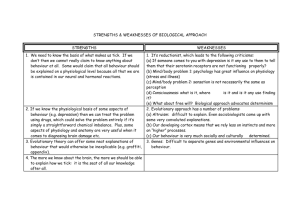
MSc Palaeoanthropology and Palaeolithic Archaeology: dissertation
advertisement

The neural substrate of religiosity and its emergence in the archaeological record Philip D Hawker Dissertation submitted in partial fulfilment of the requirements for the degree of MSc in Palaeoanthropology and Palaeolithic Archaeology of University College London in 2011 UCL INSTITUTE OF ARCHAEOLOGY Note: This Dissertation is an unrevised examination copy for consultation only and it should not be quoted or cited without the permission of the Director of the Institute The Theology of Bongwi the Baboon This is the wisdom of the Ape, Who yelps beneath the moon‘Tis God who made me in His shape, He is a Great Baboon. ‘Tis He who tilts the moon askew And fans the forest trees, The heavens which are broad and blue Provide him his trapeze; He swings with tail divinely bent Around those azure bars And munches to his Soul’s content The kernels of the stars; And when I die, His loving care Will raise me from the sod To learn the perfect mischief there, The Nimbleness of God. - Roy Campbel 1 List of contents Abstract ...……………………………………………….............................................................5 Preface: ……………………………………………......................................................................6 A brief historical background The paradoxes of religiosity Costly adherence Religious behaviour antedates the evolution of anatomically modern humans Acknowledgements…………………………………………………………………………..…11 1.0 Neural substrates of religiosity: an overview…..................................................................12 1.1 Genetic predisposition 1.2 Neurotransmitters 1.21 Dopamine 1.22 Seratonin 1.23 Endorphins 1.24 Summary of the role of neurotransmitters 1.3 Cytoarchitectural features: fossil/neuroarchaeological evidence 1.31 Temporal lobes 2.0 The contribution of Developmental psychology to an understanding of religiosity...........19 2.1 Common sense dualism in infants primes the brain for supernatural concepts 2.2 Children’s belief in gods is a default cognitive stance 2.3 Over developed sense of cause and effect in infants primes brains for God concepts 2.4 The sense of lacking control, increases illusory pattern perception in adults: Jennifer Whitson and Adam Galinsky 3.0 The contribution of Evolutionary Psychology to an understanding of religiosity……....22 3.1 Intuitive Psychology (Theory of Mind) 3.2 Thresholds of levels of intensionality 3.3 HADD (Hyperactive Agent Detection Device) 3.4 Minimally Counterintuitive Concepts 4.0 Adaptation or exaptation? ...................................................................................................26 4.1 Existential purpose generator 4.2 Causal explanation for phenomena 4.3 Religious belief is an adaptation to facilitate good physical and psychological health 4.31 Increased longevity 4.32 Attenuated experience of stress 4.33 Lower incidence of chronic physical ill health 4.34 Religious belief encourages risk-averse behaviour 4.35 Assortative sociality 4.36 Conclusion 4.4 Religious belief as a solution to the free rider problem 4.41 Early prosociality secured by kin-based altruism a function of Darwinian fitness 4.42 Prosociality secured by non-kin-based altruism as a function of inclusive fitness 2 4.43 Prosociality in early hunter-gatherers: non-kin based reciprocal altruism and the role of COMT 4.44 Costly signalling as a commitment to practices ensuring group health outcomes 4.45 Levels of inter-group competition a function of intra-group altruistic behaviour 4.46 Costly signalling behaviour most effective in a religious context 4.47 Types of costly signal in the archaeological record 4.48 Conclusion 5.0 Archaeological evidence for the presence of religious belief…………………………….39 5.1 Evidence of early ritualistic behaviour ca. 70,000 years ago 5.2 Mortuary practices 5.21 Middle Palaeolithic & Neanderthal burials 5.22 Burials of anatomically modern humans 5.3 Shamanism in Franco-Cantabrian parietal imagery in the Upper Palaeolithic 5.4 Parietal imagery exhibiting costly anatomical signalling 5.5 Anthropomorphous figurines 5.51 Pre Upper Palaeolithic evidence 5.52 Gravettian and Solutrean anthropomorphous figurines 5.53 Magdalenian anthropomorphous figurines 5.54 Neolithic figurines 5.6 Conclusion 6.0 Fossil evidence……………………………………………………………………………..56 6.1 Increased cranial capacity 6.2 Increase in EQ 6.3 Increased size of neocortex 6.4 Increase in the levels of intensionality 7.0 Discussion ………………………………………………………………………………...61 7.1 Distributed nature of religious cognition 7.2 Costly signalling does not support adaptationist argument 7.3 costly signalling underlies the pervasiveness of Palaeolithic religiosity 7.4 Distributed nature of religious cognition undermines the neurotheological programme 7.5 Weaknesses in argument 7.6 Limitations of analysis 7.7 Tentative conclusion which would be strengthened by further research 8.0 Summary and Conclusion …………………………………………………………..........69 Bibliography Appendices 3 List of Figures Fig. 1 Overview of thesis Fig. 2 Composite fMRI scan activation data (in red) shows brain sites affected by COMT gene type during a working memory task Fig. 3 The Reward System: Dopamine and Serotonin pathways in the brain (rodensor.com) Fig. 4 Robin Dunbar’s Thresholds of intensionality in the Homo lineage Fig. 5 The Löwenmensch figurine from Stadel im Hohlenstein Fig. 6 The operation of Hamilton’s Rule Fig. 7 The so-called “Python Stone” from Tsodilo, Botswana ca. 70,000ya Fig.8 The so-called “Python Stone” illuminated by night Fig. 9 Spearheads excavated from Tsodilo Fig. 10 The so-called “bear skull altar” at Chauvet Fig.11 Jean Clottes at the so-called “bear skull altar” at Chauvet Fig.12 Neanderthal burial at Kebarah Fig.13 Deliberate burial of an anatomically modern human ca. 90,000–100,000 B.P Fig. 14 Mortuary practices correlated with levels of intensionality (adapted from Stringer, 2011) Fig. 15 “The Sorcerer” from Chauvet (c. 30,000 BP) Fig. 16 Depiction of Shaman: Trois Freres, France, (upper Palaeolithic) ca. 13,000 BC Fig. 17 Distribution of hand prints featuring amputated fingers at Grotte de Gargas Fig.18 The Berekhat Ram Figurine: a false dawn? Fig. 19 Gravettian and Solutrian figurines and find sites Fig. 20 Late Magdalenian carved figurines and site locations Fig. 21 Necessary and sufficient conditions for effective costly signalling practice and examples of their signature in the archaeological record Fig. 22 Evolutionary trajectory of gross cranial capacity in the human phylogeny Fig. 23 Evolved human cranial morphology illustrating, expansion and growth of the neocortex Fig. 24 Brain mass as a percentage of body mass Fig. 25 Summary of research questions arising from this dissertation. Fig. 26 Overview of integration of disciplines in an explanation of the significance of the artefactual record in terms of costly signalling 4 Abstract: I will attempt to a) identify the neural correlates of theistic religiosity b) speculate on the selection pressures which facilitated the expression of supernatural agents in the context of costly signalling theory c) present the data which serve to illustrate its signature in the archaeological record. I shall, for the purposes of this dissertation, adopt the very narrow operational definition of religiosity which refers to propositional claims which refer to supernatural agents customarily characterised as gods or ancestors. a) I shall discuss three putative neural substrates: genetic preconditions; neurotransmitters and cytoarchitectural features, focusing on the role of neurotransmitters, in particular, dopamine and its inhibitor, catechol-O-methyltransferase. These seem critical in explaining the role of supernatural agency in early hunter gather groups. b) I shall attempt an overview of adaptive explanations for theistic belief in the literature before elaborating what I am convinced is the most plausible explanation which is its role in facilitating prosociality to cope with increased group size & external threats. This is facilitated by research suggesting that belief in gods is a default cognitive stance. c) I will present archaeological and fossil evidence which situates the emergence of religiosity chronologically and within the framework of costly signalling theory supported by ethnographic evidence. Overview of the main thesis Inhibition of dopamine receptors Makes possible deferred gratification Enables costly signalling (McNamara, P., 2006) Stimulated by interinter-group Conflict (Bowles, S., 2006) Helps solve freefree-rider problem Leads to parochial altruism Reinforced by theistic Beliefs/ancestor worship (McKay, R., et al, 2011) Fig. 1 Overview of thesis 5 Preface A brief historical background The pedigree of attempts to establish the existence of God from scrutinising the contents of the human mind is long (Anselm of Canterbury, 1077-1078; Descartes, 1641) but by the 18th century the ontological argument was being discredited by the work of Immanuel Kant (Kant, 1781). The recent popularity of so-called Neurotheology can be seen as an attempt to restock the theistic armoury in response to its depletion with the loss of the “classical” arguments for the existence of God. For the “neurotheologian”, neuroscience has been recruited to fulfil the task that philosophy has been unable to do. However, the results of any programme which attempts to establish such a causal relation between the neurological data and the ontological status of theistic propositions cannot be used to help prove or disprove the existence of putative gods. Many researchers in the field have been at pains to drawn the same conclusion: (Fingelkurts & Fingelkurts, 2009; Saver and Rabin, 1997). Methodology In addition to identifying the neural substrate of religiosity, I shall evaluate data drawn from research in developmental psychology which suggests that a teleological view of the natural world and a superstitious/theistic default state of mind which has a bearing in the emergence of religiosity in early hominins The persistence of a belief in life after death which underpins belief in supernatural agents is likewise revealed to be a default cognitive state. Much research by evolutionary psychologists demonstrates that so called “common sense dualism”, a “hyperactive agent detection device”, an innate theory of mind and other related features of intuitive psychology have primed the brain/mind of early modern humans for a belief in gods. I shall briefly review the more plausible anthropological explanations for the emergence of religiosity, but I shall place the greatest weight on costly signalling theory which strongly 6 suggests that, by enlisting the sacred in sanctions in facilitating prosocial behaviour, it constitutes an effective solution to the free rider problem faced by early human populations. I shall seek to demonstrate that the neural substrate of theistic cognition is distributed and not localised, and therefore unlikely to have been selected for. As Brown succinctly puts it, “…religiousness is clearly not like language and music with respect to specific behavioural outcomes.” (Brown, 2006, 232) Even if I am successful in identifying the neural substrate of religiosity, these features alone reveal little about the selection pressures which brought them about; there is, nevertheless, an observable archaeological residue of religious practices which can stand as proxy for the theistic religious beliefs of early modern humans: plausibly religious practices include deliberate burial belying a belief in life after death ,sometimes accompanied by grave goods; imputed religious symbolism in the portable art of the Upper Palaeolithic and typified by the so-called “Venus figurines”. Much has been made of the Franco-Cantabrian parietal “cave art” of the Upper Palaeolithic and the depiction of shamans and shamanistic practices though I am sceptical of the heuristic value of this ethnographic analogy. I shall deal with these in section 5. The paradoxes of religiosity Religious adherence demands costly signals of commitment which are often highly inimical to the reproductive fitness of the practitioner; there is a prima facie inconsistency with the operation of natural selection. Secondly, it would seem that the evolution of anatomically modern humans in sub-Saharan Africa predates by as much as one millennium the unambiguous occurrences of religious artefacts in the archaeological record. If the biological preconditions for human religiosity are a product of the genome established at this earlier date, how is the delay in its emergence in the archaeological record to be explained? 7 Costly adherence The widespread nature of religious belief was attested to by Darwin (Darwin, 1871) and as Bulbulia has mischievously put it, “God-mongering is a panhuman tendency.” (Bulbulia, 2004, 658) Its costliness both to the individual adherent and the group to which he or she belongs is delineated well by Bulbulia: ”…misperceiving reality as phantom infested, frequent prostrations before icons, the sacrifice of livestock, repetitive terrifying or painful rituals, investment in costly objects and architecture, celibacy, religious violence and non-reciprocal altruism, to name a few…” (Bulbulia, 2004, 655) Costly Signalling Theory, however, as I hope to show in section 4 below, demonstrates how benefits to individual fitness might be secured by just such costly behaviour. It would appear, prima facie, that religious beliefs are at odds with our prevailing every-day human cognitive stance. As Atran says, “From an evolutionary standpoint, the reasons for why religion should not exist are patent: …it is unrelentingly counterfactual and even counterintuitive.” (Atran, 2007, 421) Religious behaviour antedates the evolution of anatomically modern humans That religiosity is deeply rooted in prehistory is attested to by – if contested - ritual burials from the Middle Palaeolithic (Smirnov, 1989, 199) such as those of Neanderthals at Shanidar and, arguably, the earlier inhumations of Homo heidelbergensis at Atapuerca. But as Hawkes cautions us, while it is always hard to draw inferences about behaviour from artefactual data alone, “…unaided inference from the material remains to spiritual life is the hardest of all” (Hawkes, 1954, 161-162) Donald explains succinctly why such inferences are likely to be so contentious: “Symbolic artefacts…rarely encode the conventions governing their use” (Donald, 1998, 184).While climbing the ladder of inference, this is the rung on which we have the least secure purchase. However, these reservations not withstanding, Grűn and Stringer et al have argued cogently that determinations derived from U-series and ESR analyses of bones from Israel has established that the “oldest known symbolic burials are those of early modern humans at Skhul 8 and Qafzeh.” And that “the best age estimate lies between 100 and 135 ka.”(Grün and Stringer et al., 2005, 316) These early dates would suggest that characteristically religious behaviour emerged very recently in the archaeological record while the anatomically modern humans are recorded from ca. 200,000ya. Renfrew presents the difficulty well:” The problem for the neuroscientist and the evolutionary archaeologist is to understand how (religious behaviour) has come about on so short a time frame, (i.e. in the archaeologically recent past) when the human genome has been established for much longer…” (Renfrew, 2008, 2041) Archaeology has, with varying degrees of plausibility, seen chronologically patterned data in the archaeological record. Insoll (Insoll, 2004, 45) summarises well how the evolutionary process has been recruited by the earlier literature to serve an explanatory theory of staged development from the first evidence of religiosity to the contemporary religious landscape of the major world religions. Shamanism was associated with the hunter gatherers of the Pleistocene, and animal imagery was abundant testimony to the importance of these quarry of Cro Magnon European hunters; the Neolithic revolution gave rise to increased levels of reverence for the ancestors who were associated with the land which was now subject to familial ownership and embodied the cycles of seasonality and fertility; the increasing social stratification associated with sedentism saw the emergence of a priestly caste, an increasing emphasis on the value of sacrifice and the centralised control of resources; finally, the world religions are seen as emerging from the development of literacy giving rise to what Whitehouse has termed the “doctrinal mode” of religiosity (Whitehouse, 2004). This neat and tidy chronology correlating food resource strategies and ideology appealed to the evolutionary processualists and the Marxist framework adopted by archaeologists like Childe but this sequencing of stages of religiosity from the Pleistocene through the Holocene was not supported by the archaeology which evidenced a mismatch between putative religious iconography and these resource acquisition strategies; the 9 progression from animism through polytheism to monotheism is likewise not a function of time or social complexity. 10 Acknowledgements Dr. Fiona Coward, Post Doctoral Researcher, Institute of Archaeology, University College London (First Supervisor who kindly made herself available to advise at several stages of drafting this dissertation) Dr. James Steele Director, AHRC Centre for the Evolution of Cultural Diversity, Institute of Archaeology, Reader in Comparative Archaeology, Institute of Archaeology, University College London (Second Supervisor who kindly agreed to discuss the neurological substrate of religiosity) Dr. David Wengrow, Reader in Comparative Archaeology, Institute of Archaeology, University College London (who kindly fielded my questions concerning counterintuitive beliefs) Dr. Andrew Garrard, Senior Lecturer, Institute of Archaeology, University College London, MSc Palaeoanthropology and Palaeolithic Archaeology co-ordinator (who kindly made himself available at all stages of the drafting of this dissertation to answer general questions) Dr. Patrick McNamara, Director, Evolutionary Neurobehavior Laboratory Assistant Professor, Boston University School of Medicine, Department of Neurology (who kindly answered my questions on his research into the role of dopamine inhibition and religiosity) 11 1.0 Neural substrates of religiosity: an overview There would appear to be three biological levels of organisation of the neural substrate underlying any human behaviour: underlying genetic determinants, a range of neurotransmitters and specific neural pathways in the brain which – in this case - bear on theistic cognition. It will become evident that the role of the reward system and its successful inhibition is central; it is this feature which facilitates the expression of costly signals of commitment to prosocial behaviour underpinned by theistic beliefs. 