Introduction 3
advertisement

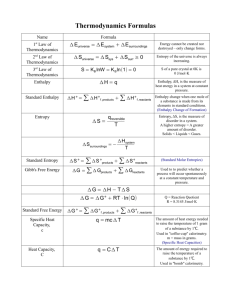
A few organizational items • Tentative topic for research project (paper) is due Feb. 1 (this week, okay) • Outline of research project is due March 9 (before spring break) • We are going to have a pizza & movie night, when would be a good date/time? • We’ll put books on reserve. • Graduate students: Please remember presentation, choose own paper, or paper from paper list on web site Introduction-3 Thermodynamics Homework 3 1. (due Mo, Feb. 6): 2. 3. 4. Consider the isomerization reaction of Dihydroxyacetone phosphate (DHP) to Glyceraldehyde 3-phosphate (GP) which occurs in glycolysis. At equilibrium the ratio of GP to DHP is 0.0475 at 25C and pH 7 (standard conditions). (a) What is the standard free energy change for this reaction? (b) If the initial concentration of DHP is 2 x 10-4 M and that of GP is 3 x 10-6 M, what is DG for that state? Is it higher or lower than the value in (a)? In which direction will the reaction proceed (toward DHP or GP)? van Holde, Problem 2.5 van Holde, Problem 2.7 Van Holde, Problem 2.8 (Solutions to odd-numbered problems are in back of book). Reading: Van Holde, Chapter 1 Van Holde Chapter 3.1 to 3.3 Van Holde Chapter 2 Van Holde Chapter 4 (cursory reading) Paper list (for presentations) is posted on web site Introduction-3 Thermodynamics • In this section, we are asking, in which direction will a biochemical reaction proceed?. • This will depend on two quantities: 1. The enthalpy, DH, of the reaction. This corresponds to the sum of all the potential energies we discussed in Introduction-2. 2. The entropy, DS of the reaction. This relates to the number of ways the system can adopt. • In other words, it will depend on the Gibbs’ free energy, DG of the reaction: DG DH TDS 0 0 0 Enthalpy Definition: H = E +PV (is a state function) • Why is it useful? – The enthalpy change between the initial and final states of a biochemical process, DH, is the easily measured heat that it generates or absorbs. – H is a state function. DH of a reaction only depends on initial state and final state of reaction; it does not matter what goes on between. Example: Measuring enthalpy of oxidation of glucose to CO2 and H2O directly in muscles would be very difficult. But, since enthalpy is a state function, we can measure the enthalpy of this reaction in any apparatus (e.g. a constant pressure calorimeter) and get the same result. We don’t even need to know the reaction mechanism, as long as we know the beginning and end state. Most biochemical processes occur in liquids or solids (rather than gases), so volume changes are small. To a good approximation, we can often neglect the difference between DH and DE in biochemistry and simply talk about the change in ‘energy’ accompanying a reaction. This is the potential energy we discussed in Introduction-2. Entropy • Still, the enthalpy alone cannot tell us if a reaction occurs spontaneously (by itself). • Two examples: – When two blocks are brought in contact, heat flows from the hot one to the cold one, never vice versa (‘reaction’ occurs, but enthalpy of system stays the same). – Two bulbs of equal volume connected by a stopcock. All molecules are on one side first; when the stopcock is opened, molecules diffuse back and forth, until they are equally distributed (‘reaction’ occurs, but enthalpy of system stays the same). Total number of states: 2N. The number of ways W of putting L of the N molecules into the right bulb is: N! WL L!( N L )! Most probable state: highest value of WL Here: L = N/2. Entropy In chemical systems, the number of ways, W, of arranging a system in a particular state is huge. Define entropy of a system: S kB lnW kB … Boltzmann constant kB = 1.38*10-23 J/K S is function of state For gas bulb example (previous slide): Entropy of the system is S = kB*lnW = kB*ln2N = kB*Nln2 Entropy of all molecules on one side: WN = 1 S=0. Entropy of having N/2 molecules in each bulb is largest. The laws of random change cause any system of reasonable size to spontaneously adopt its most probable arrangement, the one in which entropy is a maximum, simply because it is so overwhelmingly probable. Once the most probable state has been reached, the system stays there (macroscopically) and is said to have reached equlibrium. (Here: Assume all states have same energy) Boltzmann’s grave, Vienna Example: A certain 100 amino-acid long polypeptide chain has only one alpha-helical conformation but there are three possible orientations for each residue in the random-coil state. 1. Calcuate DS for the conformational change Random coil alpha helix. 