The discovery of the past
advertisement

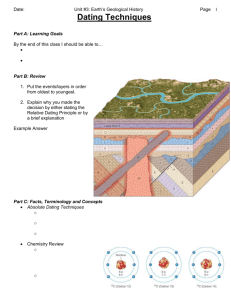
The discovery of the past To study evolution means to dig in the past. The science of past organims is paleontology (greek: palaews: old, logos: science) Paleontology deal with fossils (lat. fodere = to dig) Charles Lyell Georges Cuvier Early paleontology mainly described ancient life within the Linnean framework Modern paleontology tries to reconstruct ancient life. It links therefore ecology and taxonomy. Mary Anning (1799-1847) Richard Owen (1804-1892) How do animals fossilize? Taphonomy (Greek: tafos: burial; nomos: law) Immediate burial Death Living organism Remains Mineralization Fossil Buried remains Decomposition Decay Bleaching Delayed burial Exposed remains Ginkgo biloba Stratinomy Ginkgo adiantoides Much less than 1% of all organisms fossilize Coral fish Coral fish from Jura Bioerosion A fossil forest in Dorset, England formed by fossilized bacteria around old tree stumps. A fossilized dinosaur footprint from New Mexico Fossilized Cyanobacteria (stromatolites) from South Africa A mammoth coprolith (fossilized excrements) What fossilizes? Hard body materials Substance Examples Calcite (CaCO3) Octocorallia Bryozoa Brachiopoda Polychaeta Ammonita Belemnita Echinodermata Aragonite (CaCO3) Hydrozoa Gastropoda Calciumphosphate (Ca5(OH)(PO4)3) Vertebrata Trilobita Crustacea Opal (SiO2.H2O) Radiolaria Diatomea Porifera Chitin Algae Fungi Arthropoda Cnidaria Priapulida Annelida Cellulose Plantae Tunicata Soft body materials Soft tissues very seldom fossilize (of about half of all major evolutionary lines no fossils are known) Exceptions are Fast drying out in very arid climates Permanent frozen Preservation in amber or asphalt A feathered Dinosaur: Sinosauropteryx Under what conditions do organisms fossilize? Moisture gradient Nutrient rich soils River sediments Anaerobic conditions (moorlands) Volcanic ashes Probability of fossilization Salinity gradient How complete is the fossil record? 0.7 0.6 0.5 0.4 0.3 0.2 0.1 0 SCI Divergence time inferred from cladogram GER Divergence time inferred from fossils RCI SCI: Quotient of consistent to inconsistent nodes 0 5 10 15 20 25 30 PZ PZ/MZ NZ MZ/CZ CZ Older -----Younger RCI: Relative completeness index GAP: Gap excess index 100% Completeness Metrics Benton MJ, Willis MJ, & Hitchin R. 2000. Quality of the fossil record through time. Nature 403: 534-537. ? At the family level about 50% of all taxa are known from fossils. 50% 0% Species Family Order Class Taxonomic levels Type Fossils of soft-bodied types are not well known Continental drift Alfred Lothar Wegener (1880-1930) The tectonic plates (from David Sanfwell, Scripps Inst. Oceanography) Evidence for plate tectonics: Fit of coastlines Distribution of mountains Continuity of fossils Continuity of geological features Isostasy: Earth acts like a fluid From Press et al.. 2004. Understanding earth, http://www.whfreeman.com/presssiever/con_index.htm?99iex Continental drift From C. R. Scotese: http://www.scotese.com/future.htm How to match phylogeny and plate tectonics Cassowary 35 62 Emu New Guinea 0.1 65 Kiwi Australia 55 69 Ostrich 82 South America 79 Rhea 100 New Zealand Moa Africa Tinamou Fossil dating Relative dating methods Relative dating uses geological strata to infer whether fossils are older or younger than a given stratum Layer 1 Younger Layer 2 Time Layer 2 Older Stratigraphy Morphological primitivism Absolute dating methods Radiometric absolute dating Radioactive Element Potassium 40 Rubidium 87 Thorium 232 Uranium 235 Uranium 238 Carbon 14 Stable element Argon 40 Strontium 87 Lead 208 Lead 207 Lead 206 Nitrogen 14 Half time 1.25 billion yrs 48.8 billion yrs 14 billion years 704 million years 4.47 billion years 5730 years Most minerals that contain radioactive isotopes are contained in igneous rocks. The dates they give indicate the time the magma cooled. Potassium 40 is found in: potassium feldspar (orthoclase) muscovite amphibole glauconite Surviving atoms C14 Daugther atoms N14 Volcanic rocks Sometimes in sediments Uranium may be found in: zircon urananite monazite apatite sphene Volcanic rocks Carbon 14 is used for bones Radiometric dating Stratigraphy Raw data Relative time scale Absolute time scale Recognition of unique events to subdivide time Geological time scale Post eruption 2 time Depth [m] Volcanic ash 2 