1.1 Genetic predisposition Genetic studies of religious behavioural traits seem to suggest that genes which code for monoamine neurotransmitters are highly heritable. Much work has been done in analysing the role they play in eliciting states associated with religiosity since the Minnesota Study of Identical Twins Reared Apart (Bouchard et al, 1990; Stallings et al 1996; D’Onoforio et al, 1999). Hamer has identified a gene (VMAT2) coding for monoamine neurotransmitters which he claims to be “…associated with the self-transcendence scale of spirituality” (Hammer, 2004, 11) although Zimmer has argued cogently that VMAT2 performs an entirely secular prophylactic function (Zimmer, 2004, 111). The work of Comings et al (Comings et al, 2000) has been used to support the case for another genetic component to “self transcendence”: the DRD4 gene which codes for the dopamine D4 receptor though his sampling methodology has attracted much criticism. The critical role of the dopaminergic system in rewarding religious behaviour which engages the prefrontal cortex is testified to by several researchers (e.g. McNamara, 2002) while comparative studies examining both dopamine and serotonin point to their critical role in innervating areas of the prefrontal cortex (Brodman’s areas 32 and 9) in humans and apes (Raghanti, et al, 2008) It should not be a startling conclusion that genes have been identified which code for neurotransmitters critical to so-called spiritual states of mind associated with religious experience, but this observation alone does not support the adaptationist argument. As Koenig and Bouchard point out, “Demonstrating that it (religiousness) is a heritable trait, does not 12 prove it is an adaptation.” (Koenig and Bouchard, 2006, 32) I will discuss the implications for the adaptationist argument in section 4 below. If it makes sense to speak of genetic determinants of religiosity, then the COMT gene, which codes for the enzyme catechol-O-methyltransferase, might be said to be more critical in the early evolution of religious behaviour in early modern humans. McNamara reports that the critical mutation in the COMT gene which is expressed in the most effective inhibition of dopaminergic activity “appears to be a unique human mutation because it has not been found in apes, suggesting that it may be a factor in the evolution of the human prefrontal cortex and thereby of human consciousness more generally.” (McNamara, 2006). The salience I will later give to costly signalling theory relies heavily on work done by researchers on the COMT gene and will be presented in a more detailed discussion of the role of this dopamine inhibitor in section 4.43 below. 1.2 Neurotransmitters 1.21 Dopamine Dopamine has long been implicated in heightened arousal levels in several brain regions (Van Rossum, 1967). Imbalance in dopamine levels have been correlated with Parkinson’s disease (Butler et al 2004): it has been highly correlated with reduced levels of religiosity when depleted, and highly correlated with schizophrenia when raised. The activation of dopaminergic pathways in religious practices has been demonstrated to reinforce such practices and has been held to contribute to the dependency and elevated mood associated with them. Andrew Newberg’s research confirms the critical role that these neurotransmitters play in religious states (Newberg, 2010). Other studies report the key role of this neurotransmitter in fixing repetitive religious behaviour via the reward system. Schjødt et al (Schjødt et al,2008) report that studies of praying activities of a Danish Christian group reveals the engagement of the ventral striatum, specifically the, 13 nucleus accumbens, in activating the dopaminergic reward system. The significance of this region being engaged lies in the role the striatum is understood to play in fixing reward-related learning. The authors conclude that both the dorsal striatum and the ventral striatum are implicated. McNamara has also drawn attention to the role that the reward system plays in prayer whereby “activation… is both intrinsically rewarding and necessary for (the) acquisition of many of the behaviours that religion seeks to foster…” (McNamara, 2002, 143) However, it is the inhibition of the dopamine receptors at sites in the prefrontal cortex which is of greater significance in making possible the costly signalling of the agent’s predisposition to engage in altruistic actions. Altruistic behaviour was facilitated once the gene coding for COMT had become fixed in the human genome. Religious beliefs would have ensured that cooperative behaviour was selected for and uncooperative behaviour subject to sanction. The Composite fMRI scan in Figure 2 illustrates well, the location of the sites affected. Fig. 2 Composite fMRI scan activation data (in red) shows brain sites affected by COMT gene type during a working memory task (clockwise from upper left: right lateral, left lateral, right medial, left medial). Source: National Institute of Health news release Tuesday, May 29, 2001. (Retrieved 08/05/2011) 14 1.22 Serotonin The role of serotonin in regulating mood has long been understood. It is now clear that serotonin pathways are recruited in religious states, in particular, the personality trait of “selftranscendence”. “A role for the serotonin system in relation to spiritual experiences is supported by observations of drugs such as LSD, psilocybin, N,N-dimethyltryptamine, mescaline, and 3,4methylenedioxymethamphetamine that are known to cause perturbations of the serotonin system in several brain regions.” (Borg et al, 2003, 1967) Figure 3 illustrates the neural pathways involved in both the dopamine and serotonin innervation of cortical areas. Fig.3 The Reward System: Dopamine and Serotonin pathways in the brain (rodensor.com) Note particularly the dopaminergic pathway from the ventral tegmental area to the frontal lobes. 1.23 Endorphins 15 Several researchers have identified the presence of heightened endorphin levels as critical in reinforcing group bonds in a religious context. Robin Dunbar’s work suggests that Karl Marx’s dictum that “religion …is the opium of the people” (Marx, 1844) may not have been purely metaphorical. His central claim is that religious behaviour performs a similar function to grooming activities in primates and is mediated by similar biological mechanisms being activated by opioid receptors. Just as language is effective in social grooming in members whose group size exceeds that in which individual physical one-on-one grooming is practicable, so collective religious behaviours cause the release of endorphins which are critical in developing group bonds; at an individual level, it constitutes an effective reinforcement mechanism in securing successful alliance-building behaviour. He further shows that when this activity is synchronised, the volume of endorphins released is vastly increased. (Cohen, et al, 2010, 106108) vf 1.24 Summary of the role of neurotransmitters The work of Way and Lieberman suggests that there is, “a robust cross-national correlation between the relative frequency of variants in (central neurotransmitter systems, particularly the serotonin (5-HTTLPR, MAOA-uVNTR) and opioid (OPRM1 A118G) genes and the relative degree of individualism–collectivism” (Way and Lieberman, 2010, 203) in contemporary culturally and geographically diffuse population groups. This provides an explanatory framework for the integrated functionality of both genetic determinants and group-level expression of those determinants of religiosity; in doing so they combine both Dunbar’s insight into the role of endorphins in collective religious behaviour and a plausible explanation of the genetic mechanism which explains the distribution of such behaviour. The crucial role of the reward system and – as will become clear below- its inhibition, in overcoming the free rider problem will be central to this dissertation. I would expect the role of oxytocin to be a critical factor in securing the trust which underlies delayed reciprocal altruism but there have been few studies which have examined the role of this neurotransmitter in such behaviours, exceptions being that of De Dreu et al, 2010, and Israel et al, 2008 who point to the possible role of oxytocin 16 and vasopressin in social behaviour associated with music and altruism. Raised levels of oxytocin were also shown to be implicated in altruistically generous “anonymous one-shot interactions” in a study by Zac et al. (Zac, et al, 200). 1.3 Cytoarchitectural features: processing of executive functions and intensionality in frontal lobes The proportion of the total cranial capacity devoted to executive function in the frontal lobes would presumably have needed to cross some threshold before actions could be planned with a view to securing prosocial goals. Furthermore, such goals would also have to have been executed in such a way as to ensure the acknowledgement and approbation of other group members to have been effective. Fourth level intensionality, necessary to incorporate the collective attribution of god’s understanding of the motives and predicted responses of those affected by actions issuing from such altruistic intent, could likewise only be entertained once a threshold prefrontal volume had been crossed. The capacity to generate effective, costly, religiously sanctioned signalling, presupposes the prior development of a suite of behaviours which are executed by pathways in the prefrontal lobes; these would include, evaluating alternative actions, predicting likely effects of the alternative actions (specifically evaluating their impact on the inclusive fitness of the agent) and face recognition (to identify past co-operators and defectors). Critically, regions of the right dorsolateral and orbito-frontal cortices enervated by the dopaminergic reward pathways were shown to be recruited during “intentional and private religious practices” in findings reported by Beauregard & Paquette (Beauregard & Paquette, 2006) and Azari (Azari et al, 2001); its effective inhibition at sites in this region by the enzyme catechol-O-methyltransferase facilitates the expression of costly signals which leave a characteristic signature in the archaeological record. This process will be clarified in section 4.4 below. Other regions of the brain especially the temporal lobes have been associated with heightened self-reported levels of religiosity and the parietal lobes have been the focus of d’Aquili and Newberg’s research into so-called “self transcendence” states associated with meditative experience. But as these features of religiosity are only tangentially associated with 17 theistic belief, I will not be discussing them here. For similar reasons I shall not dwell on Persinger’s research on the role of the temporal lobes in religious experience. In summary, the significance of the genetic component of characteristically religious cognition and behaviours has been demonstrated and the critical role of the role of the reward system, at sites in the prefrontal cortex and the mechanism for its inhibition has been identified. 18 2.0 The contribution of developmental psychology to an understanding of religiosity A belief in the survival of the mind/soul after death is widespread and is a central feature of all formalised religions with a literary tradition. That such beliefs were already prevalent in prehistory is an inference drawn from several excavated mortuary sites the evidence for which I present in section 5.2 below. Here I present data from developmental psychology which strongly suggests that, “…belief in supernatural agents pirates the brains mental inference systems that are designed to reason about everyday intentional (living) agents.” (Bering, 2006a, 128) 2.1 Common sense dualism in infants primes the brain for supernatural concepts Several factors have worked to predispose the human mind to believe in life after death and so populate the Palaeolithic theological landscape with ancestors: our inability to conceive (with or without accompanying mental imagery) of our own death is what he calls the “simulation constraint” hypothesis - the relevance of such constraints for ancestor worship is clear; “offline social reasoning” which makes possible a belief in the continuing existence of others even though they are neither visible nor communicatively accessible. This results in the predilection to believe that the afterlife is somehow a place. (Bering, 2006 b, 456) That such beliefs were primed by the human mind/brain imply that, “developmental mechanisms underlie intuitive accounts of dead agents’ minds.”(Bering and Bjorklund, 2004, 217) and it is not the result of acculturation. 2.2 Children’s belief in gods is a default cognitive stance Barrett’s “preparedness hypothesis” suggests that there is “a certain conceptual bias that makes acquiring God concepts particularly natural” among children, though conceding that the extent to which such capacities were realised was a function of other socio/cultural variables. (Barrett, 2003, 310) Furthermore, this incipient theism of preschoolers was not found to be a product of extrapolation from prior ascriptions of god-like characteristics to adults. The work of Olivera 19 Petrovitch concluded that even children not exposed to the cultural priming of a belief in a creator God nonetheless believe in the existence of such a God, “which suggest that there are cognitive universals in a number of domains of human knowledge” (Petrovitch, 2007, 360). 2.3 Over developed sense of cause and effect in infants primes brains for god concept Deborah Kelemen and her colleagues found that very young toddlers have a predilection to describe artifacts in terms of their possession of teleofunctional capacities. (Kelemen, 2007) This cognitive bias persists in pre-schoolers who are as predisposed to offer a functional explanation for artifacts as for natural phenomena. (Kelemen and Di Yanni, 2005a, Kelemen and Di Yanni, 2005b) This bias may persist into adulthood and is confirmed by studies of Altzheimers patients whose causal, explanatory framework is compromised by the onset of the disease. Lombroso et al pointedly add, “…it is unknown whether a widespread preference for teleology is ever truly outgrown”. (Lombroso, et al, 2007, 999) 2.4 The sense of lacking control, increases illusory pattern perception in adults What may prove to be one of the most powerful explanatory theories for explaining the frequency and prevalence of superstitious belief comes from what - at first blush - would seem the most unlikely of fields: behavioural economics. Whitson and Galinski (Whitson and Galinski, 2008) strongly suggests a statistically significant positive correlation between the tendency to see illusory patterns in randomly generated data and the debilitating intensity of an individual’s sense of lack of control. The evolutionary value of pattern recognition is self evident and a number of studies have reported the significance of this variable in determining levels of both physical and psychological well-being (Klein, et al, 1976; Cohen, et al, 1991). Galinsky and Whitson’s work suggests that this strategy is effective in reducing anxiety, which would be of evolutionary benefit in enhancing the efficacy of subsequent efforts to reassert the individual’s control over the environment, counteracting the enervating effect following a sense of individual impotence. More specifically, “belief in a controlling religious deity may serve as an especially attractive resource for restoring a sense of externally controlled order when personal control 20 cannot protect one from the anxiety of a random and uncertain world.” (Kay, et al, 2010, 44) Michael Shermer draws similar conclusions from his review of the work of Vyse and Heltzer (Vyse, 1997) but states it more boldly :”Those patterns have to be given an identity, and for thousands of years many of those identities were called gods” (Shermer, 2000, 62) In summary, research in developmental psychology has demonstrated that a belief in supernatural agents is facilitated by the default cognitive stance that gods exist. This and the fact that we are primed to believe that the ontological status of persons is independent of their being perceived, prepares us for theism and the plausibility of ancestor worship. 21 3.0 The contribution of Evolutionary Psychology to an understanding of religiosity 3.1 Intuitive Psychology (Theory of Mind) The ability to attribute mental states and processes to others on the basis of inferences drawn from facial and bodily cues is a precondition for effective social interaction. The socially debilitating effects of such a deficit are well documented in autism and schizophrenia patients. The significance of the frontal lobes in processing information about the intentions of others and their states of mind subsequent to interaction with us (in addition to the predicting of the likely responses of others in response to our future actions) is critical for social engagement with others, all of which is conducive to maximising inclusive fitness. The importance of recognizing the hierarchical position of other group members and exhibiting behaviour designed to forge alliances to secure resources led to the early characterisation of this suit of behaviours as issuing from what was termed the Machiavellian mind. If Mithen is correct that attempting to ascertain and assuage or ingratiate oneself with gods is a misapplication of social intelligence then interacting with gods who are seen as able to punish or reward behaviours identified as defecting or cooperating behaviours respectively is explicable. 