Mitosis and Meiosis Lab: Cell Division & Reproduction
advertisement

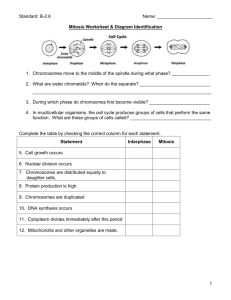
Mitosis and Meiosis LAB Group Members: Lab Station: Standard: AP Big Idea #3 and SB2ce Essential Question: How do eukaryotic cells divide to produce genetically identical cells or to produce gametes with half the normal DNA? Opening: Compare and contrast word wall activity for mitosis and meiosis. Background: Reproduction in living things can be achieved through either asexual or sexual processes. In asexual reproduction, offspring organisms are genetically similar to the parent organism with limited, if any, introduction of DNA from another individual. In sexual reproduction, offspring organisms are generated by combining one half of the DNA from each of two parent individuals to produce a diploid (2N) offspring that has two complete sets of chromosomes and genes. The process of sexual reproduction enables greater genetic variation in offspring than asexual reproduction. Multicellular, diploid, eukaryotic organisms use both types of cellular reproduction. Growth and development of the organism occurs through asexual reproduction of cells through the process of mitosis where every normal daughter cell is diploid, carrying two copies of each chromosome and set of genes. In mitotic reproduction, these daughter cells are genetic replicates of both the parent cell and the sister cell. Most single cell eukaryotes reproduce by simple mitosis, producing more replicates of the parent organism. Mitosis The mitotic cell cycle is a highly regulated process that insures accuracy in DNA replication and equal division into daughter cells. Defects in accurate reproduction will cause abnormalities that will result in decreased viability of both the daughter cells and the multicellular organism through a variety of mechanisms (including apoptosis and cancer). Mitosis is a continuous process that can be described by a sequence of stages. The stages are defined by molecular and cytosolic events. Visual inspection of dividing cells through the microscope has defined three main stages (Figure 1) that can be further subdivided (Figures 2, below figure 1, and figure 3 on the next page): Interphase: The cells look quiescent. Mitosis: The cells are separating DNA equally for two future daughter cells. Cytokinesis: The daughter cells are separated into two individual cells. Interphase In interphase, the cell may be arrested in this part of the cell cycle (referred to as the G0 part of interphase) as a diploid cell (2N). If the cell continues through the next mitotic cycle, interphase only appears to be quiescent. This is when the cell grows in preparation of another cycle (G1), DNA is replicated (S phase, cell becomes 4N), the DNA checked for errors and repaired prior to mitosis (G2). Mitosis is subdivided into four stages that can be identified through the microscope: prophase, metaphase, anaphase, and telophase. Prophase The replicated DNA condenses around histone proteins and supercoils to form visible chromosomes. • The nuclear membrane dissolves. • Centrioles form and begin to migrate to opposite sides of the cell. • Microtubules organize around the centrioles to form mitotic spindles • Spindle fibers attach to the kinetochore proteins at the centromere (holding sister chromatids together). Metaphase • Centrioles complete migration to opposite poles of the cell • Sister chromatids line up along the midline of the cell Anaphase • Spindles shorten, pulling sister chromatids apart to opposite poles of the cell Telophase • Chromosomes de-condense • 2 nuclear envelopes form around the separated chromosomes (each nucleus is 2N again) Cytokinesis Cytokinesis generally follows and divides the cytosol and cellular membrane to yield two cells that are identical. In plant cells, small vesicles move along microtubules to the mid line. These vesicles fuse to form a cell plate, which grows to form the cell wall that separates the cells. Control of the cell cycle To maintain normal growth and development it is essential that cells divide only when and where needed. Therefore, the transition between the stages of the cell cycle is a tightly controlled process. Levels of proteins called cyclins build up in the cell and bind to cyclin dependent kinase, forming CDK complexes. These CDK molecules either add or remove phosphate groups from substrates to move the cell to the next stage of the cycle. In order to progress, however, the cell must pass through several “checkpoints.” These checkpoints assure that cells only divide when needed and when DNA duplication is completed and without errors. The three main checkpoints are: Restriction Point – this occurs in G1. After division the daughter cells will continue to grow in size for a short time but are halted at the restriction point. Here cells will either terminally differentiate and enter what is termed G0, or the cells will be stimulated a by growth factor and continue through the cycle again. G2/M Checkpoint - occurs between the end of G2 and the beginning of mitosis. Specialized molecules read the newly formed DNA and will delay the cell from entering mitosis if there are strand breaks or if inappropriate nucleotides are incorporated. The cell cycle will only continue if repairs can be made, otherwise, the cell will die without completing the cycle. Metaphase/Anaphase Checkpoint - early in mitosis. Specialized proteins on each centromere, called the kinetochores, activate if microtubules are attached and appropriate force is being applied. Activation allows the chromosomes to separate. If activation does not occur, mitosis will halt and the cell will die without completing the cell cycle. Mutations in cells that lead to the loss of cell cycle control result in a loss of control over growth, cell differentiation, and death. Mutations may accumulate over time causing the cell clones to become a tumor; the tumor may later become metastatic if the cell acquires the ability to establish separate masses in distant tissues. These cancerous mutations usually affect genes which encode proteins involved in the control of other genes (transcriptional regulation), cellular metabolism (speeding up or slowing down cell growth), and/or the cell cycle of division and growth. Also affected are hormones and their receptors, regulatory molecules such as cytokines and their receptors, and DNA repair mechanisms. Meiosis Sexual reproduction occurs through the production of gametes which contain only one set of genes (haploid, 1N). When two gametes fuse, a new diploid organism with a different complement of gene copies than either parent is produced. Haploid gametes are produced through the process of meiosis. Meiosis consists of one round of DNA replication followed by two nuclear divisions, meiosis I and meiosis II (See the figure to the right). This results in the formation of four daughter cells, each with only half the number of chromosomes of the parent. When two gametes combine during fertilization to form a zygote, the diploid chromosome number is restored in the resulting organism. Typically, we think of gametes as cells that come from either a male (sperm) or a female (ova) that are specialized to fuse with each other in species specific way (mouse sperm cannot fertilize a whale ovum). Under a microscope, mitosis and meiosis do not look very different. The phases of division are visually distinct and named as their visual look-alikes in mitosis. However, the way replicated DNA is divided is very different in meiosis than mitosis. Prophase I: The duplicated sister chromosomes condense and are attached at a centromere as in mitosis. Unlike mitosis, these sister pairs form a tetrad with duplicated chromosome homologs from both the maternally and paternally contributed chromosomes. At this point DNA may cross over between non-sister, chromatid homologs forming unique chromatids that contain DNA from both maternal and paternal chromosomes. Metaphase I: The tetrads align at the center of the cell and spindle microtubules attach to the kinetochores. Anaphase I: The tetrads separate into two, duplicate chromosomes that remain attached at the centromere which move to separate poles of the cell. Anaphase I is followed by telophase 1 and cytokinesis, resulting in two daughter cells that each have 2N DNA. Mitosis II follows immediately after meiosis I is completed without further duplication of the DNA. The single set of chromosomes (held together at the centromere) are separated on the second spindle thus forming daughter cells that are now 1N. A starting diploid germ cell has now formed 4 haploid gametes, each containing a mixture of DNA from both the mother and the father. This method of division produces gametes that vary greatly between each other with respect to the allele combinations available to pass to the next generation. The example of meiosis that will be used in this investigation is Sordaria fimicola. S. fimicola is an ascomycete fungus that is haploid for the bulk of its life cycle; the haploids comprise the individual fungal filaments, called hyphae, which normally exist in a mass, called a mycelium, representing the “body” of the fungus, and the ascospores, from which mycelia develop. The only diploid portion of the life cycle of S. fimicola occurs when the nuclei of specialized hyphae come together. These hyphae fuse to form a diploid zygote. This zygote then undergoes meiosis to produce the haploid ascospores, yielding four haploid nuclei contained in a sac called an ascus. After meiosis I and II, the four haploid nuclei undergo mitosis, resulting in an ascus containing eight haploid ascospores from one diploid zygote. Many asci form inside a fruiting body called a perithecium. Wild types sordaria are a dark brown, but we will grow this cultures in the presence of a tan mutant so that when mycelium from a wild type fuses with the tan to form a zygote, we will be able to observe the results of crossing over events that occurred