Gastrulation & Organogenesis
advertisement

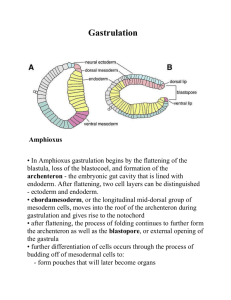
Overview: A Body-Building Plan for Animals It takes only about 9 months for a single-celled zygote to be transfigured into a newborn human, built of trillions of differentiated cells organized into specialized tissues and organs. Tail Brain Pharyngeal Pouches Heart 6 to 8 week human embryo Limb bud 1 mm Vertebrae Preformation 2000 yrs. earlier, Aristotle proposed “homunculus” epigenesis – form emerges gradually 1694 Development is determined by the genome of the zygote & differences that arise between early embryonic cells that set the stage for the expression of different genes. In some species, this is due to the uneven distribution of cytoplasmic determinants in the unfertilized egg. These affect development of the cells that inherit them during mitosis. In other species, this is due to their location in different embryonic regions. In most, it’s a combination. Selectively controlled gene expression leads to cell differentiation (specialization). The timely communication of instructions during morphogenesis occurs by cell signaling among embryonic cells, so differentiated cells end up in the appropriate locations. 47.1: After fertilization, embryonic development proceeds through cleavage, gatrulation, & organogenesis 1 Contact – The sperm contacts the egg’s jelly coat, triggering exocytosis from the sperm’s acrosome. 2 Acrosomal reaction – Hydrolytic enzymes make a hole in the jelly coat, while actin filaments form the acrosomal process. This protrudes from the sperm head and penetrates the jelly coat, binding receptors in the egg membrane that extend through the vitelline layer. 3 Fusion – A hole is made in the vitelline layer, allowing contact and fusion of the gamete membranes. The membrane becomes depolarized, resulting in the fast block to polyspermy. 5 Cortical reaction – Fusion of the membranes triggers an increase of Ca2+ in the egg’s cytosol, causing cortical granules to fuse with the plasma membrane. This leads to swelling of the perivitelline space, hardening of the vitelline layer, and clipping of sperm binding receptors. The fertilization envelope is the slow block to polyspermy. One function of fertilization is egg activation. Acrosomal reaction Sperm plasma membrane Contact and fusion of sperm and egg Entry of sperm membranes 4 nucleus Cortical reaction Contact Basal body (centriole) Sperm head Sperm nucleus Actin Acrosome Jelly coat Sperm-binding receptors Acrosomal process Na+ Na+ Na+ + Fused plasma Na Na+ Cortical membranes 2+ + Ca Ca2+Na Na+ granule Ca2+ Ca2+Na+ Hydrolytic enzymes Perivitelline 2+ Ca space + 2+ Cortical granule Na Vitelline layer Ca 2+ Ca membrane Ca2+Na+ Egg plasma EGG CYTOPLASM Ca2+ 2+ 2+ membrane Ca2+ Ca Ca Fertilization envelope Na+ 500 µm More About The Cortical Reaction 1 sec before fertilization 10 sec after fertilization Point of sperm entry 20 sec 30 sec Spreading wave of calcium ions Sperm binding appears to activate an IP3-DAG signal transduction pathway that causes Ca2+ to be released from ER beginning at the site of sperm entry and propagating in a wave across the egg. Within seconds, the high [Ca2+] induces fusion of cortical granules that release mucopolysacharides, producing an osmotic gradient, drawing water into the perivitelline space, separating the vitelline layer. Cortical granule enzymes: 1) degrade proteins that hold the vitelline layer to the plasma membrane, 2) clip off sperm receptor proteins and 3) harden the vitelline layer into the fertilization envelope. Although studied in detail in sea urchins, this is known to occur in vertebrates, including mammals. Activation of the Egg Ca2+, Seconds Increased [Ca2+] also induces increased pH, cellular respiration & protein synthesis (activated). Sperm cells do not contribute any materials. Eggs can be first cell division occurs. Minutes activated by injection of & temperature shock, (parthenogenesis), even with the egg nucleus removed! Proteins & mRNAs in the egg cytoplasm are sufficient for activation. The sperm nucleus begins to swell and merges with the egg nucleus forming the diploid zygote. DNA synthesis begins and the In other species the timing differs partly because the eggs are arrested at a specific stage of meiosis (humans = metaphase II). Upon fertilization, meiosis is quickly completed. 1 Binding of sperm to egg 2 3 4 Acrosomal reaction & fusion: plasma membrane depolarization (fast block to polyspermy) 6 8 10 20 30 40 50 1 Increased intracellular calcium level Cortical reaction begins (slow block to polyspermy) Formation of fertilization envelope complete 2 Increased intracellular pH (MINE = not in mouse!) 