3.2 Intensionality Subsequent studies have confirmed that the volume of the orbital pre frontal cortex has a linear relationship with levels of intensionality (Powell, et al, 2010) More specifically, while for most purpose three levels of intensionality are habitually used in modern human interactions, religious cognition imposes a fourth embedded layer to accommodate the “mind of god” as described in section 1.3 above: “For a supernatural-based religion to have any force in making us toe the social line, I have to believe that you suppose that there are supernatural beings who can be made to understand that you and I desire that things should happen in a particular way.” (Dunbar, 2003, 177) As figure 4 below shows, Dunbar has demonstrated persuasively that the threshold for fourth level intensionality was crossed only with the advent of Homo sapiens. 22 Fig.4 Robin Dunbar’s Thresholds of intensionality in the Homo lineage (Dunbar 2003) 3.3 HADD (Hyperactive Agent Detection Device) Stewart Guthrie was the first researcher to suggest that the human mind was biased to overattribute agency, “where there was some uncertainty about the provenance of an unexpected phenomenon.” (Guthrie, 1980) but as in most things, David Hume had presaged much of the 20th century psychology of religiosity in the 18th. In 1757 he wrote about what Boyer has since called the “hypertrophy of social intelligence”: “We find human faces in the moon, armies in the clouds; and by natural propensity, if not corrected by experience and reflection, ascribe malice and good-will to everything that hurts or pleases us.”(Hume, 1757) The seemingly innate human propensity to infer the presence of agents which might impact on the inclusive fitness of the individual has come to be called the Hyper Active Agent Detection Device (HADD). The agency may constitute a threat, for, example, a predator, or an opportunity, for example, the timely arrival of a caregiver or a potential mate for a reproductively fertile individual. The evolutionary advantage to being hypersensitive to such agency is obvious: a false positive will likely be in some measure distressing but a false negative may be fatal. In the absence of evidence that the agent is either human or in some other way natural, the inference is habitually drawn that it is 23 supernatural. The human predisposition to seek patterns in randomly arranged natural phenomena is discussed in section 2.5 above and must work to compound the activation of the HADD. 3.4 Minimally Counterintuitive Concepts Some attempts to explain the “stickiness” of supernatural concepts has involved the notion of the Meme (Blackmore, 1999; Dawkins, 1976; Dennett, 2006) but as I don’t feel a case can be made that the principle of evolution through natural selection is substrate neutral, I shall not devote any space to a consideration of the concept in this context. A more cogent explanation supported by experimental empirical data is that put forward by Pascal Boyer (Boyer, 2001) based on the modular view of human cognition closely associated with the work of Jerry Fodor (Fodor, 1983). Boyer demonstrated that counter intuitive concepts which violated expectations and which were embedded in narratives which were for the most part intuitive were more readily retained than those which did not. Dr. Wengrow (Wengrow, 2011) has criticised Boyer for underestimating the significance of institutional and technological factors in explaining the epidemiology of counterintuitive concepts which he sees as explaining the traction that monstrous iconography gained after the Neolithic revolution but not before it. While his paper provides a valuable reminder that explanations for the epidemiology of such concepts is incomplete without looking at the social and technological causal factors thrown up by the Neolithic revolution, I think he misrepresents the notion of minimally counterintuitive beliefs as developed by Boyer which I believe does illuminate the cognitive fluidity of anatomically modern humans in entertaining counterintuitive concepts that violate the expectations set up by the cognitive domains elucidated by Mithen. Figure 4 below illustrates the kind of cognitive fluidity embodied in the blending of animal and human forms, (and possibly also displaying violation of expectations concerning folk psychology and natural history in the manipulating of the material used). Therianthropic imagery from Chauvet and Trois Freres (and discussed in the context of shamanism in section 5.3) likewise display this blending of traits from animal and human forms and is included as Figures 14 and 15 below. That this is what Boyer had in mind, when he speaks of counterintuitive 24 concepts, however, is doubtful; such counterintuitive images alluded to by Wengrow are not embedded in any intuitive context as demanded by Boyer (but in the absence of a literate tradition how would we know?). Dr Wengrow (Institute of Archaeology, UCL) kindly agreed to meet to discuss his paper last term, and I took the opportunity of outlining my objections to his criticisms of Boyer and these are appended as appendix 1. Fig. 5 The Löwenmensch figurine from Stadel im Hohlenstein In summary, research in evolutionary psychology has allowed me to isolate several mechanisms including inherited folk psychology, our understanding of intensionality, our hyperactive propensity to detect agency and the memorability of minimally counterintuitive concepts further facilitate our predisposition to believe in supernatural agency. 25 4.0 Adaptation or exaptation? Researchers, who have argued for an adaptationist explanation of religiosity, have focused on a number of mechanisms which facilitate inclusive fitness. Bulbulia, discussing the claims for such evolutionary adaptedness, summarises the array of explanations given in the literature: “enhanced solidarity and co-ordination among the faithful (the cohesive function), an answer book to life’s riddles, an existential purpose generator, a means for providing hope and solace to the suffering, an adaptation for inter-group warfare, or for morality, and various combinations thereof…” (Bulbulia, 2004, 656) My aim is to discuss those theories which are salient in the literature and focus on costly signalling theory as the most consistent with the working of “natural selection to solve problems that our ancestors faced during our species’ evolutionary history.” (Cosmides and Tooby, 1997, 5) I shall attempt to argue that there is no plausible “neurotheological” explanation for a widespread belief in gods but that the evidence strongly supports the exaptationist position. That the neural substrate of religiosity has been selected for in response to distinctly non-religious pressures is cited in support of the exaptationist argument. 4.1 Existential purpose generator Questions which ask about the meaning of life which have been answered by some form of religious response have been as enduring a feature of human history as the brevity of the religions which have so self-confidently offered spurious solutions. The surface grammar of some interrogative utterances lends them a prima facie credibility whereas they do not, in fact, function as genuine questions. They are examples rather, of what Gilbert Ryle calls a category mistake (Ryle, 1949). Because it makes sense to ask, “What is the meaning of this sign?” and “What is the purpose of this process/component/feature etc.?” it seems plausible – if mistaken to ask questions where the predicate includes the word “ life”. The answer to such questions has often been contingent on the putative existence of supernatural deities of one kind or another. (Park, 2005) While it may not be a rational question, it is still a frequently asked one and one to which a belief in God or gods can provide a psychologically - if not logically – satisfying answer. Any answer which is predicated on the 26 prior assumption that a god exists, is only as satisfying as the logical force of that epistemologically prior demonstration of the god’s existence. Such a demonstration is entirely lacking. 4.2 Causal explanation for phenomena This approach has been most closely associated with the work of Lewis Wolpert (Wolpert, 2006). He argues that the selection pressure that drove encephalization was the need to understand phenomena in terms of causes and effects. He cites the characteristically human propensity for tool-making as evidence for this contention. Belief in gods, he argues, was a misapplication of this drive to attribute causality to the, frequently dangerous and seemingly inexplicable, ancestral environment of early humans. This is not a plausible explanation: it is ,as Atran would say, “mind-blind” ignoring the role of natural selection in priming the human mind for theistic beliefs; it is at odds with the observations that heightened religiosity is often a feature of cultures which enjoy high levels of scientific sophistication such as the USA where far from declining in the face of scientific progress, religious beliefs are increasingly widespread. 4.3 Religious belief is an adaptation to facilitate good physical and psychological health Several studies have reported a positive correlation between religiosity and physical and psychological health, and in view of its salience in Bulbulia’s CST approach which is highly plausible, I will deal with the salubrious argument at length here. The strength of this correlation has lead a number of medical practitioners to urge the incorporation of the patient’s religious/spiritual beliefs into clinical interventions (Cadge, et al, 2009) That active religious observance and symptomatic pathology are inversely related, is likewise attested to in the literature (Hannay, 2006) 4.31 Increased longevity Several studies have reported a high correlation between increased longevity and religious observance (Schnall, et al,, 2010; Matthews et al, 1998, 123; Hummer et al, 1999) 27 4.32 Attenuated experience of stress It has long been understood that religious beliefs can play a powerful role in ameliorating stress. Pargament detailed the mechanisms by which this religiously based coping strategy is effected (Pargament, 2000). Research by Inzlicht et al, suggests that the anterior cingulate cortex, normally activated in the experience of anxiety, is much attenuated in electroencephalographic neural reactivity recorded in subjects undertaking the Stroop task leading him to conclude that, “religious conviction provides a framework for understanding and acting within ones environment, thereby acting as a buffer against anxiety and minimizing the experience of error.”(Inzlicht, 2009, 385) These findings are supported by other studies (Ano and Vasconcelles, 2004, 477; Maselko, et al, 2008, 1009) 4.33 Lower incidence of chronic physical ill health A causal relationship between religious observance and physical health has long been accepted. Typical in this regard is Levin and Schiller’s wide ranging review of the literature which revealed a high correlation between physical wellbeing and religiosity: in particular, “cardiovascular disease, hypertension and stroke, colitis and enteritis, general health status, general mortality, cancer of the uterine corpus and cervix, all other non-uterine cancers, morbidity and mortality in the clergy, and cancer in India.” (Levin and Schiller, 1987, 9) In a review of the literature bearing on the correlation of religious belief (measured by religious attendance) and health outcomes, (Levin and Vanderpool, 1987) it was concluded that 81% of studies had revealed this correlation. Furthermore, this correlation held across a wide range of pathologies from cardiovascular disease to cancer. These authors found in a later study that religious observance was also positively correlated with lower blood pressure (Levin and Vanderpool, 1989). A more cautious note is sounded by research conducted by Powell, et al, who concluded that while the “evidence fails to support a link between depth of religiousness and physical health”, they did suggest that “church/service attendance protects healthy people against death” (Powell, et al, 2003, 36) Several studies report a positive correlation between patient religiosity and the recovery process. (Oxman, et al, 1995) 28 4.34 Religious belief encourages risk-averse behaviour Not only is risk averse behaviour correlated with religious observance but studies have also shown that even among those whose practices can compromise their health status, religiosity can result in a diminution of the associated risks (Matthews, et al, 1998; Bahr and Hoffman, 1998; Button et al, 2010, 1619) 4.35 Assortative sociality Research by Fincher and Thornhill suggests that the number of religions practiced in any given climatic zone is highly correlated with the frequency of pathogens present in the area leading them to conclude that, “religion manifests from evolved behavioural strategies for the avoidance and management of infectious disease.” (Fincher and Thornhill, 2008, 2587) More precisely, they concluded that religious diversity decreased the further you travelled from the tropics. Supporting evidence is provided by Murray et al, who claim to have found a negative correlation between the tendency of cultures to enforce conformity and the prevalence of infectious pathogens present in that area over which that culture prevails. (Murray, et al, 2011) 4.36 Conclusion While the data does suggest that religiosity is positively correlated with desirable health outcomes in some groups, it is often weak and can be explained by the therapeutic affect that such social support networks provide rather than the efficacy of intersessionary prayer. The prophylactic role of religious belief can frequently be explained by recourse to the placebo effect. 4.4 Religious belief as a solution to the free rider problem Religion has long been seen as exercising what Talcott Parsons called a tension management function (Parsons, Bales and Shils, 1953); it is an effective mechanism for resolving intra-group conflict. A more nuanced approach has been taken by more recent research which suggests that theistic belief plays a critical role in providing a solution to the free rider problem. 29 4.41 Early prosociality secured by kin-based altruism a function of Darwinian fitness An otherwise solitary individual gains many advantages by being a member of a group including greater access to and defence of resources (critically including both energetic resources and a larger pool of potential mates) enhanced security and sources of support (e.g. non-kin “grandmothering” reducing infant mortality, and vigilance conducive to enhanced collective security). Group living makes many demands of an individual attendant upon the greatly increased need for cooperative behaviour. Cooperative behaviour involving some form of altruism is more intuitively comprehensible when such sacrifices are made on behalf of offspring or other individuals to whom the benefactor is closely related. “Selection can favor altruism toward close relatives because recent common descent provides a cue of genetic similarity” (Boyd, 2006, 1555) However there has been some criticism of the principle that the propensity for altruistic behaviour is not unambiguously a function of genetic relatedness e.g. West et al, 2001. 4.42 Prosociality secured by non- kin-based altruism as a function of inclusive fitness Within groups of increasing size, however, members are more distantly related to each other yet the demand for cooperation does not lessen but tends to increase. Herein lays the problem, as Atran makes clear, “Commitment falls off precipitously as genetic difference increases between individuals.” (Atran, 2002, 119) Hamilton’s Rule (Hamilton, 1964) explains how this commitment to perform altruistic acts in the service of cooperation where a more distant kinship relationship subsists between the benefactor and the recipient. Even if an action impairs an individual’s “Darwinian fitness”, it may be conducive to that individual’s “inclusive fitness” where the beneficiaries are more distantly related as illustrated in the transaction between an “agent” and”recipient” in fig 5 below. 30 Fig.6 The operation of Hamilton’s Rule: : r = the genetic relatedness of the agent to the recipient, B = the reproductive advantage conferred on the recipient and C = the reproductive cost to the agent of the act “A key feature of inclusive fitness is that, as defined, it describes the components of reproductive success which an actor can influence, and therefore which they could be appearing to maximise.” (West, et al, 2010, 3) Providing costly signals which serve to demonstrate a predisposition to cooperate rather than to defect in social transactions is an example of just such a maximisation strategy. 4.43 Prosociality in early hunter-gatherers: non-kin based reciprocal altruism and the role of COMT However, the degree of genetic relatedness of the members of a group is likely to be inversely proportional to the size of the group: increasing group size entails that the members are increasingly likely to be only distantly related to most other members of the group while the demands for the expression of altruistic actions increases and with it the urgency to detect freeriders. Given that group living confers a reproductive advantages of its members, how is prosociality to be secured between members of the group who were not conjoined by sanguinity? Peter Singer has written persuasively of the innateness of the tendency to expand the circle of the recipients of our altruism beyond immediate kin. (Singer, 1981) The mechanism by which reciprocal altruism 31 operates has been well understood since Robert Trivers’ ground-breaking article in 1971.