community’s species diversity is strongly influenced by its geographic location and its size. Biogeographic effects on community diversity Plant and animal life is more abundant near the topics and small or remote islands have fewer species than large island or those near continents. These observations suggest that community diversity may be strongly affected by biogeographic processes. Equatorial-polar gradient Tropical habitats support many more species than temperate ones. 6.6 ha plot in Malaysia contained 711 species of tree. A similar plot in Michigan contains only 10-15 species. Equatorial-polar gradient Two key factors in equatorial-polar gradient are probably evolutionary history and climate. Over evolutionary time species diversity may increase as speciation occurs. Tropical habitats are older (having been less disturbed by e.g. glaciations) and so there has been more time for speciation to occur. Equatorial-polar gradient Main reason for higher diversity at tropics, however, is probably climate. There is a very strong correlation between solar energy input and water availability (combined into one measure as evapotranspiration) and species richness. Evapotranspiration highest in areas of high temperatures and rainfall. Area effects Species-area curve. Everything else being equal, the larger the area a community occupies the greater the number of species. Most likely this is because larger areas include a greater diversity of habitats and microhabitats than smaller areas. Area effects Species–area curves differ among taxa, but basic pattern applies across all groups. In conservation biology knowledge of species area curves for key taxa in communities allows ecologists to predict how loss of habitat may alter a community’s biodiversity. Island Equilibrium Model Islands have been popular model systems for studying the biogeographic factors that affect community diversity. “Island” refers not only to true islands but to any patch surrounded by unsuitable habitat (e.g. ponds, forest fragments, mountain peaks). Island Equilibrium Model MacArthur and Wilson in the 1960’s developed a general model to explain diversity on islands. Simple model used extinction and immigration rates to predict the number of species an island could support. Island Equilibrium Model MacArthur and Wilson assumed that small islands would have higher extinction rates than large islands because small islands have smaller populations. Also assumed that large islands and islands close to landmasses would have higher rates of immigration than smaller or more distant islands. Island Equilibrium Model Because immigration and extinction are opposing forces, by graphing the two curves (immigration and extinction) MacArthur and Wilson were able to predict an equilibrium number of species that an island should contain. Island Equilibrium Model The MacArthur and Wilson model is simple and applies only when colonization is major factor affecting species composition, but had a major effect on stimulating research into the effects of habitat size on species diversity (a topic of vital importance to conservation biology).