3 4 5 Increased cellular respiration & protein synthesis 10 20 30 40 60 90 Fusion of egg and sperm nuclei complete Onset of DNA synthesis First cell division Fertilization in Mammals In terrestrial animals, fertilization is internal. Female reproductive tract secretions “activate” sperm by altering molecules on the surface and increasing motility. Human sperm requires ~6 hrs exposure. Nuclei do not fuse immediately. Their envelopes disperse and the chroms. come together after mitosis. (12-36 hrs.) 1 Sperm migrates through follicle cells and binds to receptors (not shown) in the zona pellucida. 2 Acrosomal reaction – Hydrolytic enzymes make a hole in the zona pellucida. 3 Breakdown of the zona pellucida allows the sperm to reach the egg plasma membrane. Sperm membrane proteins bind receptors on the egg membrane and they fuse. 4 The sperm nucleus and other components enter the egg. (basal body becomes centrosome - duplicates) 5 Cortical reaction – Cortical granule enzymes harden the zona pellucida, which blocks polyspermy. (no fast block) Follicle cell 4 3 2 1 Zona pellucida Egg plasma membrane Acrosomal vesicle 5 Sperm Cortical basal granules body Sperm nucleus EGG CYTOPLASM Establishing The Axes Polarity of the egg determines the anterior-posterior axis before fertilization. The animal hemisphere is deep gray due to melanin granules in the cortex. The absence of melanin in the vegetal hemisphere allows the yellow yolk to be visible. The animal membrane and associated cortex rotate toward the point Animal hemisphere Animal pole Point of sperm entry Vegetal hemisphere of sperm entry. The vegetal cortex rotates toward the animal hemisphere. This allows molecules in the vegetal cortex to interact with cytoplasmic Vegetal pole molecules in the animal hemisphere, forming cytoplasmic determinants that initiate development of dorsal structures. Cortical rotation establishes the dorsal-ventral axis and also exposes the gray crescent, a marker for the dorsal side that is covered by animal cortex prior to rotation. The first cleavage bisects the gray crescent. Point of sperm entry Gray crescent Anterior Once anterior-posterior and dorsal-ventral are defined, Right so is the left-right axis. Future dorsal side Dorsal Ventral Left First cleavage Posterior Cleavage Blastomeres Blastocoel fertilization envelope Zygote Four-cell stage Morula Blastula During cleavage, cells undergo S and M cell cycle stages, but virtually skip the G1 and G2 stages. This results in no protein synthesis and the embryo does not enlarge. The cytoplasm is partitioned into blastomeres. The first 5-7 divisions form a cluster, known as the morula within which the fluidfilled blastocoel begins to form and is completed in the blastula, a hollow ball of cells. Different regions of cytoplasm with different cytoplasmic determinants end up in separate blastomeres. Uneven distribution of mRNAs, proteins and yolk determine the egg and zygote’s polarity. (NOT IN MAMMALS) The planes of division follow a specific pattern relative to the poles. Yolk is most concentrated toward the vegetal pole and decreases toward the animal pole (also where polar bodies are budded from during oogenesis). Cleavage In the frog, the first two divisions are meridional (vertical), resulting in four blastomeres of equal size extending from the animal pole to the vegetal pole. The third, is equatorial (horizontal). However, the uneven distribution of yolk in the zygote, displaces the mitotic apparatus so that the four blastomeres in the animal hemisphere are smaller. The effect of the yolk persists, so that the blastocoel is located in the animal hemisphere. Zygote 0.25 mm 2-cell stage forming 4-cell stage forming Other animals have less yolk, but still have an animal-vegetal axis due to uneven distribution of other substances. The blastomeres are more likely to be of similar size and the blastocoel centrally 8-cell located, but the general cleavage pattern seen in stage frogs is seen in all deuterostomes. Animal pole Blastula (cross section) Eight-cell stage (viewed from the animal pole) 0.25 mm Blastocoel Vegetal pole Blastula (at least 128 cells) Cleavage Disk of cytoplasm Yolk Fertilized egg In birds, what we call the yolk is actually the egg cell with a very small disk of cytoplasm at the animal pole. This enormous cell is surrounded by a protein-rich solution (white) that also provides nutrients. Cleavage is restricted to the yolk-free cytoplasm (meroblastic cleavage vs. holoblastic cleavage). This produces Zygote Four-cell stage (just the disk) cap of cells called