(Trivers, 1971) These costs which are incurred in order to evince a preparedness to behave cooperatively, can range from the relatively minor inconveniences of spending time and effort in practices which have no direct benefit to the practitioner; personal disfigurement; various levels of personal discomfort including high personal economic costs; behaviour which compromises the physical wellbeing or reproductive fitness of the individual, and even life threatening and self-sacrificing practices which can confer a benefit on surviving genetically related members of the group. The reward system which functions to reinforce behaviour which is generally pleasurable must, however, be inhibited in order to effectively defer the gratification which costly signalling demands. I draw heavily on the work of Patrick McNamara and colleagues at Boston who have demonstrated the critical role than the enzyme catechol-O-methyltransferase has in securing the inhibition of the dopamine pathway that enervates the frontal lobes. (McNamara, 2006) Costly signalling is designed to secure a commitment to perform altruistic practices. These are defined as, “…behaviours that involve strategic costs to the individual displaying them, costs that extend beyond the baseline costs that all behavioural actions entail, and are therefore hard to fake by individuals not able to bear the relevant cost.” (McNamara, 2006, 193) The temptation to defect rather than cooperate is, in part, a function of the perceived levels of relatedness among members of the population but it becomes increasingly important to be able to detect free riders as the genetic relatedness of group members decreases. Detecting defection becomes increasingly important when there are those who will embrace the dictum attributed to Marx that, “Sincerity is the key to success; if you can fake that you’ve got it made”. (Reilly 1998, 137) (Groucho’s letters are a rich source of insight in so many fields.) The preparedness to display costly signals demonstrates the commitment of that individual to the group, resolves the free-rider problem which is so inimical to successful group cooperative behaviour be that a hunting expedition, the coordination necessitated by inter-group hostility, or the trustworthiness of an agent in honouring the promise made in the course of interpersonal social transactions. 32 My contention is that the sequence of events is as follows. The reward system, particularly the dopaminergic pathways which are normally activated when impulsive behaviour is satisfied, must be inhibited by the individual if he or she is to demonstrate their ability to , as McNamara says, “,inhibit over a long period of time the kinds of free-rider behavioural strategies that would exclude them from participation in the cooperative group.” (McNamara 2006, 194) This inhibitory effect is achieved by the action of catechol-O-methyltransferase which methylates released dopamine as part of its metabolism to homovanilic acid. This activity takes place in the frontal lobes. It is the public demonstration of costly signals underpinned by religiosity which enables the individual to claim membership of the group of co-operators to which he or she wishes to belong. What the proximal causes of such inhibition are, I am still uncertain, but that appropriate practices are taught by the host community is implied by his remarks that the candidate group member gains admittance by “learning to inhibit current appetitive or consumatory responses”. (McNamara 2006, 197, my emphasis) This raises the intriguing, but at this stage highly speculative, possibility that the walls of Chauvet are testimony to such tutelage. 4.44 Costly signalling as a commitment to practices ensuring group health outcomes Bulbulia (Bulbulia, 2006) has made a cogent argument for a version of the CST that sees the primary function of costly signalling as expressing a hard-to-fake, commitment to signals of health which is underpinned by socially-shared religious/supernatural beliefs. A discussion of Bulbulia would take me too far from my concern to establish the case for CST qua CST, but I flag this approach up as a significantly nuanced version of the CST hypothesis which is also potentially testable but alas, not by a significant set of archaeological data. I will only remark that these findings are lent plausibility in the light of the established correlation with health adumbrated in section 4.3 above and my observations on the shaman’s role as healer in section 5.3 below. 33 4.45 Levels of inter-group competition a function of intra-group altruistic behaviour Groups of early modern humans inhabiting the Environment of Ancestral Adaptedness would have been differentially predisposed genetically to practice non-kin based altruistic behaviour and those populations in which the appropriate alleles were selected would be more successful in attaining group goals than those groups in which they weren’t. Something like this was hinted at by Darwin in “The Descent of Man”, who maintained that a tribe whose members, “…were always ready to give aid to each other and to sacrifice themselves for the common good, would be victorious over most other tribes…” (Darwin, 1981, 166) Supporting data is provided by researchers, such as Samuel Bowles, who have claimed to identify data in the ethnographic record which is applicable to the demographic structure of modern humans at the critical period in our prehistory. (Bowles, 2006) 4.46 Costly signalling behaviour most effective in a religious context The moral argument for a belief in the existence of God has always been one of the most popular and that moral actions are those approved of by the gods has always exercised a potent fascination. This is in spite of the fact that it was discredited by Plato writing in 5th century BC Athens. He demonstrated that – in spite of a widely held belief to the contrary - attempting to draw inferences about morally relevant behaviour from the belief that a God exists such that he rewards certain behaviours and proscribes others, is logical fallacious. Plato’s eponymous hero of the dialogue Euthyphro, asserts that right conduct is defined thus: “…what all the gods love is pious and holy, and the opposite which they all hate is impious”. (Plato, 1970, 46, 9e) Plato’s Socrates (Euthyphro’s interlocutor) asks for the grounds for this assertion: “…I should wish to understand …whether the pious or holy is beloved by the gods because it is holy, or holy because it is loved by the gods.” (Plato, 1970, 46, 10a) The point here being that if the answer is the former then the statement that “x is right because god determines that it is”, becomes a semantically vacuous apriori truth (god commands as good, those things which god commands as good); if the latter, then it is an aposteriori truth but the inconvenient corollary is that presumably god is using some criterion other than his dictat to determine the rightness or 34 wrongness of any action. In spite of the logical fallacy of the moral argument, which would not have been available to the hunter gatherers of the Palaeolithic, that cooperative behaviour was favoured by the gods would have gained considerable traction. There appear to be many reasons why moral values are usually underpinned by religious sanctions; clearly this is likely to have been most effective if the notion of “fictive kin” could be recruited. Genetic relatedness is an effective constraint on the temptation to defect in social transactions and just such a notion is a characteristic of religiosity. The kinds of cooperative actions demanded by the group to which an individual belongs because are more likely to be forthcoming if there is a widespread belief in a kinship relation subsisting between biologically unrelated individuals. Empirical data exists which supports the contention that prosociality and its concomitant behaviour of costly punishment of defectors is more effective when religiously primed. McKay concludes that: “religions harness the by-products of genetically inherited cognitive mechanisms in ways that enhance the survival prospects of their adherents” (McKay et al, 2011, 1858) Shariff and Norenzayan have published studies that support these conclusions identifying “religion as a facilitator of the emergence of early large-scale societies of cooperators.” (Shariff and Norenzayan, 803, 2007) Dunbar has demonstrated the effectiveness of collective religious behaviour in overcoming the free rider problem by triggering the release of endorphins and thus reinforcing group bonding behaviour. Empirical support for this function of religious behaviour first identified by Rappaport (Rappaport, 1999) is provided by a study into nineteenth century communes in America which concluded that “… communes that imposed costlier requirements survived longer than less demanding communes and whether costly requirements and religiosity interact to promote cooperation” (Sosis and Bressler, 2003, 212); theistic priming in religious ritual was found to be highly correlated with levels of cooperation in Israeli Kibbutzim by Sosis and Ruffle. (Sosis and Ruffle, 2003) Chris Stringer has suggested that intensified levels of intragroup conflict for resources and mates was a function of increased density of larger human groups and that the friction engendered by such competition could “have driven…changes favouring cooperative and even sacrificial behaviours within the conflicting groups” and that 35 “the development of religion may have provided an important means” of maintaining equilibrium. (Stringer, 2011, 221) 4.47 Types of costly signal in the archaeological record Richard Sosis (Sosis, 2006) sees costly signals as being one of three kinds: rituals, badges and bans. Only the first is likely to leave an enduring signature in the archaeological record. This tripartite division is helpful in setting the parameters of my dissertation. While bans are critical to modern world religions as proxies for commitment, they are unlikely to leave a trace in the archaeological record of the earliest expressions of religiosity. For example, while it is possible to argue that dietary bans, might leave an anomalous profile in the faunal remains of a Palaeolithic group that practiced such a ban, it is very unlikely that proscribed behaviour of other kinds could be inferred from the material record of early modern humans. I am unaware of any attempts to draw such inferences from either fossil or artefactual remains. Likewise, badges are unlikely to constitute a significant archaeological presence. However, body decoration using ochre and shell beads may be evidence of such ritual and has been reported from several sites from the Middle Stone Age in Africa (e.g. MacKay and Welz, 2008; Jacobs and Roberts, 2009; see further references in section 5.0 below) but while they clearly embody some symbolic function we cannot know if this was religious or carried a very secular meaning. (eg the therapeutic purposes of tattoos found on ӧtzi the so called “Iceman”) I shall therefore focus my research on the first type, for the purposes of this paper. Rituals typically involve objects and votive offerings are perhaps the archetypical vestiges of costly rituals. That they were materially costly is indubitable, the energetic and emotional costs and the costs they exacted in human suffering and increased mortality is harder to divine. Atran makes explicit the causal connection between the religiously inspired sacrificial act and the social role it has in discouraging defection and act in the interests of the group: “Religious offerings always involve non-recuperable costs” (material and human)…Why? Because human representations of agency and intention include representations of false belief and deception, human society is 36 forever under threat of moral defection. Simple consent among individuals seldom, if ever, successfully sustains cooperation among large numbers of people over long periods of time. Emotionally hard to fake and materially costly displays of devotion to supernatural agents signals sincere willingness to cooperate with the community of believers.” (Atran, 2002, 114) Offerings may be rendered “non-recuperable” by being thrown into deep water, broken or burnt or measured in the time spent on an activity which has among its opportunity costs the compromising of reproductive fitness. It is in just this state that such objects as diverse as Gravettian and Neolithic statuettes, the images on the walls at Lascaux and Chauvet, to the Iron Age Battersea shield; wherever objects have been put beyond use either by design or by destruction. The chart in Fig. 20 below correlates the necessary and sufficient conditions for effective costly signalling and illustrates the range of artefactual and fossil data it leaves in the archaeological record. Limited inter-group migration would have ensured that the genetic composition of such groups would have been biologically homogeneous and facilitated the widespread acceptance of “fictive kin” applicable to those group members less closely related. As such, it could be expected to constitute a mechanism which secured the “objective” of inclusive fitness and therefore Hamilton’s Rule would still apply. CST provides a powerful theoretical framework which integrates the neurology with the archaeological and anthropological data. If this approach is correct, it satisfies all the objectives delineated in the opening paragraph of my abstract and will support my conclusion that much artefactual evidence is best explained as arising from the engagement in culturally determined costly signalling, much of it guaranteed by the watchful eye of ”fictive gods”. Sharif and Norenzayan endorse the conclusions of Bowles alluded to in section 4.45 above, that cooperative behaviour would have been facilitated by a widespread belief in gods and that these groups would most likely have been more successful than those which lacked such a widespread belief: “ancestral societies with culturally widespread God concepts would have outcompeted societies without such concepts, given the cooperative advantage of believing groups” (Shariff and Norenzayan, 808, 2007) Stringer reaches much the same conclusion: 37 ”computer simulations have shown that in many situations of conflict between hunter-gatherer groups, beliefs that encourage self-sacrifice, sometimes including death on behalf of the group, can actually flourish culturally and genetically.” (Stringer, 2011, 223) 4.48 Conclusion The emergence of costly signalling theory (CST) presents a powerful and coherent explanatory framework which integrates data bearing on the neurochemistry of the reward system, and its inhibition, (McNamara, 2006) with behaviours which facilitate the diffusion of prosociality in a population (Axelrod, 1984) and in particular, religious behaviours. It also holds out the prospect of integrating the neurological data with the archaeological data by identifying the signature detectable in the ritual material culture of the late Pleistocene and early Holocene by such ritual behaviours I discuss below in section 5. In summary, the grounds for believing theistic religious belief an adaptation are strongest when the critical importance of evolutionary pressures for resolving the free rider problem in early human groups is fully understood. This is not group selection: the benefits bestowed by cooperative behaviour confer a fitness advantage on the individual which possesses the appropriate alleles relative to the selfish behaviour of members of early human groups who lacked them. I am still inclined to believe that the evidence points to the theistic religious cognition under-pinning such Palaeolithic prosociality is an exaptation. 38 5.0 Archaeological evidence for the presence of religious belief I first propose to allude briefly to artefacts from the Late Stone Age in Africa which have been held to embody symbolic ritual religious behaviour. I shall then attempt an evaluation of the European Upper Palaeolithic data which includes parietal cave imagery and mortuary practices, Gravettian and Solutrian anthropomorphic mobiliary sculpture and briefly mention costly signalling practices from the Late Holocene. Since McBrearty and Brooks’ seminal paper in 2000, (McBrearty and Brooks, 2000), the origins of human symbolic behaviour have been firmly located in sub-Saharan Africa in the Middle Stone Age. There is much artefactual evidence from sites such as Blombos Cave and Still Bay in southern Africa which suggests that symbolic behaviour as exemplified by the use of red ochre and shell beads was already present in the MSA. Iron rich hematite blades have been reported from Pinnacle Point in South Africa and have been dated to 165,00ya (Minkel, 2008) while inscribed red ochre has been retrieved from Blombos cave and dated to 75,000ya (Henshilwood, et al, 2002). Perforated N. kraussianus hell beads have been recovered from Blombos dating from 75,000ya,, Nassarius gibbosulus shellbeads dating from 82,00ya have been reported from Grotte des Pigeons in Morocco (Bouzouggara, et al, 2007) and also from Es-Skhul (Israel) and Oued Djebbana (Algeria) dated to 100,000 to 135,000 years ago. (Vanhaeren, et al, 2006), ostrich shell beads from the Loiyangalani River valley, Tanzania (Knight, 2004) Textiles too could be expected to have been a prominent feature of shamanic ritualistic practices if the ethnographic record is a guide. That such beliefs were instrumental in producing the artefacts that have been recovered is, however, at least highly likely in that they were produced by anatomically (and a fortiori neurologically) modern humans although there is little evidence of unambiguously symbolic behaviour which could be said to issue from a belief in supernatural entities. 5.1 Evidence of early ritualistic behaviour ca. 70,000 years ago When so much so-called modern human behaviour can be found in Africa in the MSA, we might be forgiven for feeling optimistic that we will find some evidence of religious ritualistic behaviour here too. Unfortunately the evidence is very tenuous. The so-called “python stone” 39 from Tsodilo in Botswana, received a great deal of attention when the report was published (figures 6 and 7). Dating from 70,000 years ago, the authors claim that the stone represents a python snake which they hold to be central to a ritual practised by the ancestors of the San who currently inhabit the area. I don’t find their claim plausible for several reasons. The strata from which they claim the tools used to indent the “skin” of the python were recovered, is not securely dated; it is not at all obvious that the stone resembles a snake and were it not for the fact that the python is accorded a significant place in present-day San religious belief, it is difficult to see how such an interpretation could have been made. Although the stone spearheads found at the site (fig 8) exhibit a colour consistent with their having been selected for ritualistic purposes and might well have originated from some distance away, we can never know what ritualistic purpose they served. There is no indication of the system used for dating the spearheads themselves. This is critical to the interpretation of the site as they are used to establish the date of the “python stone” itself and to attribute religious ritual significance to it. Fig. 7 The so-called “python stone” from Tsodilo, Botswana. ca.70,000 ya www.afrol.com/articles/23093 (Science Daily, retrieved Nov. 30, 2006) Fig.8 The “Python Stone” illuminated at night. 