W09micr430Lec16 - Cal State LA

Bacterial Physiology (Micr430)
Lecture 16
Bacterial Development
(Text Chapter: 18.15; 18.18)
Bacterial development
Cellular differentiation in which a cell acquires phenotypic properties that clearly differentiate it from a precursor cell;
Cellular differentiation in which a cell divides to produce 2 daughter cells that can be distinguished morphologically and/or physiologically; or
Multicellular development to form specialized structures – fruiting bodies and biofilms
Bacterial development
During development (or differentiation), differential gene expression requires cellto-cell signaling (quorum sensing) as well as signaling within cells (two-component systems)
Differentiation Advantages
Generating “resting” cell forms that are more resistant to environmental stresses such as endospores and myxospores
Generating a number of cell forms specifically
“designed” for dispersal of bacteria (such as swarmer cells of Caulobacter )
Producing cell forms performing specific functions (heterocysts of Anabaena)
Producing cell forms designed to establish a symbiotic relationship with another organism for their mutual benefit such as the nitrogenfixing nodules.
Endospore Forming Genera
Six separate genera of bacteria produce a developmental forms called endospores
Bacillus
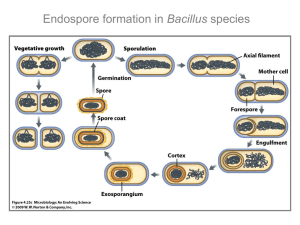
Endospore Formation
Life Cycle of
Bacillus
All endospore-forming bacteria undergo a life cycle that includes vegetative growth in the presence of adequate nutrition and favorable environmental conditions.
At the beginning of the stationary phase, when nutrients become limiting, the cells have a variety of overlapping genetic networks available with which they can respond to this changing environment.
Among the processes that can be activated are motility and transformation competence.
The decision to undergo sporulation is a response of last resort to survive.
Life Cycle of
Bacillus
As cells enter stationary phase, nutritional deprivation can trigger entry into sporulation.
This process can be visualized by light and electron microscopy as a series of complex morphological changes that result in the formation of a highly resistant dormant form called endospore.
When conditions become favorable again, the spore can undergo activation, germination and outgrowth into a metabolically active cell capable of entering vegetative growth cycle.
Sporulation in
Bacillus
The decision to sporulate is regulated by a phosphorelay signal transduction system.
Sporulation is only one of several physiological changes during adaptation to nutrient deprivation.
Motility increases chances of finding nutrients
Motile cells secrete degradative enzymes to generate food sources.
Stages of Sporulation
Sporulation can be divided into seven stages.
Stage 0 (vegetative cells)
Stage I (axial filament formation)
Stage II (septum and prespore formation)
Stage III (forespore development)
Stage IV (cortex formation)
Stage V (coat formation)
Stage VI (maturation)
Stage VII (release of the mature spore)
Stages of Sporulation in Bacillus
Sporulation in
Bacillus
Sporulation is an example of cell division ending in 2 different developmental fates for the daughter cells.
B. subtilis
differentially expresses genes in the mother cell and forespore, resulting in two different types of cells, by compartmentalization of sigma factors, which determine which genes are expressed.
Sporulation genes
B. subtilis
sporulates when it expresses
spo
genes, which were discovered by examining mutants that failed to complete sporulation.
The genes are named after the stage of blockage and are distinguished from one another by a letter.
E.g., mutants in
spo0A
fail to initiate sporulation and do not proceed to stage I
Sigma Factors in Bacillus subtilis
Sigma subunit of RNAP determines the specificity of promoter utilization.
There are at least 10 different sigma factors in
B. subtilis
, each of which directs RNAP to a different set of promoters.
Most sigma factors make sequencespecific contacts at -10 and -35 regions upstream of regulated genes or operons.
Spo0A Protein
The amino-terminal portion of the Spo0A protein is homologous to response regulators of procaryotic two-component systems. It is this portion of the molecule that is phosphorylated
(via phosphorelay) to activate its transcription functions.
The carboxy half of the protein contains the
DNA-binding specificity of the protein and interacts with the transcriptional machinery.
Higher levels of Spo0A-P stimulate axial filament formation, polar septation and transcription of genes required for cell typespecific gene expression.
Phosphorelay
There are five histidine sensor kinases involved in sporulation but two are most important (Kinases A and
B) for sporulation in lab media by phosphorylating
Spo0F.
Spo0F has a strong similarity to the response regulators in two-component systems except it lacks an additional carboxy-terminal domain. Its function is to accept phosphate from the activating kinases for the phosphorelay and serve as a substrate for the Spo0B protein.
Spo0B protein is the phosphoprotein phosphotransferase. The enzymatic function of the Spo0B is to transfer phosphate from Spo0F-P to Spo0A, producing
Spo0A-P.
Phosphorelay system
Role of phosphatases
In phosphorelay that controls sporulation, the levels of the phosphorylated response regulator proteins are determined by multiple specialized phosphatases
The existence of multiple phosphatases alllows multiple inputs into the regulation of the phosphorelay pathway
One of the phosphatases, Spo0E, dephosphorylates the master transcriptional factor Spo0A-P
Role of phosphatases
Two of the phosphatases, RapA and
RapB, dephosphorylate the phosphorelay protein, Spo0F-P
A two-component signal system consisting of ComP (HK) and ComA (RR) regulates gene expression of an operon containing
rapA
and
phrA
genes
Since ComP/ComA system produces an inhibitor of the RapA phosphatase, it stimulates sporulation
Regulation of RapA phosphatase