as the zygote underwent meiotic division. Figure 6 shows a culture plate that was seeded with two squares of wild type sordaria containing agar and two squares of tan type sordaria containing agar. The mycelium grow out from these blocks so fusion will occur where the two types meet. This region of fusion is where you will find perithecium containing ascospores from zygotes that came from a wild type and a tan parent (arrows). After meiosis I and II, the haploid ascospores will be either tan or wild type. When the parathecium are gently ruptured, the ascospores will be in strings of 8 that can easily be scored as tan or wild. How these ascospores are arranged within the ascus is a direct representation of whether or not crossing over has occurred between the centromere and the site for the gene for ascospore color. If no crossing over has occurred, the ascospores will be arranged in a 4:4 manner. If crossing over has occurred, they will occur in a 2:4:2 or 2:2:2:2 manner(s). Meisosis with crossing over By observing the ascospore arrangement, the percentage of asci exhibiting crossover can be determined. This frequency appears to be affected largely by the distance from the gene to the centromere. From the crossover frequency, the distance in map units from the gene for ascospore color, and the chromosome centromere, can be calculated. Pre-lab Questions: 1. Explain the difference between mitosis and binary fission. 2. Explain what crossing over is, when it occurs, and why it is important. 3. How is the cell cycle different for animals and fungi? 4. How do mutations affect the cell cycle? 5. When do checkpoints occur during the cell cycle? What are the purpose of checkpoints during the cell cycle? 6. Summarize how mitosis and meiosis different and similar? 7. Explain the various ways the cell cycle controlled? 8. How is cell division different for plant and animal cells? Part 1 - Materials 1 bag of: 40 red, 40 pink, 4 clear centrioles, 4 yellow centromeres Part 1 – Procedure (write your descriptions in the table provided at the end of the procedure section) To simulate Interphase 1. Construct two strands of seven red pop beads and attach each strand to a yellow centromere. Repeat with two strands of seven pink pop beads and a yellow centromere. These represent a homologous pair of chromosomes (red from the father and pink from the mother). 2. Picture an imaginary boundary in the center of your desk. This boundary represents the nuclear membrane. Place the chromosomes in the center of the imaginary nucleus. 3. DNA replication occurs, producing a duplicate of each chromosome. Construct two chromosomes identical to the ones you made previously. Each of the duplicated chromosomes is called a chromatid. Join both red chromatids at the centromere to form a pair of sister chromatids. Repeat this process for the pink chromosome. 4. Place a pair of plastic centrioles at the ninety degree angle, just outside of your nuclear membrane. The centrioles also replicated during interphase, so place another pair next to them in your cell. 5. Illustrate your simulation of interphase and in your own words provide a brief description of what happens during interphase. To simulate Prophase 6. Move your two pairs of centrioles to opposite poles (sides) of the cell (your desk). 7. Illustrate your simulation of prophase and in your own words provide a brief description of what happens during prophase. To simulate Metaphase 8. Center your chromosomes along an imaginary metaphase plate with the centrioles still at the opposite poles of the cells. 9. Illustrate your simulation of metaphase and in your own words provide a brief description of what happens during metaphase. To simulate Anaphase 10. Separate and move the centromeres of each chromosome toward opposite poles of the cell. Notice how the arm of each chromosome trails the centromeres to the poles. 11. Illustrate your simulation of metaphase and in your own words provide a brief description of what happens during anaphase. To simulate Telophase and Cytokinesis 12. Move one red strand and one pink strand to the centrioles they were heading toward during anaphase. Imagine a cleavage furrow developing between each nuclei and separating the cell into two daughter cells. 13. Note how each cell now contains one red and one pink chromosome, as well as one pair of centrioles, exactly like the cell with which you began. 14. Illustrate your simulation of metaphase and in your own words provide a brief description of what happens during telophase/cytokinesis. Interphase Illustration Description Prophase Metaphase Anaphase Telophase/Cytokinesis Part 2 – Procedure: Mitosis using prepared animal and plant slides 1. Obtain a compound microscope, the animal cells slide and the onion mitosis slide. 2. Observe the prepared microscope slide of onion root tip first at 100X (10X objective and 10X ocular) then at 400X (40X objective, 10X ocular). Look for cells in mitosis. Depending on the quality of your microscope you may be able to distinguish the various phases of mitosis. Use oil immersion if available. 