40 Fig.9 Spearheads excavated from Tsodilo, knapped from stone transported from hundreds of kilometres distant from the site The Cave at Chauvet is well known for the parietal imagery found there but the so-called “bear skull altar” is claimed to be diagnostic of human symbolic ritualistic behaviour. That the stone on which the skull rested functioned as an “altar” seems an unwarranted assumption as well as starkly anachronistic. Zach Zorich, writing more soberly in Archaeology, notes that many Cave bear remains were found at Chauvet, but confesses,” Why this one was placed at the edge of a large stone block remains a mystery.” (Zorich, 2011) Fig. 10 The so-called “bear skull altar” at Chauvet (www.bradshawfoundation.com/clottes) 41 Fig. 11 Jean Clottes at the so-called “bear skull altar” at Chauvet 5.2 Mortuary practices That ancestors continue, in some sense, to exists in order to monitor and “police” the social behaviour of their ancestors is prevalent in modern day human populations and was widespread in populations of behaviourally modern humans in the past. Evidence for belief in some form of life after death has been sought from human inhumations but this association is, however, often highly contestable. Interment with or without grave goods of some kind does not constitute either a necessary or sufficient condition of belief in an afterlife (Ucko, 1969). However, it does seem to me that such goods do lend support to the prevalence of such a belief but Ucko’s is correct to caution against over reliance on mortuary practices alone to support any contention that a belief in life after death was widespread in the absence of any corroborating evidence. I can find no references in the literature to unambiguously deliberate burials before the Middle Palaeolithic. 5.21 Middle Palaeolithic & Neanderthal burials There is abundant evidence of the practice of deliberate interment among Neanderthal populations. Evidence for elaborate preparation of the corpse and grave goods is less compelling. Stringer, however, is prepared to concede that such burials are imbued with ritual symbolic significance: “…special rocks or tools seem to have been placed with the bodies, as tributes or perhaps even in anticipation of an after life.” (Stringer, 2011, 89) I am inclined to agree with Pettitt who concludes that while “no convincing example of grave goods is known from Neanderthals…it seems that at least in some Neanderthal groups the dead body was explored and treated in socially meaningful ways.” (Pettit, 2002, 18) Neanderthal mortuary practices seem rather to constitute caching. 42 Fig.12 Neanderthal burial at Kebara, (Kebara 2) ca. 60, 000 BP piclib.nhm.ac.uk/piclib/www/image.php?img=85721 5.22 Burials of anatomically modern humans Burials in anatomically modern humans are less ambiguously ritual interments both in the manner of the burial and the presence of grave goods. Although Skhul 5 burial is cited by Stringer as the earliest symbolic burial dating from ca. 115,000 years ago, I agree with Mellars who argues that the Qafzeh burial (ca. 92,000 years ago) presents the earliest evidence of: “unmistakably intentional grave offerings”; (the presence of) “a number of deliberately perforated seashell ornaments, together with large quantities of used and apparently heattreated fragments of red ochre” (Mellars, 2006, 9384). Such interments is suggestive of what many developmental psychologists have called “common sense dualism” and identified as “an inevitable by product of self-consciousness” (Bering, 2008, 34) Also some belief in an after life can reasonably be inferred from the treatment of the corpse which in a foetal or recumbent position (as at Kebara and Qafzeh respectively) clearly mimics postures adopted in sleeping. 43 Fig.13 Burial of an anatomically modern human (skeleton at the Qafzeh Cave (Israel), accompanied by a large deer antler and dated to ca. 90,000 – 100,000 B.P. (Mellars, 2006) An intriguing application of Dunbar’s levels of intensionality to mortuary practices is made by Stringer. He draws heavily on Pettit’s analysis here. Just as thresholds of increasing levels of intensionality are inferred from the enlargement of the prefrontal cortex throughout the Pleistocene, so methods of interment are suggestive of the increasing sophistication of these levels. Figure 13 below indicates the way in which Stringer sees these mortuary practices as corollaries of the threshold levels of intensionality. Species Chimpanzee Attitude to dead Interest in body of deceased Deposition of corpse Intensionality “I believe that you are dead” Mortuary practice No attempt to move or bury corpse Caching of corpse “I know that you must be deposited in a specific place” Reverential “Because of your role, you Full burial in special treatment of corpse must be disposed of in this location way , by this method, at this accompanied by place, as recognized by our ritual and symbolic social rules” objects Fig 14 mortuary practices correlated with threshold levels of intensionality (adapted from Stringer, 2011, 212) (Heidelbergensis?) Neanderthal Anatomically Modern Human 5.3 Shamanism in Franco-Cantabrian parietal imagery in the Upper Palaeolithic Explanations of the religious significance of the Franco-Cantabrian parietal imagery in the Upper Palaeolithic have customarily stressed the central role of the shaman as an intermediary between 44 the world of the living and that of the dead. Therianthropic imagery, for example, is usually interpreted as depicting shamanic practices which are held to be identical to those of their contemporary hunter gatherer counterparts which are well documented in the ethnographic record. However, there are several reasons why any attempt to explain the iconographical significance of European Palaeolithic cave imagery as analogous to the San or !Kung is flawed. I give fuller reasons for my scepticism below as appendix 2. It is more plausible that they exercised a role on a par with Jain ascetics identified by David Sloane Wilson as enforcing prosociality and the shaman’s intervention would clearly satisfiy the role of an agency imposing a plausible pattern and causal explanation on otherwise random events that compromise human well-being consistent with Whitson and Galinsky’s research alluded to above in section 2.5 above. Fig.15 The “Sorcerer” from Chauvet (c. 30,000 BP) www.bradshawfoundation.com/clottes 45 Fig. 16 Depiction of Shaman: Trois Freres, France, (upper Palaeolithic) ca. 13,000 BC faculty.gvsu.edu/websterm/Myth/Sorcerer.html For all of the reasons adumbrated in Appendix 2, I am very sceptical of any explanation of Franco-Cantabrian cave imagery which is exhausted by reference only to its depiction of shamanistic practices. My concern in this section has been to offer a plausible answer to the question, “Why were the caves painted at all?” not to attempt an evaluation of the competing explanation of their meanings for the Upper Palaeolithic people who painted them. The evidence is consistent with numerous hypotheses. It is beyond my brief to assess the merits of the theories that have included the following: expressing hunting magic (Henri Breuil, 1952); embodying dyadic and gender symbolism (Leroi-Gourhan, 1968 and Laming-Emperaire, 1962); calendrical symbolism (Marshack, 1991); shamanism (Lewis Williams and Jean Clottes, 1998). Likewise, the nonrepresentational shapes which have traditionally been attributed to shamanic entopic experiences have recently been reinterpreted by von Petzinger who has made a detailed study of the distribution and frequency of such non-figurative geometric images in Palaeolithic cave sites and concluded that they could plausibly have functioned as a proto-pictographic system of symbolic signs (Ravilous, 2010) This finding is at odds with the long-standing theory that such imagery issued from entopic experience. I am sceptical of von Perzinger’s explanation as it seems self-evident to me that it is a precondition of communicative signs that, that which was symbolically represented by them, should be in the public domain and interpersonal not refer to 46 private and personal experience which could not be coded for in this way. I am prepared to concede, however, that there is some likelihood of some of these images depicting some kind of ritualistic behaviour. The less ambitious explanation for the production of cave imagery which limits itself to the practice of costly signals of commitment by the individual to the group outlined above in section 4.47 at least has the merit of not recruiting assumptions that are unsupported by the data. The creation of images would have been time consuming both in the acquisition of raw materials and in their use in creating the images; the opportunity costs sustained would likely have been great as no hunting or other resources would have been acquired while the men (and the assumption made is that they were usually men) were engaged in painting; the danger and discomfort in reaching the locations deep within the caves would have been very stressful. 5.4 Parietal imagery exhibiting costly anatomical signalling It is not for nothing that CST is sometimes known as the “handicap approach” (Bliege Bird and Alden-Smith, 2004) It is relevant to my argument that there are frequent depictions of mutilated hands at Chauvet and at other Upper Palaeolithic cave sites such as Grotte de Gargas in France. There are several strands of supporting evidence: the high proportion of hand depictions which show evidence of mutilation (of 231 hands, 144 were missing digits at Gargas); the digits were more likely to have been amputated than lost through disease such as frostbite (which is in any event, rarely recorded amongst Eskimo who inhabit a similar climate to that inhabited by the Upper Palaeolithic people whose hands are depicted at sites such as Chauvet and Gargas) and the fact that thumbs are amputated probably has more to do with the its critical importance in manipulating, repairing and servicing artefacts involved in resource acquisition rather than their being enervated with a richer blood supply to ward off frostbite ; finger bones which were clearly amputated have been found at Gargas embedded in clay (Barrière, 1976); some researchers have suggested the loss of digits was the result of diseases such as Raynaud's disease, Ainhum's disease, or even leprosy(e.g. Sahly, 1969) but it is not explained why such high incidents of these pathologies should afflict the painters, or if it did, why this should have 47 privileged them in being able to give their handicap such publicity. Fig 16 below shows the frequency of hand images featuring amputated fingers at the site of Grotte de Gargas. The Bradshaw Foundation web site raises the possibility that what appear to be amputated fingers may be the result of bending back the finger in order to convey some kind of code. But there is no attempt to explain what this behaviour may have been code for or what grounds there are for believing this to be true. The site does go on to concede that “it is also possible that the amputation of fingers at the joints was part of shamanic initiation rituals at certain times during the Upper Palaeolithic.” Finger mutilation, practiced by the Khoekhoe of southern Africa is cited as supporting ethnographic evidence. The matter could be conceivably settled by the positioning of the “fracture”. If the amputations occur invariably at the point of flexion at the first knuckle, then the amputation argument is much weakened. The case for mutilation enjoyed growing support (e.g. Hooper, 1980) and Janssens cites Weinert and Casteret (Casteret, 1951) who marshalled a large amount of ethnographic evidence to support their contention “that these are cases of sacrificial offerings.” (Jansenns, 1957) Fig. 17 Frequency of hand prints featuring amputated fingers at Grotte de Gargas (Leroi- Gourhan, 1967) 48 5.5 Anthropomorphous figurines Many recent researchers have bemoaned the fact that theories about the cultural significance of the sculpted anthropomorphous figurines of the Gravettian and Solutrian periods, and the more recent Magdellanian, in Europe have been of a one-size-fits-all type (Hansen, 2001). Too little attention has been paid to regional variation and the significance of the diversity of materials used. The ludicrous sobriquet of “Venus” is patently inappropriate in view of the low proportion of figurines which exhibit the typically voluptuously proportioned feminine form of their classical namesake. That they all performed the same function is likewise an assumption we are not entitled to make. 5.51 Middle Palaeolithic evidence I cannot see in the The Berekhat Ram Figurine, (Fig.17) the glimmerings of anthropomorphic sculpture as Goren-Inbar and Peltz would have us believe (Goren-Inbar and Peltz, 1995). I am convinced of the arguments of Noble and Davidson (Noble and Davidson, 1996) that the resemblance a female form is a result of taphonomic rather than anthropogenic factors. D’Errico and Nowell quote Noble and Davidson dismissively – but I think mistakenly – that the “Berekhat Ram object was formed by human agency rather than by natural forces . . . hence we cannot say that it was a thing made to resemble anything’ (Noble and Davidson, 1996, 75 quoted in D’Errico and Nowell, 2000, 127). Fig. 18 The Berekhat Ram Figurine: a false dawn? D’Errico, F. and A. Nowell, 2000, 124 49 5.52 Gravettian and Solutrian anthropomorphous figurines A great deal of Palaeolithic (and Neolithic) figurative sculpture is either an unambiguous depiction of aspects of the female sexuality or such a depiction can reasonably be inferred from the artefact. Marija Gimbutas has developed an influential theory of the original purpose of such artefacts: that they are expressions of a mother goddess religion which issued from the fertility. This is not a hypothesis which enjoys as much consensus as it once did and for several reasons: the diversity of the figurines stylistically and in materials used does not suggest a single explanation of this kind. Bailey’s dismissal of Gimbutas (Bailey, 2005, 16) as lacking any support for her twin contentions that women exercised a dominant position in what she claimed was a matriarchal society seems to me to be fatal to the mother goddess theory. Unambiguously anthropomorphic figurines do not emerge until the Gravettian in Europe ca. 28,000-22,000ya. It is not central to my dissertation question to evaluate the competing theories which have sought to explain the cultural meaning of these figurines beyond establishing that their fabrication entailed an enormous expenditure of time and effort and that the key to their meaning must lie in the costliness of their production. That this costliness can be calibrated with reference to group commitment underpinned by religiosity is a hypothesis that has not yet been adequately tested. It seems incontestable that there is a preponderance of female rather than male depictions and there is prominence accorded to secondary sexual characteristics in many of them. This supports the view that they embody fertility symbolism of some kind. There is enormous diversity, both in the materials used, the sexual characteristics foregrounded and the stylistic variety displayed in the depiction of hair, body posture and fabric “worn”. It is highly likely that the craftsmen were motivated by as wide a range of intentions in crafting them. 50 Gravettian and Solutrean “Venus figurines” So-called “Venus of Willendorf” ca. 24,00022,000ya (Germany) Venus of Brassempouy Ca. 25,000 ya (France) Dolni Věstonice figurine ca. 28,000 31,000ya (Czech Republic) Fig.19 Gravettian and Solutrian anthropomorphous figurines and find sites 5.53 Magdalenian anthropomorphous figurines Figurines continued to be produced throughout the Magdalenian and while the range of materials from which they were made increases, it seems entirely plausible that while they seem to display a more stylised form, they were produced for similar reasons to the previous Gravettian and Solutrian anthropomorphous figurines. 51 Late Magdalenian Carved Venus figurines and Late Magdalenian sites Female figurines of stone, bone and ivory from various Late Magdalenian sites. Antiquity Publications, Ltd, published in Fiedorczuk et al. 2007 The Lady of Monruz. Jet pendant representing a feminine figure, Monruz (Neuchâtel). Magdalenian, ca. 13000 B.C Fig. 20 Late Magdalenian carved anthropomorphous figurines and site locations 5.54 Neolithic figurines Figurine production persisted into the Neolithic and it has been argued that the fertility of domesticated crops and animals gave added impetus to production. Although fertility was not a characteristic preoccupation of the Neolithic: it was a difference in degree rather than of kind from the hunter gatherer groups which preceded it relying as they did on a regular harvest and fecund livestock. Most plausibly, some researchers have argued that the patterns of distribution and exchange throughout both this and the preceding periods reflects their use as mediating inter-group rivalries and alliances. (Chapman, 2001) Population growth and dispersals at this period would likely have resulted in greater contact between hitherto isolated groups and increased the urgency of, and scope for, such costly signals. 