Cell Cycle Control Abnormalities result in
cancer or developmental disorders
Cell Growth and Oncogenesis
Cancer cells
Proliferation
c40 divisions
Normal cells
- PCD
Senescent
DNA errors
1.
Reduction in differentiation and/or
PCD – no change in rate of
proliferation
2.
Increased rate of proliferation
without inhibition of
differentiation/PCD
Time
100
Cancer diameter (mm)
Cell number
Mechanisms
1012 cells
death
10
109 cells
lump
1
0.1
108 cells
X-ray visible
10
20
Divisions
30
40
Early detection saves lives
Cancer and the Cell Cycle
• Basic function to
replicate exactly
chromosomal DNA
• DNA duplication
occurs in S
(synthesis) phase
• Cell division in M
phase
• Gap phases G1 and
G2
Phases of the Cell Cycle
• G1 phase between M
phase and S-phase
• G2 phase S-phase
and mitosis
• G1 G2 S called the
interphase
• Monitoring and
deletion function
Flow Cytometry of DNA content
• DNA content in a cell
population identifies
cells with 2x content
as dividing cells
• Tumours are
frequently aneuploid
ie irregular cellular
DNA content
S-phase identification
• Thymidine
incorporation
• BrdU incorporation
• Can be measured
easily in whole
animals and in tissue
sections
The concept of the Check point
• Multiple errors can
occur in this process
• Errors must be
controlled
• Elaborate machinery
to control for errors
• Genetic errors in error
controlling machinery
may be crucial
Cyclins
• Originally identified in yeast
• Tend to be evolutionarily conserved
• Regulate progression through phases of
the cell cycle
• Act as regulatory units for cyclin
dependent kinases CDKs
• Cyclin –CDK complex regulated by
phosphorylation
Cyclins and Cyclin-dependent
Kinases
Control of the Cell Cycle
B
Cyclin Levels
A
Interactions between
cyclins and cyclindependent kinases
regulate cycle
D
E
G0
G1
S
cyc
cycA
G2
M
cycB
cycB
cdc25
CDK
cycD
cycE
CDK4
CDK2
p27
p27 may regulate exit
from G0
CDK2
pY
P
Rb
P
P
Rb
Unphosphorylated
Rb forms complex
with E2F
cdc2
cdc2
Y
Phosphatase cdc25
dephosphorylates cdc2
allowing entry into M
phase
E2F1
No transcription
E2F1
Transcription
Phosphorylation of Rb
(cycD-CDK4) promotes
dissociation of E2F1
and Rb
Viral products subvert RB function
P53 discovery
SV40 T is a viral protein that transforms human cells
P53- “guardian of the genome”
• P53 activated
following genotoxic
insult
• Induces transcription
of p21
• P21 locks CDK in the
off position
P53 and cancer
•
•
•
•
•
Mutations cause Li Frameini syndrome
Multiple cancers in different organs
Somatic mutations often occur in cancer
Somatic mutations affect cell growth
Somatic mutations often signal
progression to malignant phase
The p53 Signaling Pathway
SIGMA-ALDRICH
Check-point Control – an integrated
picture
Onyx virus targets cells with nonfunctioning p53
ONYX 015
P53 gene therapy – prostate and
lung
Drugs may have phase-specific
effects
• Cytotoxic effects are
frequently phase
specific
• Multiple phases may
be targeted
• Single stage may be
targetted by multiple
drugs
End of Talk
Supplemental Information
G and S Phases of the Cell Cycle
1
SIGMA-ALDRICH
CDK4
Why is this important?
Colon Cancer CDK4
CDK4 modelling
and drug design
G2 and M Phases of the Cell Cycle
SIGMA-ALDRICH
Regulatory Cascade of Cyclin Gene Expression
SIGMA-ALDRICH
P27 and breast cancer
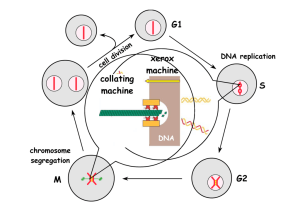
Structure of the cell cycle
The cell cycle consists of four phases.
Gap 1 (G1) is the interval between mitosis and DNA
replication that is characterized by cell growth.