Calibrating geological time Radiomtric dating of layers Post fossil B time 160 ± 10 mya 165 mya Fossil B time Last occurrence of B: First occurrence of Pre fossil B time Volcanic ash 1 190 ± 8 mya Last occurrence of A: First occurrence of 180 mya Pre eruption 1 time Fossil B time Older than 190 mya Pre fossil B time Modified from Andy MacRae: Radiometric Dating and the Geological Time Scale. http://www.talkorigins.org/faqs/dating.html Fission track Fission Tracks (FT) are micrometer-sized, linear damage tracks that occur in insulating minerals and that are caused by the spontaneous fission of heavy, unstable nuclides (mostly 238U in natural minerals). Dendrochronology Dendrochronology analyses treering growth patterns. History of the earth Steno founded stratigraphy by stating that geological layers are horizontal and superposed. Deeper layers are older. Nicolas Steno (1638-1686) The Red Rock Canyon, California The geological time scale Eon Era Phanerozoic Cenozoic Mesozoic Paleozoic Proterozoic Neoproterozoic Mesoproterozoic Palaeproterozoic Archean Hadean Period Quarternary Neogene Paleogene Cretaceous Jurassic Triassic Permian Carboniferous Devonian Silurian Ordovician Cambrian Ediacaran (Vendian) Cryogenian Tonian Age at Base (Mya) Duration (Mya) 2 2 23 21 65 42 140 75 205 65 250 45 290 40 355 65 410 55 440 30 490 50 540 50 630 90 850 220 1000 150 1600 600 2500 900 3800 2950 4550 750 The reconstruction of phylogeny The first Darwinian principle told that every phylogenetic tree has one common ancestor. Phylogenetic analysis is the study of taxonomic relationships among lineages. Phylogenetic systematics Cladistics (greek κλάδος: branch) Willi Hennig (1913-1976) Numerical taxonomy Robert Sokal (1926-2012) http://www.faunaeur.org/ http://tolweb.org/tree/phylogeny.html http://www.eol.org/ The cladistic methodology A B adf ade e f C D abc c d e b a Ancestor abd Apomorphies are common derived characters. Autapomorphies are characters that are restricted to single lineages. Plesiomorphies are ancestral derived characters. e: Autapomorphy of lineage D b: Synapomorphy of lineage C+D d: Plesiomorphy of lineage A It is a symplesiomorphy a: Apomorphy of the whole tree It is the ancestral state. The collective set of plesiomorphies defines the ground plan of a phylogenetic tree. A B C adf ade abd e f C is the sister taxon of A and B Character d in lineages A, B, and C is not homologous because it derived twice. It is homoplasious d d Character a in lineages A, B, and C is homologous because it synapomorph b a Ancestor Monophyletic taxon Paraphyletic taxon B A e f Polyphyletic taxon C D f d b d Ancestor b E The ultimate aim of taxonomy is to group higher taxa into monophyletic subtaxa. For this task we have to infer autapomorphies Autapomorphy defines monophyly Tetrapoda The diversification of an evolutionary tree is called cladogenesis Actinopterygia Dipnoi Amniota Archosauria Anura Urodela Mammalia Squamata Aves Therosauria Loss of tail apomorph Mammae autapomorph Reptilia (paraphyletic) Feathers apomorph Amnion apomorph Common ancestor Tetrapod limbs apomorph Lungs plesiomorph The evolutionary change within a lineage is called anagenesis Linnean systematics and cladistics Linnean approach Hennigean approach Hierachical encaptive system Hierachical encaptive system Phenomenological method based on similarity Analytical method based on lineage branching It uses grades (groups of similar body plan) It uses clades (groups of identical root) Different taxonomies are possible Only one taxonomic solution is allowed There is no clear decision intrument for taxonomies Autapomorphies decide about taxonomic position The number of higher taxa is rather small The number of higher taxa is large (Pisces, Amphibia, Reptilia, Aves, Mammalia) (Pisces, Amphibia, Reptilia are not valid taxa ) It does not assume common evolutionary history It is based on common evolutionary history It does not reconstruct evolution It does reconstruct evolution Taxonomy is independent of evolution Taxonomy is a part of evolutionary theory Low resolution trees High resolution trees The construction of phylogenetic trees from numerical methods The principle of maximum parsimony (Occam’s razor) holds that we should accept that phylogenetic tree that can be constructed with the least number of morphological changes. The raw data Species A B C D E 1 1 1 0 