5.6 Summary of the archaeological evidence 52 In short, I have tried to argue that the data supports an explanation for a great deal of the artefactual record for which costly signalling theory is central; this does not commit me to any of the competing theories which have sought to explain the cultural meaning of the FrancoCantabrian cave imagery or the Upper Palaeolithic from Europe where the main weight of the interpretation of such artefacts falls upon ethnographic evidence. I am sceptical of such a program. An account of the significance of Franco-Cantabrian cave imagery which is consistent with its production as a type of costly signal is possible however. The representational imagery is still viewed through the prism of the earliest theories of so-called “cave art” even though the artfor-arts-sake hypothesis was discredited nearly a century ago. The images were not produced to satisfy the aesthetic of contemporary viewers but the motivation for their production is best sought in the act of production itself. The manner of their production is inconsistent with the intention to present images which are to be viewed in the same way as Michelangelo’s Sistine Chapel ceiling in Rome or Giotto’s frescoes in the Scrovegni Chapel in Padua: the images display little respect for perspective or proportion, sequences of images are rarely connected by any coherent narrative form and the surface of the walls of the caves themselves are palimpsests of superimposed images which is hardly consistent with the imputed intention to depict an individual subject or a scene composed by a collection of subjects (e.g. the “herd” of horses at Chauvet). These characteristics are however what you would expect if the act of production was more important that the product. The philosopher J.L. Austin coined the phrase “performative utterance” to differentiate speech acts which did something in the speaking of them from those speech acts which could be said to describe something by speaking (Austin, 1962, 5-6) He gives as examples of the former linguistic formulae appropriate to naming (e.g. at baptism or launch ceremonies), bequeathing and proposing a bet (in the contexts of the writing of a will and speculating on an uncertain outcome respectively). In an analogous way, costly signalling theory predicts that the meaning of parietal imagery, anthropomorphic three dimensional objects and all the many other costly artefacts is the very act of production itself: it is a “performative depiction”. Their value lay in the act of enduring the danger and discomfort of entering the depths of the caves, investing time and effort in creating the images and incurring the 53 opportunity costs to inclusive fitness which such acts entailed for those whose hands held the charcoal and ochre. The table contained in Fig 20 below is designed to illustrate the range of signals and the preconditions for their effective expression. Criteria Mortality Cost must be non- Death followed by recuperable interment and or ritual cannibalism Types of costly signal Mutilation or Aquatic disfigurement deposition of votive offerings Irremediable Hydrologically mutilation or secure deposition disfigurement Sacrifice must be Impracticability of hard to fake feigning death Fabrication/destruction of ritual object Ritualistic compromising of the integrity of the object and/or time consuming fabrication of object (e.g. elaborately decorated/ carved/sculpted or by breaking/ smashing/ defacing etc., once produced) Integrity of object not restorable by repair Disability incurred subject to daily communal visibility Sacrifice must be Total loss of Anatomical feature of high value reproductive fitness critical for human reproductive fitness Deep riverine or lacustrine locations Precious metal/mineral or otherwise high status objects Sacrifice must be Publically observable Proof of disability publically context of death incurred (e.g. missing observable finger from painted hand on cave wall) Location must admit of mass observation (aquatic refection supports belief that gods are also observers) Votive offerings Late Palaeolithic and deposited in water Neolithic (especially iron age weaponry) High opportunity costs incurred by expending high levels of time/effort/ concentration on production of sacrificial object Fabrication process must be subject to high public visibility of fossil remains, of Parietal “art” human burials and evidence of ritual non-nutritional cannibalism (Iron Age bog-bodies?) Fig. 21 Necessary and sufficient conditions for effective costly signalling practice and examples signature in the archaeological record Examples Fossil/ artefactual evidence of their The currencies in which the costs of signalling are borne are many. They include the parietal imagery discussed above, the amorphous figurines of the Upper Palaeolithic, the shattered Neolithic figurines, the votive deposits of high status goods and weaponry in the Iron Age and, one can surmise, a great deal of perishable food resources leaving no trace in the archaeological record. It is also relevant here to ask if the so-called Iron Age bog bodies from northern Europe 54 are best explained as the ultimate costly signal. Although beyond my brief, complex societies record the practice of valuable offerings to the gods in exchange for their beneficence and preservation of the moral order 6.0 Fossil evidence 55 Fossil evidence which has a bearing on the growth of religiosity is of four kinds Firstly, the tempo of the increase in cranial capacity since Australopithecus afarensis which is demonstrated by fig. 21; secondly, cranial remains demonstrate the trend of an increase in size of the neo cortex in the hominin clade (fig.22); thirdly, Fig. 23 illustrates the trajectory of the encephalization quotient calculated from hominid fossils differentiated by taxon (and including Pan) As it is a ratio of cranial capacity to body size, it is likely to be diagnostically more significant of cognitive capacity than simple cranial capacity expressed in cubic centimetres/millilitres; and fourthly, thresholds representing step changes in the levels of intensionality reached over the same time frame as revealed by Robin Dunbar are included as Fig.4. I will discuss these data sets in this sequence. Many of the metrics used here to illustrate the increasing cognitive capacity of hominids have been criticised on the grounds that endocasts have been more unreliable than assumed. Wiliam Kimbel summarises these problems well in his review of a recent work on brain evolution in hominoids: “Plagued by poor preservation tiny sample sizes, and the conspiracy of the meninges (the protective tissues sandwiched between the brain and the inner surface of the brain case) to veil cerebral surface detail, endocasts are the Rorschach tests of palaeoanthropology…” (Kimbel, 2005, 387) However, the graphs are included here to demonstrate the general trajectory only. 6.1 Increased cranial capacity The timing and tempo of the upward trajectory of cranial capacity calculated in cubic centimetres is clearly indicated in Fig. 20 below. Aiello and Wheeler’s “expensive tissue hypothesis” explains the mechanism that drove the two dramatic increases in brain expansion, the first “around two million years ago from Australopithecines to H. erectus” and the “second was a little more than half a million years ago, when Homo erectus became H. heidelbergensis” (Wrangham, 2010, 113-114). Some researchers have cast doubt on attempts to date punctuational events in the evolution of the brain (Schoeneman, 2006) but the evidence that step changes occurred, at approximately these times and involving these species does seem persuasive. That religiosity presupposes a minimum cranial capacity seems clear, what that threshold is, is not. 56 Nor is it clear that H. heidelbergensis had no concept of the supernatural bearing in mind that some specimens had a cranial capacity well within the range of modern humans. When the fossil data is evaluated in conjunction with the artefactual record, it would seem reasonable to suppose that a minimum cranial capacity is a necessary condition for thinking religiously if not sufficient. Most researchers agree that the threshold for relevant cognitive processing could only have been reached with the advent of anatomically modern humans ca. 200,000 years ago. Fig.22 Evolutionary trajectory of gross cranial capacity in the human phylogeny (adapted from Aiello and Wheeler, 1995) 6.2 Increased size of neocortex The pre-frontal, parietal and temporal lobes have all been implicated in religious thinking and their expansion is well illustrated in Fig. 21 below. Religious (and non-religious) cognition has been shown to engage the ventromedial prefrontal cortex (Harris, 2009); the temporal lobes have for long been associated with hyper religiosity and several authors (e.g. Persinger, 1987) have claimed to have identified this region of the brain as primarily implicated in heightened religious states. These claims have been contested on the grounds that the frontal lobes have also to be recruited in order to generate and manipulate concepts of agency both of the devotee and of the divine (Atran, 2002, 186). I have alluded to the central role of the prefrontal cortex in religiously sanctioned costly signalling behaviour in section 1.3 above. 57 Fig. 23 Evolved human cranial morphology illustrating the expansion and growth of the neocortex. Note particularly the trajectory of the expanding frontal lobes. (A) Pan troglodytes, chimpanzee, modern (B) Australopithecus africanus, STS 5, 2.6 My (C) Australopithecus africanus, STS 71, 2.5 My (D) Homo habilis, KNM-ER 1813, 1.9 My (E) Homo habilis, OH24, 1.8 My (F) Homo rudolfensis, KNM-ER 1470, 1.8 My (G) Homo erectus, Dmanisi cranium D2700, 1.75 My (H) Homo ergaster (early H. erectus), KNM-ER 3733, 1.75 My (I) Homo heidelbergensis, "Rhodesia man," 300,000 - 125,000 y (J) Homo sapiens neanderthalensis, La Ferrassie 1, 70,000 y (K) Homo sapiens neanderthalensis, La Chappelle-aux-Saints, 60,000 y (L) Homo sapiens neanderthalensis, Le Moustier, 45,000 y (M) Homo sapiens sapiens, Cro-Magnon I, 30,000 y (N) Homo sapiens sapiens, modern 6.3 Increase in brain mass to body mass ratio Figure 24 below illustrates the trajectory and the tempo of step changes in the human clade. An increase of from 0.5% to nearly 2.5% is recorded from Australopithicines to anatomically modern humans. What is particularly striking is that, as Kappelman puts it “The most dramatic changes occur with the appearance of modern Homo sapiens at about 100,000 years ago and include a decrease in body mass and an increase in relative brain size that appears to have been driven by selection for smaller body mass.” (Kappelman, 1996, 243) 58 Figure 24 Brain mass as a percentage of body mass 6.4 Increase in the levels of intensionality Robin Dunbar’s comparative studies of primates and his extrapolation results to early hominins (Fig.4) indicate that the ability to entertain levels of intensionality is largely a function of the size of the frontal lobes. As I indicated in section 3.2 above, fourth level intensionality which includes that of God in relation to the intentions of agents vis-à-vis the other members of the group seems only to have been achieved by anatomically modern humans. In summary, the fossil record reveals the trajectories of brain size, neocortex volume and levels of intensionality explain why theistic religious cognition leaves no signature in the 59 archaeological record until thresholds were crossed with the advent of anatomically modern humans. 60 7.0 Discussion 7.1 Distributed nature of religious cognition Neural correlates of religiosity exist but, as I hope I have shown, are neural networks evolved to accomplish very secular objectives but recruited to serve some very religious causes. Darwin himself seemed to suggest that the emotional states and cognitive processes associated with religiosity were highly distributed: ”The feeling of religious devotion is a highly complex one, consisting of love, complete submission to an exalted and mysterious superior, a strong sense of dependence, fear, reverence, gratitude, hope for the future, and perhaps other elements” (Darwin, 1981, 68) That the neural substrate recruited in religiosity is distributed and not localised is what would be predicted if it is the case that religiosity is an exaptation, not an adaptation. Structures co-opted for processes involving a religious content were originally evolved for other purposes. As Barrett and Lanman aptly put it, “Religion springs naturally from the way ordinary human cognitive systems interact with ordinary human social and natural environments.”(Barrett and Lanman, 2008, 110) Boyer outlines these cognitive systems which, “…deal with detection and representation of animacy and agency, social exchange, moral intuitions, precaution against natural hazards and understanding of misfortune.” (Boyer, 2003, 119) McNamara is himself inclined to the view that the health benefits of religious belief do indeed justify what he calls a biocultural adaptation. (Harris and McNamara, 2008) Echoing Bowles, the authors also concluded that “…religiousness promoted healing in some individuals in ancestral populations… (and)… promoted outgroup antagonisms in ancestral populations” (Harris and McNamara, 2008, 79) I have tried to show that the human mind/brain was not adapted to gain access to the, speciously, numinous. I remain persuaded of the case. 7.2 Costly signalling does not support an adaptationist argument Costly signalling theory provides a sound theoretical bridge which links the neurological with the archaeological: whether it is the time consuming sculpting of European Upper Palaeolithic 61 figurines; the enduring of the opportunity costs attendant on the dangerous practice of producing parietal cave imagery; the materially costly votive offerings of the Early Holocene and the often extravagant sacrifices of early complex societies. It offers an insight into the motivation behind the energetically expensive behaviour of the Shamanic acolyte, as well as all manner of behaviours which have not left a trace in the archaeological record while leaving a vivid impression on the mind of the ethnographer of San !Kung and Australian aborigines. While I am reluctant to see any credibility in the adaptationist case, I am not so certain that some version of co-evolution might not turn out to be the case. 7. 3 costly signalling underlies the pervasiveness of Palaeolithic religiosity The distinction casually drawn between the sacred and the profane by twentieth century anthropologists and archaeologists seems increasingly spurious when applied to the preliterate religious traditions of hunter gatherers of the Palaeolithic. All the sub-systems beloved of the New Archaeology seem to have been imbued with religious beliefs; to say that the concept of religiosity bleeds profusely into the other sub systems of subsistence, kinship, social structure and so on, is an understatement – religiosity was positively haemophiliac. Whitehead’s distinction between imagistic and doctrinal modes of religiosity recruiting different memory networks might map onto the religiosity of preliterate and complex societies respectively. (Whitehouse, 2004; Whitehouse and McCauley, 2005) 7.4 Distributed nature of religious cognition undermines the neurotheological programme The distributed nature of neurological features recruited for religious purposes has implications for theist apologetics: a so-called God module does not exist therefore His existence cannot be inferred from such a module. 7.5 Weaknesses in argument I am painfully aware that this chain of argumentation has more than its fair share of weak links. Some might argue that my operational definition of religiosity is bordering on parody as it 62 ignores the powerful effect it exercises on the expression of an individual’s ethical values, sense of selfhood and relationship to the world into which were are – without consultation – thrown; my depiction of the causal connection between the role of dopamine inhibitors and altruistic behaviour will probably turn out to be too simplistic :many neurotransmitters are likely to be recruited in religious behaviour. In a personal communication, McNamara, who is in the forefront of research into neurotransmitters and religiosity, has encouraged me in my belief that though the reward system is critical: “production of costly signals likely depends on many neurotransmitters but I agree that dopamine is central” (Pers. com.) Prosociality is not underpinned solely by religious beliefs, practices and priests (kinship and nonreligious mechanisms which serve to reinforce “fictive kin” are significant but have no place in this paper). I have tried to provide what seems the most plausible explanation for much of the evidence in the archaeological record of ritual behaviour that can plausibly be attributed to religiously underpinned, costly signalling practices. Finally, it must be acknowledged that it is often impossible to distinguish between religiously sanctioned costly signals and non-religious ones in the archaeological record. I have tried to be methodologically conservative in making such attributions. 