If the conditions that signal transition to replicative phase
are not present, the cell exits the cell cycle and enters G0, a
nonproliferative phase during which growth, differentiation
and apoptosis occur.
DNA replication occurs during the synthesis (S) phase,
which is followed by a second gap phase (G2) during which
growth and preparation for cell division occurs.
Mitosis occur in M phase.
G1 and S Phases of the Cell Cycle
In proliferating cells, the cell cycle consists of four phases. Gap 1 (G1) is the interval between mitosis
and DNA replication that is characterized by cell growth. The transition that occurs at the restriction
point (R) in G1 commits the cell to the proliferative cycle. If the conditions that signal this transition
are not present, the cell exits the cell cycle and enters G0, a nonproliferative phase during which
growth, differentiation and apoptosis occur. Replication of DNA occurs during the synthesis (S)
phase, which is followed by a second gap phase (G2) during which growth and preparation for cell
division occurs. Mitosis and the production of two daughter cells occur in M phase.
Passage through the four phases of the cell cycle is regulated by a family of cyclins that act as
regulatory subunits for cyclin-dependent kinases (cdks). The activity of the various cyclin/cdk
complexes that regulate the progression through G1 -S-G2 phases of the cell cycle is controlled by
the synthesis of the appropriate cyclins during a specific phase of the cell cycle. The cyclin/cdk
complex is then activated by the sequential phosphorylation and dephosphorylation of the key
residues of the complex, located principally on the cdk subunits.
The cyclin cdk complex of early G1 is either cdk2, cdk4, or cdk6 bound to a cyclin D isoform. There
are several proteins that can inhibit the cell cycle in G1. If DNA damage has occurred, p53
accumulates in the cell and induces the p21-mediated inhibition of cyclin D/cdk. Mdm2, by facilitating
the nuclear export/inactivation of p53, becomes part of an inhibitory feedback loop that inactivates
p21-mediated G1 arrest. Similarly, activation of TGF- receptors induces the inhibition of cyclin D/cdk
by p15, while cyclic-AMP inhibits the cyclin D/cdk complex via p27. If the cyclin D/cdk complex is
inhibited, retinoblastoma protein (Rb) is in a state of low phosphorylation and is tightly bound to the
transcription factor E2F, inhibiting its activity.
Passage through the restriction point and transition to S phase is triggered by the activation of the
cyclin D/cdk complex, which phosphorylates Rb. Phoshporylated Rb dissociates from E2F, which is
then free to initiate DNA replication. Cyclin E/cdk2 accumulates during late G phase and triggers the
passage into S phase. The entire genome is replicated during S phase. The synthesis and
accumulation of cyclin B/cdc2 also begins during S phase, but the complex is phosphorylated at
Thr14 -Tyr15 and is inactive. Cyclin A/cdk2 accumulates during S phase and its activation triggers the
transition to G2, a phase characterized by the accumulation of cyclin B/cdc2, the inhibition of DNA
replication, cell growth and new protein synthesis.
G2 and M Phases of the Cell Cycle
The transition from G2 phase to mitosis is triggered by the Cdc25-mediated activation
(dephosphorylation) of the cyclin B/cdc2 complex (MPF). The activation of cyclin B/cdc2 that
is necessary for G/M progression is currently the most well-characterized step in the cell
cycle. CyclinB/cdc2 is activated by phosphorylation of Thr160 and the dephosphorylation of
Thr14 -Tyr15. Thr160 is phosphorylated by cyclin activating kinase (CAK), following the
activation of CAK by a cyclin activating kinase activating kinase (CAKAK). However, the
complex is kept in an inactive state due to the phosphorylation of Thr15, which is catalyzed
by the Wee1 kinase. Cyclin B/cdc2 activation is triggered when Cdc25, a phospatase,
dephosphorylates Thr15. In turn, the activity of Cdc25 is regulated by both activating and
inhibitory phosphorylations. Phosphorylation of Ser by Chk1 (a check point activated kinase
that participates in the G2-arrest of cells with damaged DNA) leads to the inactivation of
Cdc25, while phosphorylation by an M-phase activated kinase creates a positive feedback
loop leading to the rapid activation of the cyclin B/cdc2 complex.