3. Observe the prepared microscope slide of the animal mitosis first at 100X, then 400X. Look for cells in mitosis and classify the stages. Compare and record similarities and contrast differences between the animal and plant cell mitosis. 4. Examine at least three fields of view of the apical meristem of the onion root tip at 400X. In each view, count and record the number of cells in interphase and the various stages of mitosis. 5. Calculate the percentage of total cells counted in interphase and in each stage of mitosis. If the individual phases cannot be discerned, calculate the percentage in interphase and in mitosis. Table 1. Number of cells in each mitotic stage. Stage # of cells in % total number onion slide of cells Time (minutes in each stage) # of cells in animal slide % total number of cells Interphase Prophase Metaphase Anaphase Telophase Assuming that it takes an average of 24 hours (1,440 minutes) for onion root tip cells to complete the cell cycle, calculate the amount of time cells spend in each phase of the cycle. Use the formula provided below. Enter your results in Table 1. % of cells in phase x 1,440 minutes = _______ minutes cell spent in phase 7. Make a pie chart representing the amount of time spent in each stage of mitosis. Title and label all portions of the pie chart below. Prophase is normally the longest phase of mitosis, due to the complexity of the events occurring during prophase. These complex events take a relatively long time for the cell to perform:chromatin condensing and thickening, nuclear membrane dissolving, and the early stages of spindle development. Part 3 – Procedure: Root Tip Squash 1. Obtain some garlic root tip. Blot as much excess water from the root tips as possible. Any excess water on the slide will affect your results. Do not allow the root tips to dry out, however. 2. Using a scalpel, cut off the end of one of the emergent root tips; the section should be approximately 1 to 2 mm long. Place the root tip on a clean microscope slide and apply two or three drops of hydrochloric acid (HCl) to the root tip. 3. Holding the slide with a clothespin, pass it through the flame of a Bunsen burner for five seconds (five passes over the flame). Pass the slide through the flame of the Bunsen burner. Do not hold the slide directly in or over the flame. 4. Without harming the root tip, blot the specimen with a paper towel to remove the excess HCl. You may wish to touch a corner of the paper towel to the drop on the slide and allow the paper towel to soak it up. This may not remove the liquid from the slide as effectively as blotting, but it will not disturb the root tip. 5. Add a few drops of carbol fuchsin stain, covering the root tip. 6. Pass the slide through the flame of the Bunsen burner approximately 10-15 times. Let the slide stand for one minute. Pass the slide through the flame of the Bunsen burner. Do not hold the slide directly in or over the flame. Without disturbing the specimen, use a paper towel to remove the excess stain. 7. Cover with a coverslip. Using a pencil eraser or other blunt instrument, gently press down on the coverslip to squash and spread out the root tip. Blot off the excess stain, if any, that may have come out from under the coverslip. 8. Observe the slide under a microscope at 100X. Locate the apical meristem. Examine the slide at 400X. Locate cells in the various stages of mitosis, and make sketches of what you find. Table 2. Number of cells in each mitotic stage of the root tip. Stage # of cells in field # of cells in field # of cells in field 1 (directly at the 2 (middle of root 3 (opposite end tip) tip) of root tip) Interphase Prophase Metaphase Anaphase Telophase Field of View Images for each magnification observed (10, 40, 100x) Total # of cells % total number of cells Part 4 – Application: Environmental Effects on Mitosis Scientists reported that a fungal pathogen, may negatively affect the growth of soybeans (Glycine max). Soybean growth decreased during three years of high rainfall, and the soybean roots were poorly developed. Close relatives of R. anaerobis are plant pathogens and grow in the soil. A lectin-like protein was found in the soil around the soybean roots. This protein may have been secreted by the fungus. Lectins compounds, such as phytohemagglutinin, induce mitosis in some root apical meristem tissues. In many instances, rapid cell divisions weaken plant tissues. You have been asked to investigate whether the fungal pathogen lectin affects the number of cells undergoing mitosis in a different plant, using root tips of onions. Based on the information above create an experimental hypothesis (indicating whether you would predict onion roots to be affected in a similar manner to the soybeans having increased mitotic divisions or not). Experimental Hypothesis (Ha): _______________________________________________________________________________________________________ _______________________________________________________________________________________________________ Now write the null hypothesis (Ho) (this is the opposite of your experimental hypothesis): _______________________________________________________________________________________________________ _______________________________________________________________________________________________________ Root tips of onions were grown in both lectin exposed soil and soil will no lectin to determine the number of cells undergoing mitosis. Using the experiment described, identify the following variables: Independent variable: Dependent variable: Control: The data below is representative of number of cells seen in interphase, which would look like a dark condensed region in a cell and the combined number of cells in any of the phases of mitosis(PMAT). Table 1. Table of observed (o) number of cells for the onions treated with lectin and without lectin. Interphase Mitosis Total Control – no lectin 148 25 173 (A) Treated with lectin 161 88 249 (D) Total 309 (B) 113 (C) 422 (N) Below is a table for expected (e) values for onion root cells exposed to lectin and root cells not treat with lectin. You must complete the table for all the expected values. You will need to use the equations provided while referencing the variables indicated in table 1 ( A,B,C,D, and N) Table 2. Table of expected (e) number of cells for the onions treated with lectin and without lectin. Interphase Mitosis Control – no lectin (A*B)/N = (A*C)/N = Treated with lectin (D*B)/N = (D*C)/N = Table 3. Chi-Square Values for the onion cells treated with lectin and without lectin. Group Observed (o) value Expected (e) value (o-e) (o-e)2 (o-e)2 /e Control Interphase Control Mitosis Treated Interphase Treated Mitosis Total of (o-e)2 /e = chi square (X2) = _________________ Chi-square is a statistical test commonly used to compare observed data with data we would expect to obtain according to a specific hypothesis. For example, if, according to Mendel's laws, you expected 10 of 20 offspring from a cross to be male and the actual observed number was 8 males, then you might want to know about the "goodness to fit" between the observed and expected. Were the deviations (differences between observed and expected) the result of chance, or were they due to other factors. How much deviation can occur before you, the investigator, must conclude that something other than chance is at work, causing the observed to differ from the expected. The chi-square test is always testing what scientists call the null hypothesis, which states that there is no significant difference between the expected and observed result. That is, chi-square is the sum of the squared difference between observed (o) and the expected (e) data divided by the expected data in all possible categories. Once the Chi-square value is calculated the researcher needs to figure out if the value supports or rejects their null hypothesis. Table 4, below, indicates the critical values in which you will compare your chi-squared value. The degrees of freedom (df) equals the number of treatment groups minus one multiplied by the number of phase groups minus one. In this case, there are two treatment groups (control, treated) and two phase groups (interphase, mitosis); therefore df = (2-1)(2-1) =1. The p value is 0.05, and the critical value is 3.84. If the calculated chi-square value is greater than or equal to this critical value, then the null hypothesis is rejected. If the calculated chi-square value is less than this critical value, the null hypothesis is not rejected. Table 4. Critical Values of the Chi-Square Distribution Probability 0.05 0.01 0.001 Analysis Questions: 1 3.84 6.64 10.8 2 5.99 9.21 13.8 Degrees of Freedom (df) 3 7.82 11.3 16.3 4 9.49 13.2 18.5 5 11.1 15.1 20.5 1. Based on the data is the null hypothesis accepted or rejected? 2. Explain what it means if your null hypothesis is rejected. 3. What other experiments should be performed to verify your findings? 4. Does an increased number of cells in mitosis mean that these cells are dividing faster than the cells in the roots with a lower number of cells in mitosis? Part 5 – Crossing Over in Sordaria Refer to the introduction for information on the life cycle of Sordaria and crossing over. Class sets of data and calculations for Sordaria crossing over is located in table 1. A cross was made between wild type (+, black in color) and tan strains. The resulting zygote produces either parental type asci, which have four black and for tan spores in a row (4:4 pattern) or recombinant asci, which do not have this pattern (2:2:2:2, 2:4:2). By studying the frequency of crossing over, you can gather information that lets you draw a map of the relative location of genes on a chromosome. A map unit is a relative measure of the distance between two linked genes, or between a gene and the centromere. The greater the number of crossovers, the greater