7.6 Limitations of analysis It should be acknowledged that I have not sought a definitive answer to the question, “What are the causes of religiosity and what unambiguously situates it palaeoarchaeologically.” There is no single point in time or place that religiosity could reasonably be said to emerge. Durkheim, writing at the turn of the twentieth century was prescient in this matter (as in so much else) when he wrote that, “like any human institution, religion begins nowhere” (Durkheim, [1912], 2001, 9). However, perhaps he was a little too pessimistic. He hadn’t had the benefit of reading McBrearty and Brooks’ groundbreaking paper on symbolic behaviour in Middle Stone Age Africa (McBrearty and Brooks, 2000). As a sub-set of symbolic behaviours, we should not be surprised to find archaeological evidence for religious practices in sub-Saharan Africa contemporaneous with the Howeisons Poort and Still Bay industries. However, the dating of 63 such artefactual remains is a good deal more secure than attempts to explain the behavioural context in which such remains were imbedded. I have attempted an explanation of why costly signalling practices were conducted but I have not speculated on what the culturally determined meanings of these practices and associated artefacts were. The following scenario seems broadly correct and is suggested by the foregoing analysis. The increasing frequency of theistically underpinned costly signalling in helping to solve the free rider problem seems to be correlated with demographic variables of population density and post-African dispersal patterns which seems to have been contemporaneous with the explosion of symbolism in the Upper Palaeolithic: Hamilton’s Rule would have been less effective in securing reciprocal altruism when early human group sizes had exceeded thresholds at which the members of early human groups could rely upon their genetic relatedness to ensure co-operative behaviour and the disinclination to defect in social transactions. It is a hypothesis which has yet to be tested but which is entirely consistent with all data bearing on the emergence of religiosity in the archaeological record and current estimates of population distribution patterns in the Palaeolithic. I believe my attempt to provide some kind of chronology is plausible and merits further research and it is in the light of this aspiration that I hope this paper will be judged. Finally, I am all too aware that-in spite of the erroneous claims of the post-processualists that twenty-first century eyes can see the cultural attributions of the Palaeolithic minds that lie behind them: our understanding of much of the meaning of the symbolism of the past is for ever beyond our grasp. It is often those who study the distant past who are most reluctant to accept the irretrievability of much of it. Our ability to generate analogies to explain the past too often places a burden on prehistory that it cannot bear. 7.7 Tentative conclusion would be strengthened by further research: 7.71 To what extent is the incidence of costly signalling a function of the intensity of climatic and social stress in the Palaeolithic? If I am right, that religiously sanctioned costly signals are a mechanism for addressing the free rider problem, then the theory would predict that the incidence of costly signals would be 64 positively correlated with periods of social stress when pressures to demonstrate costly signals of group commitment would be greater. I am not aware of any studies which have sought to establish such a correlation in the context of prehistory. The theory predicts a causal relationship between the incidence of the kind of artefacts discussed in section 4.47 above and what would be predicted if the producers were motivated by an intention to demonstrate a propensity to reciprocal altruism through costly signalling as a result of higher levels of stress arising from environmental, inter or intra-group conflict. Bowles’ work clearly points to inter-group conflict as one determinant of such gestures of preparedness to produce altruistic behaviour. The fabrication of costly time-consuming “artistic” endeavours and the practice of ritual disfigurement etc. are likely to be highly correlated with such periods and anecdotal evidence supports this linkage in prehistory as it does also for the incidence of such practices in complex societies. What may turn out to be of greater significance is the demographic impact of sedentism in the Neolithic if Verhoeven is correct (Verhoeven, 2004) 7.72 How highly correlated is the preparedness/demand to produce costly signals of cooperation with other indices of the perceived reproductive fitness of the producers? Is there a demographic profile unique to costly signallers? The age and gender profiles of the fabricators of religiously sanctioned costly signals may be instructive; preparedness to signal cooperation is likely to be highly correlated with other indices of perceived reproductive fitness. Disentangling the artefactual evidence for costly signalling as a non-indexical sign of group commitment from other functions of such signals such as signals of reproductive fitness, demonstrations of status of individuals in groups structured hierarchically, proofs of provenance and demonstrations of entitlement to group membership rights is likely to be difficult, but would seem to be required if religiously inspired signals of altruism are to be unambiguously identified in the archaeological record from background cultural “noise”. 65 7.73 What determined the effectiveness of such costly signals (used as indices of predictably cooperative behaviour) by individuals to other group members in the Palaeolithic? I should like to see research which examines the effectiveness of the range of costly signals which were used as indices of predictably cooperative behaviour by individuals to other group members. Cox et al have demonstrated that the ability to recall the identity of those who have and who have not cooperated in previous encounters is crucial in determining who and who not to engage with, in such social transactions in the future: “In an isolated prisoner’s dilemma game, unconditional defection is an evolutionarily stable strategy (Axelrod 1984). In the repeated game, when strategies (strategists- sic?) are able to use past experience of previous rounds, cooperation sometimes prevails.” (Cox et al, 1999, 370) Costly material signals broadcast with considerable visual publicity or made in a manner calculated to render the agent readily identifiable as a result of conspicuous body modification are two effective means of ensuring that, as Cox et al suggest, “…cooperation can be sustained in large groups when there are sufficient interactions on each round and players are able to base future play on their observations of other players’ past actions.” (My emphasis) (Cox et al 369, 1999) There is considerable supporting ethnographic evidence that such body modification functions in this way (E.g. the lower lip plate-insertions of Mursi women in Ethiopia and the practice of circumcision etc.,) Expensive expenditure of time or effort in a highly visible public domain constitutes much of the evidence for costly signalling frequently underwritten by the activities of supernatural agents whose omniscience permits them a more accurate assessment of the supplicants sincerity than the human witnesses possess. 7.74 Were fragmented figurines apportioned to neighbouring settlements in the locality of the site of fragmentation as a costly signal of commitment to inter-group not simply intragroup cooperation? There is evidence that fragmented figurines in the Neolithic were deliberately apportioned to neighbouring settlements in the locality of the site of fragmentation. (Chapman, 2001) This 66 raises the intriguing possibility that their distribution was a costly signal of commitment to intergroup not simply intragroup cooperation. 7.75 Are religiously sanctioned costly signals a function of the demographic changes in the composition and size of human groups after the dispersal from Africa? An answer to the question “Why does evidence for theistic religiosity only emerge in the archaeological record so long after the human genome had become fixed?” may be answered by the demographic profile of early human groups. Only once Homo sapiens had left Africa, would group size increase to the point where mechanisms needed to obviate the temptation to defect in social transactions by members who were either very distantly related or entirely unrelated to the majority of the group become necessary. Is Dunbar’s number pertinent here? It seems plausible that costly altruistic signalling underpinned by the sanction of theism coevolved with the associated social grooming function of language. Religious beliefs which centred on gods or omnipotent ancestors, and for which the human brain is primed, were likely to have been readily adopted by groups in excess of ca. 150 members. (Dunbar, 1993) These research questions and issues of methodology are listed below in figure 26. 67 Research Questions 2 3 Religiously sanctioned CS Religiously Propensity of and periods of social stress sanctioned CS early human correlated with groups to indices of inclusive thrive as a fitness function of religiously sanctioned CS 1 Research Topic Research Question Were periods of social stress (when pressures to demonstrate costly signals of group commitment would be greater) highly correlated with the incidence of religiously sanctioned CS in the Palaeolithic? How significant a causal factor was the demographic impact of sedentism with the onset of the Neolithic? How highly correlated was the preparedness to signal cooperation through religiously sanctioned CS, with other indices of fitness? How effective were such costly signals as indices of predictably cooperative behaviour by individuals to other group members in the Palaeolithic? Research Methodology Identify proxies for Palaeoclimatic and Palaeodemographic causes of social stress (e.g. dietary indicators such as dental enamel hypoplasia/ostiopathology/s keletal isotopic profile). Map spatio-temporal distribution of proxies onto spatiotemporal distribution patterns of religiously sanctioned CS data. 1. Can Palaeoclimatic and Palaeodemographic factors be unambiguously differentiated? Is it necessary to differentiate them? 2. Is it necessary to distinguish proximal/ distal and endogenous/ exogenous causes? 2. Are there proxies for palaeodemographic contributory factors to social stress? Determine age and gender profile of makers of religiously sanctioned CS in Upper Palaeolithic Identify population centres which produced religiously sanctioned CS and those which did not. Did they prosper differentially? Anticipated Problems 1. Does the Identifying archaeological unambiguous record permit such proxies in the attributions? material record of 2. How much “prospering” reliance would I groups would have to place on be ethnographic problematic analogy? I would be reluctant to do this. 4 Incidence of figurine fragmentation in Upper Palaeolithic correlated with intergroup cooperation Were fragmented figurines apportioned to neighbouring settlements as a costly signal of commitment to inter-group not simply intra-group cooperation? Identify distribution patterns of fragmented figurines and correlate with other indices of cooperation between neighbouring groups 5 Religiously sanctioned CS to solve free rider problem as a function of the African dispersal Is religiously sanctioned CS, to solve free rider problem, a product of the African diaspora post dispersal? Measure the incidence and distribution patterns of religiously sanctioned CS in Palaeolithic 1. Does data exist to enable the identification of such distribution patterns? How can the probable degree of genetic relatedness of early human groups 2. What “other before and indices of after the cooperation African between dispersal be neighbouring established? groups” in the Palaeolithic are available? Fig. 25 Summary of research questions arising from this dissertation. 68 8.0 Summary and Conclusion It is not my intention to argue in this dissertation that all artefactual evidence of ritual religious behaviour is the product of costly signalling, nor, even more modestly, that most of it is. I am suggesting that more of it might be than has hitherto been conceded. There is some dissonance between what I had hoped to achieve when I started this dissertation and what the constraints of theory and evidence permitted me to do: it was my intention to argue that the identification of the inhibition of the reward system in the pre frontal lobes would be sufficient to explain its critical role in the expression of altruistic behaviour; having established the status of altruistic behaviour as a proxy of a public observable commitment to intra-group prosociality. I have tried to explain how this behaviour might have been selected for in that costly signalling likely emerged as a mechanism for resolving the free rider problem; I then tried to argue that such selection pressures for cooperative behaviour and its regulation were recruited by a religiosity which served to underpin it, but which was subverted to serve quite extraneous purposes. The foregoing formulation might appear to be a nod in the direction of group selection. This is emphatically not the case. That selection pressures operating at the individual level may well be seen as operating at the level of the group is well explained by Plotkin: “traits that favour the group (may) have higher fitness value for the ultimate survival of individuals in the group than traits that favour the individual at the expense of the group” (Plotkin, 1997, 229) my emphasis. This process was facilitated by those features of human developmental psychology which suggest the human mind is strongly predisposed to believe in gods and the supernatural. Further support for this process of the suborning of psychological predispositions, is provided by the role religiosity plays in harnessing the predilection of human cognition for, among other things, minimally counterintuitive representations, high levels of intensionality and a well developed hyperactive agent detection device. The archaeological record is, in part, the residue of ancestral costly signalling behaviour. Much of the archaeological evidence for ritual behaviour is most profitably to be interpreted as costly signals of the predisposition of those responsible for the creation of such artefacts to behave in a way which is consistent with what is understood to be the welfare of the group to whom the fabricator of the artefact belongs. I have attempted to integrate the 69 archaeological and neurological data using costly signalling theory and in doing so I have tried to realise this programme’s objectives by integrating both Palaeolithic archaeology and Palaeoanthropology. The diagram below is intended to illustrate this programme. Neuro-biology Fixation of COMT gene which codes for catechol-O-methyltransferase Inhibition of dopamine reward system making altruistic actions possible Psychology Beliefs in omniscient Gods (and ancestral proxies) militates against defection in social transactions-reinforcing the propensity to cooperate and not to defect. Anthropology Costly signals are an indication of sincere commitment to the group and a preparedness to practice reciprocal altruism Archaeology Artefactual record of costly signals (e.g. Parietal imagery, inhumation (later with grave goods) and Palaeolithic/Neolithic figurines, Iron Age votive offerings etc.) Fig. 26 Overview of integration of disciplines in an explanation of the significance of the artefactual record in terms of costly signalling I have tried to demonstrate that the neural substrate of religiosity is distributed and operates on a number of levels of functionality: genetic, neurochemical and the gross morphological features of the brain. Amongst these I have focused on the activation and inhibition of the dopamine pathway as this regulates the effectiveness of behaviours which favour prosociality. As such, it would have favoured those individuals who were predisposed to reciprocate altruistic acts rather than defect in social transactions. My overview of the archaeological record suggests it is moist effectively expressed within a religious context and I have attempted to indicate those features of 70 the archaeological and fossil record which evidence such behaviours. These would be expected to constitute a critical part of that repertoire of “cultural buffering” behaviours which gave modern humans an adaptive advantage in the European Upper Palaeolithic (Stringer, 2011) and which was not shared with the relatively, and fatally, disadvantaged Neanderthals (Henke and Tattersall, 2007). Although Dr McNamara supports my general programme, he has warned of the difficulty involved: “I think you are right to think that CST might be capable of integrating the archaeologic and neurologic data (but) it will be a hard road to do that.” (Pers. Com.) This dissertation marks only the beginning of the journey along this road, much work would need to be done to establish the role of all the neurotransmitters involved and the extent to which costly signalling explains the significance of the archaeological data I have discussed here. What I have tried to argue, is that a human default belief in gods was recruited to address the free rider problem in early human groups which had reached a size such that perceived genetic relatedness was no longer effective in deterring defection and rewarding cooperation. McNamara has indicated the enormous significance of this biological event in human evolution for the prosociality of humanity: “It may be that in the distant evolutionary past when the mutation that led to the COMT polymorphism regulating aminergic, particularly dopaminergic activity in the frontal lobes, our ancestors began to find that it was unusually easy to feel inclined to affiliate and cooperate with others.” (McNamara, 2006, 198) Now that the true significance of the role of COMT in securing prosociality is understood, further light will thrown on the origin of prosocial-man and the role religion has played in his prehistory. 