the blastoderm which sort into upper (epiblast) and lower (hypoblast) layers. The cavity between is the blastocoel and this embryonic stage is the avian equivalent of the blastula. In insects, the zygote’s nucleus is within a mass of yolk. Cleavage begins with the nucleus undergoing mitotic divisions without cytokinesis. Several hundred nuclei spread throughout the yolk, then migrate to the outer edge of the embryo. Membrane forms around each, forming the blastula, which consists of a single layer of about 6,000 cells surrounding a mass of yolk. Blastoderm Cutaway view of the blastoderm Blastocoel BLASTULA YOLK MASS Epiblast Hypoblast Gastrulation (Sea Urchin) VIDEO CLIP The morphogenetic process called gastrulation is a dramatic Future ectoderm rearrangement of the blastula to form a 3-layered embryo Future mesoderm Germ Layers Future endoderm (gastrula) with a primitive gut. Gastrulation is driven by Animal pole the same general mechanisms in all species: changes in Blastocoel cell motility, shape and adhesion. The three layers Mesenchyme produced, are embryonic germ layers that will eventually cells develop into all the tissues and organs of the adult animal. Vegetal Vegetal Key plate Gastrulation begins at the vegetal pole where cells detach and enter the blastocoel as migratory mesenchyme cells. Cells near the vegetal pole flatten into a vegetal plate that buckles inward through invagination forming the endodermlined archenteron. Mesenchyme cells send out filopodia from the archenteron tip that contract and drag the archenteron across the blastocoel. The open end of the archenteron, which will become the anus, is called the blastopore. When the other end touches the ectoderm, the two layers fuse, forming the mouth, completing the primitive digestive tube (gut). pole Blastocoel Filopodia pulling archenteron tip Archenteron Mesenchyme cells Blastopore 50 µm Ectoderm Mouth Some of the mesenchyme cells (mesoderm) will eventually secrete calcium carbonate to form a simple internal skeleton. Mesenchume The gastrula develops into a ciliated larva that drifts as plankton until it metemorphoses into the adult benthic animal. Blastocoel (mesoderm forms future skeleton) Archenteron Blastopore Digestive tube (endoderm) Anus (from blastopore) GASTRULA Gastrulation (Frog) Gastrulation is more complicated in the frog: 1) yolk-laden cells of the vegetal hemisphere 2) blastula wall more than one cell thick. Gastrulation begins on the dorsal side along the gray crescent and produces the dorsal lip of the blastopore. Invagination continues to be initiated until the two ends of the blastopore meet on the ventral side forming a complete circle. All along the blastopore, future mesoderm and endoderm cells roll over the edge of the lip into the interior (involution). CROSS SECTION SURFACE VIEW Animal pole Blastocoel Dorsal lip of blastopore Dorsal lip of blastopore Vegetal pole BLASTULA Blastocoel shrinking Archenteron Involution The blastocoel collapses during this process, displaced by the archenteron cavity and the blastopore encercles a yolk plug. As is the sea urchin, the frog is a deuterostome. Blastocoel remnant VIDEO CLIP Ectoderm Mesoderm Endoderm Key Future ectoderm Future mesoderm Future endoderm Yolk plug Yolk plug GASTRULA Gastrulation (Chick) BLASTODERM Epiblast Primitive streak Future ectoderm Endoderm Migrating cells (mesoderm) Hypoblast YOLK All the cells that will form the embryo come from the epiblast. Cells move toward the midline of the blastoderm, then detach and move inward through the primitive streak. Some move down to become endoderm, some laterally into the blastocoel to form mesoderm and remaining epiblast cells become ectoderm. Hypoblast cells later segregate from the endoderm and form portions of the yolk sac and a stalk connecting the yolk mass to the embryo. Organogenesis (Frog) Organogenesis involves more localized morphogenic changes in tissue. Folds, splits and dense clustering (condensation) begin organ formation. The notochord Neural folds Neural fold is formed from dorsal mesoderm that condenses just above the archenteron. Neural plate Outer layer of ectoderm Signals from the notochord cause the LM 1 mm Neural Neural fold plate Notochord Ectoderm Mesoderm Endoderm Archenteron Neural plate formation ectoderm above it to form the neural Neural tube plate, which curves inward, rolling itself Neural crest into the neural tube. The neural crest develops where the tube pinches off from the ectoderm. These cells migrate to various parts of the embryo, forming peripheral nerves, teeth, skull Neural crest bones, and many other types of