the map distance as seen in genes D-E and E-F in the diagram to the right. One map unit equals one recombinant per 100 total events. The percentage of asci showing crossover divided by 2 equals the map units in this activity. Each crossover produces two spores like the parents and two spores that are a result of the crossover. Thus, to determine the number of crossovers, you must divide the number of asci counted by two since only half the spores in each ascus result from crossing over. Table 1. Number of Asci indicating crossing over events. Number of Asci Number of Asci 4:4 pattern Crossing over Total number of Asci %Asci showing crossover /2 Gene to Centromere distance (Map Units) 425 51/2 = 25.5% 26 1st period 6th period 208 217 Analysis Questions: 1. Explain why we divide the percentage of asci showing crossing over (recombinant) by 2. 2. The published map distance between the gene and the centromere is 26 map units. How did the class data compare with this distance? 3. What can account for disparities, if any, between the class data and the published data? If there were no disparities, why might that be? 4. Can the same (or any) environmental factors you tested above affect the amount of crossing over that occurs in Sordaria? How would you set up an experiment to test this? For example, how does humidity or pH affect the crossover frequency? 5. Knowing that there is a single gene that determines the spore color for Sordaria, use the data from your class to indicate how far the gene is away from the centromere using the blank chromosome below. 6. Calculate the map units between gene and centromere for the small sample of Sordaria asci given below. Number of Asci Counted Spore Arrangement 7 4 light/4 dark spores 8 4 dark/4 light spores 3 2 light/2 dark/2 light/2 dark spores 4 2 dark/2 light/2 dark/2 light spores 1 2 dark/4 light/2 dark spores 2 2 light/4 dark/2 light spores 7. Chi-squared value Asci Configuration Observed (o) Expected (e) ((o-e)2)/2 4:4 wild:tan 208 Crossing over 217 totals 425 X2= Create your hypothesis below. Anytime something is known, such as Mendels laws, that is the null hypothesis. By doing the experiment you are trying to disprove the null or say that the law or theory is incorrect. Remember when calculating the expected value you must multiply the expected map unit percent by 2 since you divide by 2 calculating the map unit value. In other words, 26x2=52 so .52x425= the expected value for crossing over. H0- The map units for the Sordaria gene for color is 26 map units from the centromere. Ha- ________________________________________________________________________________ 8. Does the data hold true to what scientists have come to accept for the gene distance to centromere? Explain why. Part 6 – Loss of cell cycle control in cancer. Background questions: 1. How are normal cells and cancer cells different from each other? 2. What are the main causes of cancer? 3. What goes wrong during the cell cycle in cancer cells? 4. What makes some genes responsible for an increased risk of certain cancers? 5. Do you think that the chromosomes might be different between normal and cancer cells? Many of us have family members who have or have had cancer. Cancer can occur when cells lose control of their cell cycle and divide abnormally. This happens when tumor suppressor genes, such as p53 or Rb (retinoblastoma), are mutated. The last background question is the focus of this part of the lab. With your group, form a hypothesis as to how the chromosomes of a cancer cell might appear in comparison to a normal cell and how those differences are related to the behavior of the cancer cell. For each of the following cases, look at pictures of the chromosomes (karyotype) from normal human cells. Compare them to pictures of the chromosomes from cancer cells. For each case, count the number of chromosomes in each type of cell, and discuss their appearance. Case 1: HeLa Cells HeLa cells are cervical cancer cells isolated from a woman named Henrietta Lacks. Her cells have been cultured since 1951 and used in numerous scientific experiments. Henrietta Lacks died from her cancer not long after her cells were isolated. Lacks cancer cells contain remnants of human papillomavirus (HPV), which we now know increases the risk of cervical cancer. To the right is a karyotype of a HeLa cell. 1. What differences do you observe between the normal karyotype and that of the HeLa cell? 2. Explain how cancer cells could end up looking like this. Case 2: Philadelphia Chromosomes In normal cells, mitosis is usually blocked if there is DNA damage. Sometimes, though, DNA damage makes cells divide more often. Certain forms of leukemia have a unique feature called a Philadelphia chromosome. Look at the karyotype of leukemia cells in diagram to the right. 1. How do cells monitor DNA integrity? 2. How are the chromosomes in this cell, with leukemia, different than that of chromosomes in normal cells? 3. How might these differences lead to cancer?