MPF catalyzes the phosphorylation of lamins and histone 1, and is involved in the regulation
of events preceding cell division, such as spindle formation, chromatin condensation, and
fragmentation of the nuclear envelope and of organelles such as the Golgi and endoplasmic
reticulum. The metaphase to anaphase transition is triggered by inactivation of MPF and the
degradation of cyclin B. This induces the separation of chromatids and their movement to
the poles of the mitotic spindle, after which the mitotic apparatus disappears, the nuclear
membranes reform and the nucleoli reappear. During cytokinesis, the cytoplasm divides and
the resulting daughter cells enter G1.
References:
Smits, V.A., and Medema, R.H., Checking out the G2/M transition. Biochim.Biophys.Acta., 1519, 112 (2001).
Taylor, W.R., and Stark, G.R., Regulation of the G2/M transition by p53. Oncogene, 20, 1803-1815
(2001).
Bulavin, D.V., et al., p38 and Chk1 kinases: different conductors for the G2/M checkpoint symphony.
Curr. Opin. Genet. Dev., 12, 92-97 (2002).
The p53 Signaling Pathway
The tumor-suppressor protein p53 exhibits sequence-specific DNA-binding,
directly interacts with various cellular and viral proteins, and induces cell cycle
arrest in response to DNA damage. In response to signals generated by a
variety of genotoxic stresses, e.g, UV irradiation or DNA damage, p53 is
expressed and undergoes post-translational modification that results in its
accumulation in the nucleus. The p53-dependent pathways help to maintain
genomic stability by eliminating damaged cells, either by arresting them
permanently or through apoptosis. For example, -irradiation activates p53 to
turn on the transcription of p21CIP1, that, in turn, binds to and inhibits cyclindependent kinases, causing hypophosphorylation of retinoblastoma (Rb), thus
preventing the release of E2F and blocking the G1-S transition. Some of the
cellular effects of p53 can be blocked by the deregulated expression of c-Myc,
Bcl-2, or E2F. p53 activity is controlled through an autoregulatory loop involving
Mdm2. The binding of Mdm2 to p53 targets p53 for degradation and inhibits
p53-induced cell-cycle arrest and apoptosis.
References
Gu, J., et al., Identification of a sequence element from p53 that signals for Mdm2-targeted
degradation. Mol. Cell Biol., 20, 1243-1253 (2000).
Jimenez, G.S., et al., p53 regulation by post-translational modification and nuclear retention
in response to diverse stresses. Oncogene, 18, 7656-7665 (1999).
King, K.L., and Cidlowski, J.A., Cell cycle regulation and apoptosis. Annu. Rev. Physiol., 60,
601-617 (1998).
Regulatory Cascade of Cyclin Gene Expression
When cells traverse the G0 to G1 phase to the S-phase transition, a series of cyclindependent kinases is activated. The addition of serum growth factors to quiescent cells
promotes transcription of the cyclin D1 gene. Cyclin D1 then associates with pre-existing
cdk4 to form an active complex. The kinase activity associated with this complex can
phosphorylate specific sites on the retinoblastoma protein (pRb), leading to inactivation of
pRb and the activation cyclin E transcription by E2F. Activation of the cyclin E gene can be
blocked by the cdk inhibitor p16. Cyclin E associates with existing cdk2 and this active
complex regulates the function of several sets of target proteins. First, cyclin E/cdk2
complexes associate with E2F/p107 complexes to activate expression of the cyclin A gene.
Also, cyclin E/cdk2 complexes cooperate with cyclin D1 to amplify the phosphorylation of
pRb. Cyclin A associates with cdk2 to form a kinase complex that phosphorylates
downstream targets involved in the initiation of DNA replication.
References
Ohtani, K., Implication of transcription factor E2F in regulation of DNA replication. Front.
Biosci., 4, D793-D804 (1999).
Lania, L., et al., Transcriptional control by cell-cycle regulators: a review. J. Cell. Physiol.,
179, 134-141 (1999).
Hatakeyama, M., et al., The role of RB in cell cycle control. Prog. Cell Cycle Res., 1, 9-19
(1995).
 0
0