2. Does the entropy increase or decrease for this transition? Examples of entropy • Quasi-reversible heat transfer (const Temp): q DS T • Isothermal, reversible expansion of gas from V1 to V2 (p. 84): V2 DS nR ln V1 • Isothermal dilution of a solute from concentration C1 to C2 (p. 85): • Entropy of mixing (p. 86): Xi mole fraction of species i n… number of moles of species i C1 DS nR ln C2 DS R ni ln X i i Systems at constant temperature and pressure (most biochemical systems) • Define Gibbs free energy: G = H – TS – Then dG = VdP – SdT – For const. P & const. T systems (dP = dT = 0): – dG = 0. What does this mean? G must be an extremum (minimum) for such a system to be at equilibrium!! Gibbs free energy • Is of enormous importance in deciding the direction processes & equilibrium positions in biochemical systems • If DG for a particular process is negative, that process is spontaneous, because it leads in the direction of equilibrium. DG DH TDS 0 0 0 DH and DS are equally important Energy minimization and entropy maximization play a part in determining the position of equilibrium. Gibbs free energy DG DH TDS 0 0 0 DG RT ln Keq 0 Gibbs free energy The native and denatured forms of a protein are generally in equilibrium. Native Denatured For a certain protein, (total conc 2.0 * 10-3 M) the concentration of the denatured and native forms at 50°C and 100°C is given in the table. Temp Denatured (M) Native (M) 50 5.1*10-6 2.0*10-3 100 2.8*10-4 1.7*10-3 1. Determine DH and DS for the folding reaction (assuming they are independent of T) 2. Calculate DG for this protein at 25°C. Is the folding process spontaneous? 3. What is the denaturing temperature for this protein at standard conditions? Van’t Hoff Plot DG 0 DH 0 TDS 0 ln Keq RT RT ln K eq 1 0 DH DS R T Van’t Hoff equation Keq and DG0 are temperature dependent Enthalpy can be calculated from slope; Measure Keq as a function of T: ln Keq Then get DS from van’t Hoff equation Slope: -DH/R DH and DS are tempindependent over small temp ranges. 1/T (1/K) Gibbs free energy in real life • For real biochemical reactions we need to consider DG for the object under study (e.g. protein, reaction, etc) AND DG for the solvent (usually water) • Water molecules form Hydrogen bonds (enthalpy). • “Fixing” water molecules will “cost” decrease entropy of system (entropy). DGtotal DGreactants DGwater Application of Gibbs free energy to protein and DNA stability • Need to consider enthalpy part of protein/DNA. These are the potential energies we discussed. Bonding potential, Hydrogen bonds, chargecharge interactions, dipole-dipole interactions, van-der-Waals, etc. • Need to consider entropy part of protein/DNA. Folded protein has one conformation (low entropy) and unfolded protein has many conformations (high entropy). • Need to consider enthalpy of water. Water forms many H-bonds. • Need to consider entropy of water. • Need to consider enthalpy of ions in solution (charge-charge interactions) • Need to consider entropy of ions in solution (binding (fixing) will lower entropy). Hydrophobic effect • Perhaps most important contribution to protein folding. • Hydrophobic (non-polar) substances don’t want to “touch” water. – hydrophobic residues are on protein inside – bases are on DNA inside (base-pairing (Hbonds), don’t contribute much) Hydrophobic effect It is an entropy effect: Transfer of hydrophobic residues from water to non-polar solvent (e.g. benzene) Often a small DH, but a large, favorable DS component. Why? Sticking a hydrophobic substance into water, makes the water form a fixed cavity around the substance This “costs” (decreases) the entropy of the water. Hydrophobicity determines placement of amino acid in protein (related to protein folding). In aqueous environment, hydrophobic residues hide inside; (this is reversed in the membrane). Hydropathy – feeling about water; hydrophilic – likes water; hydrophobic – does not like water. DGtrans.vap water aq RT ln P , where P , mole fraction nonaq Kyte-Doolittle Scale: They used vapor to water, others have used ethanol to water. In this equation: If Xaq > Xnonaq then DG is negative – hydrophilic If Xnonaq > Xaq then DG is positive – hydrophobic e.g. DGtransfer for val is 2.78, DGtransfer for Glu is - 8.59 (in Kcal/mole) Non-polar aa aa Kyte and Doolittle actually used combination of: 1. 2. 3. 0.69*DGtransfer + 2.32 48.1*(fraction 100% buried) – 4.5 16.45*(fraction 95% buried) – 4.71 They combined these three things to get a hydropathy index that ranges from +5 (very hydrophobic) to -5 (very hydrophilic). aa aa aa aa aa aa aa aa H2O aa aa