0 1 2 1 1 1 0 0 Characters 3 4 0 1 1 1 0 0 1 1 1 1 5 1 1 1 0 0 6 1 1 0 1 1 A B D E C 001101 110111 101101 010010 8 changes Distance matrix Species A B C D E A 0 1 3 4 3 B 1 0 4 3 2 C 3 5 0 5 6 111111 D 4 3 5 0 1 E 3 2 6 1 0 We are looking for such a tree that minimizes the sum of distances. A Outgroup B D E C 001101 101101 010010 111111 How to define the root? 010111 110111 7 changes Parsimony analysis To find the most parsimonious tree we have to cross all combinations of lineages (trees) with all character combinations at the root. The number of possible trees Number of trees Species 2 1 3 3 4 15 5 105 6 945 7 10395 8 135135 9 2027025 10 34459425 N (2S 2)! 2S1 (S 1)! Assumption of the numerical methods Birds Characters (or transitions) have to be independent. Impossible character states have to be excluded. Fish Loss of hairs Mammals Loss of feathers Hairs Feathers Scales Characters are assumed to have equal importance. In reality transitions are not comparable. To overcome this problem you give character weights. Technically you multiply the occurrence of a character in a distance matrix Incompatible Trees from molecular data Distance matrix Species A B C D E Sequence A A C A C G C G G G T T T T T T T T G G A A T T T A A G G G C C G G C C C A A C C C A A C A A T T A A A G A A T T A A T A A C A A A B C D E A 0 1 11 10 5 B 1 0 10 9 5 C 11 10 0 3 9 D 10 9 3 0 6 E 5 5 9 6 0 Evolutionary time scales The molecular clock Numbers of amino acid substitutions and therefore trespective numbers of nucleotide substitutions are for many proteins and genomes approximately proportional to time. Motoo Kimura Emile Zuckerkandl Tomoko Ohta (1933-) (1924-1994) (1922-2013) 80 Hence, numbers of substitutions are a measure of time of divergence from the latest common ancestor. Substitutions alone provide a relative time scale 70 acid differences Nuumber of amino c c Linus Pauling (1901-1994) 60 50 40 30 20 Errors 10 0 0 200 400 600 800 1000 Paleontological divergence estimate Superoxide dismutase An appropriate calibration adds the absolute time scale Paleontological versus molecular timescales Molecular estimates point frequently much more ancient divergences of lineages than estimates based on the fossil record. The reason are different speeds of morhological and genetical changes. Changes in genetic constitution accumulate to a point where basic regulatory elements are involved First fossils of placental orders (65 mya) Eomaia (125 mya) Molecular divergence of placental orders (120-140 mya) Morphological change Gene flow up to 2 mya Genetical change Genetical change Time axis Changes in genetic constitution involve first basic regulatory elements. First fossils of erect hominids (6-7 mya) Morphological change Time axis Molecular divergence (4-5 mya) Paleontological versus molecular timescales z Matching of molecular and paleontological timescales in Echinodermata 250 estimate Molecular divergence 300 200 150 100 50 0 0 100 200 300 Paleontological divergence estimate For the majority of Echinoderm subtaxa molecular divergence estimates are higher than the paleontological estimates. Data from Smith et al. (2006) Paleontological versus molecular timescales Divergences Placental-marsupials Amniotes-amphibians Myriapods-chelicerates Mosses-vascular plants Crustaceans-insects Echinoderms-chordates Spiralian-Ecdysozoans Protostomes-deuterostomes Arthropods-chordates Cnidaria-bilaterians Sponges-chordates Data from Qun et al. (2007) Earliest fossil record 175–145 310 530 450 530 <530 560–540 560–540 560–540 <600 <600 Molecular estimates 185–161 375–345 705–579 899–515 726-539 1001–586 643–544 678–556 1200–588 724–615 1350–592 Today’s reading History of palaeontology: http://en.wikipedia.org/wiki/History_of_paleontology History of earth: http://wiki.cotch.net/index.php/History_of_the_Earth Radiometric dating details: http://www.tulane.edu/~sanelson/eens211/radiometric_dating.htm Geological time scale: http://en.wikipedia.org/wiki/Geologic_time_scale Phylogenetic systematics: http://evolution.berkeley.edu/evolibrary/article/phylogenetics_01 Cladistics: http://en.wikipedia.org/wiki/Cladistics Ernst Haeckel: Kunstformen der Natur (Internet exhibition of original drawings: http://caliban.mpiz-koeln.mpg.de/~stueber/haeckel/kunstformen/liste.html The modern molecular clock: http://awcmee.massey.ac.nz/people/dpenny/pdf/BromhamPenny_2003.pdf