cells. Fourth germ layer?? Formation of the neural tube Organogenesis (Frog) Somites Eye SEM Neural tube Tail bud 1 mm Notochord Neural crest Coelom Somite Somites are condensations that occur in strips of mesoderm lateral to the notochord. Some dissociate into mesenchymal cells. The notochord functions as a core around which somites gather and form vertebrae. Parts of the notochord persist as the inner portion of vertebral disks. Somite cells also form the muscles of the axial skeleton. Lateral to the somites, the mesoderm splits into two layers that form the lining of the coelom. In the chick, the borders of the blastoderm fold downward and come together, pinching the embryo into a three-layered tube joined to the yolk. Other events occur much as they do in the frog. Rudiments of most major organs have formed by 56 yours. Organogenesis (Chick) Eye Neural tube Notochord Archenteron (digestive cavity) Somites Forebrain Somite Coelom Endoderm Mesoderm Ectoderm Archenteron Lateral fold Heart Blood vessels Somites Yolk stalk Form extraembryonic Yolk sac membranes Neural tube YOLK Early organogenesis Late organogenesis Adult derivatives of the three embryonic germ layers in vertebrates. Developmental Adaptations of Amniotes (dehydration & shock) Amnion Amniotic cavity with amniotic fluid (metabolic waste disposal & gas exchange with chorion) Allantois Albumen (nutrients, dehydration & shock) Embryo Yolk Shell (semipermeable) (nutrients) Chorion (gas exchange) Yolk sac (blood vessels transport yolk nutrients) Vertebrate embryos require an aqueous envirnonment. Two effective structures have evolved to enable this on land: 1) The shelled egg, and 2) the uterus. In both, embryos are surrounded by fluid within a sac formed by the amniotic membrane, or amnion. Reptiles, birds & mammals are amniotes. Germ layers outside the embryo develop into four extraembryonic membranes that provide life support for the embryo. NOTE: Each membrane forms from two germ layers. Mammalian Development Uterus -Fertilization and the earliest stages of development are in the oviduct. -Eggs are small, store little nutrients, and have not been shown to be polar. -Cleavage is holoblastic, however, gastrulation and early organogenesis Blastocyst is similar to birds and reptiles. arrives -Early cleavage is slow (human: 1st @ 36 hrs., 2nd @ 60 hrs., 3rd @ 72 hrs.) -Blastomeres are equal size and tightly adhere, so the embryo appears smooth. Maternal blood vessel 1 At the completion of cleavage, the embryo has over 100 cells (blastocyst) around a central cavity (blastocoel). At one end, is the inner cell mass, which develops Blastocyst into the embryo and extraembryonic implants. membranes. Endometrium (uterine lining) Inner cell mass Trophoblast Blastocoel Expanding region of trophoblast Epiblast Hypoblast Trophoblast 2 The trophoblast (epithelium) secretes enzymes that break down the endometrium (implantation). The trophoblast thickens and extends projections into the maternal tissue, eroding capillaries, so that blood bathes trophoblastic tissues. The inner cell mass forms a flat disk with an epiblast and a hypoblast. As in birds, the human embryo develops almost entirely from epiblast cells. Mammalian Development 3 Epiblast cells move inward through a primitive streak to form mesoderm and endoderm. Amnion At the same time, extraembryonic membranes begin to form. The invading trophoblast, mesodermal cells derived from the epiblast, and adjacent endometrial tissue all contribute to the formation of the placenta. 4 The three layered embryo is now surrounded by proliferating extraembryonic mesoderm and the four extraembryonic membranes: The chorion completely surrounds the embryo and other membranes and functions in gas exchange. The amnion eventually encloses the embryo in a fluid filled cavity. The yolk sac encloses another fuid filled cavity and is the site of early blood cell formation. The allantois is incorporated into the umbilical cord and forms blood vessels to exchange gasses, nutrients and waste with the placenta. Gastrulation is followed by the organogenesis of the notochord, neural tube, and somites. Expanding region of trophoblast Amniotic cavity Epiblast Hypoblast Chorion (from Trophoblast) Yolk sac (from hypoblast) Extraembryonic membranes start Extraembryonic mesoderm cells (from epiblast) to form and gastrulation begins. Allantois Amnion Chorion Ectoderm Mesoderm Endoderm Yolk sac (no yolk!) Gastrulation has produced a three-layered embryo with four extraembryonic membranes. Extraembryonic mesoderm By the end of the first trimester, rudiments of all the major organs have developed.