71 Bibliography and References Aiello L., and P. Wheeler, 1995, “The Expensive-Tissue Hypothesis: the Brain and the Digestive System in Human and Primate Evolution”, Current Anthropology, 36: 199-221 Ano, G., and E. Vasconcelles, 2004, “Religious coping and psychological adjustment to stress: A meta-analysis”, Journal of clinical Psychology, vol. 61, issue 4, 461-480 Anselm of Canterbury, 1077-78, “Proslogion” Atkinson, and Wheeler, 2003, “Evolutionary Psychology's Grain Problem and the Cognitive Neuroscience of Reason” in “Evolution and the psychology of thinking: The debate”, D. Over (Ed.) Hove: Psychology Press, 61-99 (Retreived from the internet 14/03/2011) www.dur.ac.uk/a.p.atkinson/AtkinsonWheeler_2003preprint.pdf Atkinson, and Wheeler, 2004, “The Grain of Domains: The Evolutionary Psychological Case Against Domain-General Cognition”, Mind & Language, Vol. 19 No. 2, 147-176 Atran, S., 2002, “In Gods We Trust”, New York, Oxford University Press Atran, S., 2007, “Religion’s Social and Cognitive Landscape: An Evolutionary Perspective” in “Handbook of Cultural Psychology”, Kitayama, S., and D. Cohen (Eds), New York, The Guildford Press Austin, J.L., 1962, “How to Do Things with Words”, Oxford, Oxford University Press Axelrod, R., 1984, “The Evolution of Cooperation”, London, Penguin Books Azari, N. P., et al, 2001. Neural correlates of religious experience. European Journal of Neuroscience, 13, 1649-1652 Bahr, S., and J. Hoffman, 2008, “Religiosity, Peers, and Adolescent Drug Use”, Journal of Drug Issues, Vol.38, Issue 3, 743-770 Bailey, D., 2005, “Prehistoric Figurines: representation and corporeality in the Neolithic”, Oxford, Routledge Barrett. J., 2003, “Anthropomorphism or Preparedness? Exploring Children's God Concepts”, Review of Religious Research, vol. 44:3, 300-312 Barrett J., and J. Lanman, 2008, “The science of religious beliefs “, Religion 38, 109-124 Barriere,C., 1976, ” L'art parietal de la grotte de Gargas”, Parts I and II, Oxford, British Archaeological Reports, Beauregard, M., & Paquette, V., 2006, Neural correlates of a mystical experience in Carmelite nuns. Neuroscience Letters, 405, 186-190 Bering, J., and D. F. Bjorklund, 2004, “The Natural Emergence of Reasoning About the Afterlife as a Developmental Regularity”, Developmental Psychology, Vol. 40, No. 2, 217–233 Bering, J., 2006a,”The Cognitive Psychology of Belief in the Supernatural”, in “Where God and Science Meet”, vol. 1, P. McNamara (Ed.), Westport, CT, Praeger 123-134 72 Bering, J., 2006b, “The folk psychology of souls”, Behavioral and Brain Sciences, 29, 453–498 Bering, J., 2008, “The End? Why so many of us think our minds continue on after we die”, Scientific American Mind, 21, 60-63 Blackmore, S., 1999, “The Meme Machine”, Oxford, Oxford University Press Bliege Bird, R., and E. Alden Smith, 2004, “Signaling Theory, Strategic Interaction, and Symbolic Capital”, Current Anthropology, vol.46, number 2, 221-248 Borg, J., et al, 2003, “The Serotonin System and Spiritual Experiences”, The American Journal of Psychiatry, vol.160, 1965-1969 Bouchard, T., et al, 1990, “Sources of human psychological differences: the Minnesota Study of Twins Reared Apart” Science, Vol. 250 no. 4978, 223-228 Bouzouggar, A., et al, 2007, ““82,000-year-old shell beads from North Africa and implications for the origins of modern human behavior”, Proceedings of the National Academy of Sciences of the United States of America vol. 104 no. 24, 9964-9969 Bowles, S., 2006, “Group Competition, Reproductive Leveling, and the Evolution of Altruistic Behaviour”, Science 314, 1569-1572 Boyd, R., 2006, “The Puzzle of Human Sociality”, Science 314, 1555-1556 Boyer, P., 2001, “Religion Explained”, London, Vintage Boyer, P., 2003, “Religious Thought and Behaviour as by-products of brain function”, TRENDS in Cognitive Sciences Vol.7 No.3 March 2003 “British Social Attitudes, 2009”, 2010, National Centre for Social Research, London, “Religion and Ethnicity”, 70-71 Breuil, H., 1952, “Quatre cent siécles d’art pariétal”, Centre d’Études et de Documentation Préhistoriques, Dordogne, Montignac Brown, W., 2006, “The Brain Religion and Baseball: comments on the potential for a neurology of religion and religious experience” in “Where God and Science Meet”, vol.2, P. McNamara (Ed.), Westport, CT, Praeger, 87-121 Bulbulia, J., 2004, “The cognitive and evolutionary psychology of religion”, Biology and Philosophy, 19: 655–686 Butler, P., et al, 2011, “Disease-associated differences in religious cognition in patients with Parkinson’s disease”, Journal of Clinical and Experimental Psychology, 1-12 Bulbulia, J., 2006, “Nature’s Medicine: Religiosity As An Adaption for Health And Cooperation” in “Where God and Science Meet”, vol. 1, P. McNamara (Ed.), Westport, CT, Praeger, 87-121 Button, T., et al, 2010, “The Moderating Effect of Religiosity on the Genetic Variance of Problem Alcohol Use.”, Alcoholism: Clinical and Experimental Research, vol.34, issue 9, 16191624 73 Cadge, W., et al, 2009, “Religion and Spirituality: A Barrier and a Bridge in the Everyday Professional Work of Pediatric Physicians”, Social Problems, vol.56, No.4 Casteret, N., 1951, « Dix ans sous terre », Libraire Académique Perrin, Paris Chapman, J, 2001, “Object Fragmentation in the Neolithic and Copper Age of Southeast Europe”, in “The Archaeology of Cult and Religion”, (P. Biehl , F. Bertemes and H. Meller Eds.) Budapest, Archaeolingua, 89-105 Clottes J., and D Lewis-Williams, 1998, “The Shamans of Prehistory: Trance and Magic in the Painted Caves”, New York: Harry N. Abrams Coghlan,A., 2010, “Brain Shuts off in response to healer’s prayer”, New Scientist, Issue 2757, 9 Cohen. E., et al, 2010, “Rowers' high: behavioural synchrony is correlated with elevated pain thresholds”, Biology Letters, Vol.6, 106-108 Cohen, S., et al, 1991, “Psychological stress and Susceptibility to the Common Cold”, The New England Journal of Medicine, vol.325, no. 9, 606-612 Comings, D.E., et al, 2000, “The DRD4 gene and the spiritual transcendence scale of the character temperament index”, Psychiatric Genetics, 10:4 185-189 Cosmides, L., and J. Tooby, 1997, “Evolutionary Psychology: A Primer”, Center for Evolutionary Psychology, University of California, Santa Barbara (Retrieved from http://www.psych.ucsb.edu/research/cep/ on 08/07/11) Cox, S., T Sluckin and J Steele (1999) ‘Effects of group size, memory, and interaction rate on the evolution of cooperation.’ Current Anthropology 40: 369-377. D’Aquili, E., and A. Newberg, 1999, “The Mystical Mind”, Minneapolis, Fortress Press Darwin,C., 1981, “The Descent of Man”, Princeton, New Jersey, Princeton University Press, , [1871 Edition] Dawkins, R., 1976, “The Selfish Gene”, Oxford, Oxford University Press De Dreu, C., et al, 2010, “The Neuropeptide Oxytocin Regulates Parochial Altruism in Intergroup Conflict Among Humans”, Science, 328, 1408-1411 Defleur, A. et al, 1999, “Neanderthal Cannibalism at Moula-Guercy, Ardèche, France” Science, 286, 128-131 Dennett, D., 2006, “Breaking the Spell: religion as a natural phenomenon”, London, Allen Lane, Penguin Press D’Errico, F. and A. Nowell, 2000, “A New Look at the Berekhat Ram Figurine: Implications for the Origins of Symbolism” Cambridge Archaeological Journal, vol.10, Issue 1 Descartes, R., 1968, “Discourse on Method and Other Writings”, Harmondsworth, Middlesex, Penguin Books Donald, M., 1998, “Hominid enculturation and cognitive evolution”, In “Cognition and Material Culture: the Archaeology of Symbolic Storage” eds. C. Renfrew and C. Scarre, Cambridge: Macdonald Institute Monographs 74 D’Onoforio et al, 1999, “Understanding Biological and Social Influences on Religious Affiliation, Attitudes, and Behaviors: A Behavior Genetic Perspective”, Journal of Personality 67:6 953-984 Dunbar, I., 1993, “Coevolution of neocortical size, group size and language in humans”, Behavioral and Brain Sciences, 16, 681-735 Dunbar, I., 2003, “The Social Brain: Mind, Language and Society in Evolutionary Perspective” Annual Review of Anthropology, 32, 163-181 Durkheim, E., [1912], 2001, “The Elementary Forms of Religious Life”, Oxford, Oxford University Press Fodor, J., 1983, “Modularity of Mind: An Essay on Faculty Psychology”, Cambridge, Mass.: MIT Press Fincher, C., and R. Thornhill, 2008, “Assortative sociality, limited dispersal, infectious disease and the genesis of the global pattern of religion diversity”, Proceedings of the Royal Society B, 275, 2587-2594 Fingelkurts A.A., and A. A. Fingelkurts, 2009, “Is our brain hardwired to produce God, or is our brain hardwired to perceive God? A systematic review on the role of the brain in mediating religious experience”, Cognitive Processing, vol. 10, 293-326 Geyer, M. and F. Vollenweider, “ Serotonin research: contributions to understanding psychoses” Trends in Pharmacological Sciences, Volume 29, Issue 9, 445-453 Giordano, J., and J. Engebretson, 2006, “Neural and Cognitive Basis of Spiritual Experience: Biopsychosocial and Ethical Implications for Clinical Medicine”, Explore, Vol. 2, issue 3, 216225 Goren-Inbar, N and S. Peltz, 1995, "Additional remarks on the Berekhat Ram figure," Rock Art Research 12, 131-132 Griffiths, R., et al, 2006, “Psilocybin can occasion mystical-type experiences having substantial and sustained personal meaning and spiritual significance”, Psychopharmacology, 187:268–283 Grün, R., and C. Stringer et al, 2005, “U-series and ESR analyses of bones and teeth relating to the human burials from Skhul”, Journal of Human Evolution 49, issue 3, 316-334 Guthrie, S., 1980, “A Cognitive Theory of Religion”, Current Anthropology, Vol.21, No. 2, 181203. Hamer, D., 2004, “The God Gene: how faith is hard-wired into our genes”, New York, Doubleday Hamilton, W. D., 1964, "The Genetical Evolution of Social Behavior", Journal of Theoretical Biology 7(1): 1–52 Hannay, D., 2006, “Religion and Health”, Social Science & Medicine. Part A: Medical Psychology & Medical Sociology, Vol. 14, Issue 6, 683-685 Hansen, S., 2001, “Neolithic sculpture, Some Remarks on an Old Problem”, in “The Archaeology of Cult and Religion” (P. Biehl , F. Bertemes and H. Meller Eds.) Budapest, Archaeolingua 75 Harris, S., et al, 2009, “The Neural Correlates of Religious and Nonreligious Belief”, PLoS ONE 4(10) Harris, E., and P. McNamara, 2008, “Is Religiousness a Biocultural Adaptation?” in The Evolution of Religion: Studies, Theories, and Critiques” Edited by Bulbulia, J., et al, Santa Margarita, CA, The Collins Foundation Press Hawkes, C., 1954, “Archaeological Theory and Method: Some Suggestions from the Old world”, American Anthropologist, vol.56, issue 2, 155-168 Henke, W., and I Tattersall (Eds), 2007, “Handbook of Palaeoanthropology”, Berlin, SpringerVerlag Henshilwood, C., et al, 2002, “Emergence of Modern Human Behavior: Middle Stone Age Engravings from South Africa”, Science, vol.295, 1278-1280 Hayden, B., 2003, “Shamans Sorcerers and Saints”, Washington, Smithsonian Books Hooper, 1980, “Further information on the prehistoric representations of human hands in the cave of Gargas”, Medical History, 24(2): 214–216 House, J.S., et al, 1982, “The association of social relationships and activities with mortality: prospective evidence from the Tecumseh community health study.” American Journal of Epidemiology, 116:123-140 Hume, D., 1757, “The natural history of religion” cited by P. Bloom, “How Pleasure Works”, London, The Bodley Head, 2010, 113 Hummer, R., et al, 1999, “Religious involvement and U.S. adult mortality” Demography, 36(2):273-85 Inzlicht, M., et al, 2009, “Neural Markers of Religious Conviction”, Psychological Science, vol. 20, issue 3, 385 Israel, S., et al, 2008 “Molecular genetic studies of the arginine vasopressin 1a receptor (AVPR1a) and the oxytocin receptor (OXTR) in human behaviour: from autism to altruism with some notes in between”, Progress in Brain Research, Volume 170, 435-449 Jacobs, Z., and R. Roberts, 2009, “Human History Written in Stone and Blood”, American Scientist (online version retrieved from the internet 15/8/11) Janssens,P., 1957 'Medical views on prehistoric representations of human hands', Medical History, 1:318-322 James, W., 1902,”The Varieties of Religious Experience”, London, Longmans, Green, and Co Kant, Immanuel, [1781]1933, “Critique of Pure Reason”, A598-600:B626-628, trans. by Norman Kemp Smith, London, Methuen Kappelman, 1996, “The evolution of body mass and relative brain size in fossil hominids”, Journal of Human Evolution 30, 243–276 76 Kay, A., et al, 2010, “Religious Belief as Compensatory Control”, Personality and Social Psychology Review, 14 (1), 37-48 Kelemen, D., and Di Yanni 2005a, “Intuitions About Origins: Purpose and Intelligent Design in Children’s Reasoning About Nature,” Journal of Cognition and Development, 6(1), 3–31 Kelemen, D., and Di Yanni, 2005b, “Time to get a new mountain- The role of function in children’s conceptions of natural kinds”, Cognition, 97, 327-335 Kelemen, D., et al, 2005, “Why things happen: Teleological Explanation in Parent–Child Conversations”, Developmental Psychology, vol.41, No. 1, 251-264 Kelemen, D., 2007, “Reasoning about artifacts at 24 months-The developing teleo-functional stance”, Cognition, 103, 120-130 Kimbel, W., 2005, “Book Reviews: Holloway, R., et al, 2004, “The Human Fossil Record, Vol. 3 Brain Endocasts: The Paleoneurological Evidence, Hoboken, NJ: John Wiley and Sons”, Journal of anthropological Research, Vol. 61, No. 3 Klein, D., et al, 1976, “Learned Helplessness, Depression, and the Attribution of Failure”, Journal of Personality and Social Psychology Vol. 33, No. 5, 508-516 Knight, W., 2004, “Ostrich beads indicate early symbolic thought”, New Scientist, March 31st 2004 Koenig, L., and T. Bouchard, 2006, “Genetic and Environmental Influences on the Traditional Moral Triad – Authoritarianism, Conservatism, and Religiousness – as Assessed by Quantitative Behaviour Genetic Methods”, “Where God and Science Meet”, vol. l, (Ed. by P. McNamara), Westport, Connecticut, USA. Praeger Laming-Emperaire, A., 1962, La Signification de l’art rupestre paléolithique, Paris, Picard Leroi-Gourhan, A., 1968, “The evolution of Paleolithic art”, Scientific American, 209(2): 58-74 Leroi-Gourhan, A., 1967, « Les mains de Gargas. Essai pour une étude d'ensemble », Bulletin de la Société préhistorique française, Volume 64, Numéro 1, 107 - 122 Levin, J.S. and P. Schiller, 1987, “Is There a Religious Factor in Health”, Journal of Religion and Health, Vol. 26, No. 1 Levin J.S, and H.Y Vanderpool, 1987, “Is frequent religious attendance really conducive to better health? toward an epidemiology of religion”, Social Science and Medicine, 24:589-600 Levin J.S., and H.Y. Vanderpool, 1989,“Is religion therapeutically significant for hypertension?” Social Science and Medicine, vol. 29:69-7 Lewis Williams, J.D., 2002, “The Mind in the Cave”, London, Thames and Hudson Lewis Williams, J.D. and D.G. Pearce, 2004, “San Spirituality”, Walnut Creek, CA., USA, Altimira Press Lombroso, T., et al, 2007, “Inferring Design -Evidence of a Preference for Teleological Explanations in Patients with Alzheimer's Disease”, Psychological Science, 18 (11), 999-1006 77 Mackay and Welz, 2008, “Engraved ochre from a Middle Stone Age context at Klein Kliphuis in the Western Cape of South Africa”, Journal of Archaeological Science.35, 1521-1532 Marx, K., 1844, “A Contribution to the Critique of Hegel's Philosophy of Right”, Harmondsworth, Penguin Books Marshack, A., 1991, “The Taï Plaque and Calendrical Notation in the Upper Palaeolithic”, Cambridge Archaeological Journal , 1: 25-61 Maselko, J., et al, 2008, “Religious service attendance and spiritual well-being are differentially associated with risk of major depression”, Psychological Medicine, 39, 1009–1017 Matthews, D., et al, 1998, “Religious Commitment and Health Status: A Review of the Research and Implications for Family Medicine”, Archives of Family Medicine, vol.7, No.2, 114-185 McBrearty and Brooks, 2000, “The revolution that wasn’t: a new interpretation of the origin of modern human behaviour”, Journal of Human Evolution, 39 453–563 McKay, R., et al, 2011, “Wrath of God: religious primes and punishment”, Proceedings of the Royal Society B, 278:1858-1863 McNamara, P., 2002, “The Motivational Origins of Religious Practices”, Zygon, vol.37, no.1, 143-160 McNamara, P., 2006, “The Frontal Lobes and the Evolution of Cooperation and Religion”, Where God and Science Meet, P. McNamara (Ed.) vol. 2, Westport, CT, USA, Praeger, 189-204 Mead, S., et al, 2003, “Balancing Selection at the Prion Protein Gene Consistent with Prehistoric Kurulike Epidemics”, Science 300, 640-643 Mellars, P., 2006, “Why did modern human populations disperse from Africa ca.60, 000 years ago? A new model”, Proceedings of the National Academy of Sciences of the USA, vol. 103 no. 25, 9381-9386 Minkel, J., 2008, “Food for Symbolic Thought”, Scientific American, vol. 298, 30 Murray, D., et al, 2011, “On the Origins of Cultural Differences in Conformity: Four Tests of the Pathogen Prevalence Hypothesis”, Personality and Social Psychology Bulletin, vol. 37 no. 3, 318-329 Newberg, A., 2010, “Principles of Neurotheology”, Farnham, Ashgate Noble,W., and I. Davidson, 1996. Human Evolution, Language and Mind: a Psychological and Archaeological Inquiry. Cambridge: Cambridge University Press Oxman,T., et al, 1995, “Lack of social participation or religious strength and comfort as risk factors for death after cardiac surgery in the elderly”, Psychosomatic Medicine. 57:5-15 Pargament, K., 2000, “The many methods of religious coping: Development and initial validation of the RCOPE”, Journal of Clinical Psychology Volume 56, Issue 4, 519–543 Park, C., 2005, “Religion as a Meaning-Making Framework in Coping with Life Stress”, Journal of Social Issues, Volume 61, Issue 4, 707–729 78 Parsons, T., R. Bales and E. Shils, 1953, “Working Papers in the Theory of Action”, Glencoe, Illinois, USA, The Free Press Persinger, M., 1987, “Neurophysiological bases of God Beliefs”, New York, Praeger Petrovitch, O., 2007, “Key issues in the Study of Religion”, Psihologija, vol. 40, (3), 351-363 Pettit, P., 2002, “The Neanderthal dead: exploring mortuary variability in Middle Palaeolithic Eurasia”, Before Farming, 1 (4) Plato, 1970, “Euthyphro” (trans. By Benjamin Jowett) London, Sphere Books, 6e -10a Plotkin, H., 1997 “Evolution in Mind”, London, Allen Lane, The Penguin Press Powell, L., et al, 2003, “Religion and Spirituality: Linkages to Physical Health”, American Psychologist, Vol. 58, No. 1, 36 –52 Powell, et al, 2010, “Orbital prefrontal cortex volume correlates with social cognitive competence”, Neuropsychologia, Vol.48, issue 12, 3554-3562 Raghanti, M. A., et al, 2008a, “Cortical dopaminergic innervation among humans, chimpanzees and Macaque monkeys: a comparative study”, Neuroscience, 155, 203-220 Raghanti, M. A., et al, 2008b,” Differences in Cortical Serotonergic Innervation among Humans, Chimpanzees and Macaque monkeys: a comparative study”, Cerebral Cortex, 18, 584-597 Rappaport, R., 1999, “Ritual and Religion in the Making of Humanity”, Cambridge, Cambridge University Press Ravilous, K., 2010, “The writing on the cave wall”, New Scientist, issue 2748, 30-34 Read, D., and S. van der Leeu, 2008, “Biology is only part of the story …” Philosophical Transactions of the Royal Society B vol. 363 no. 1499 1959-1968 Reilly, G., 1998, “Stirring the Human Heart-the task of religious education”, The Furrow, Vol.49, No.3, 137-142 Renfrew, C., 2008, “Neuroscience, evolution and the sapient paradox: the factuality of value and of the sacred”, Philosophical Transactions of the Royal Society B, 363, 2041–2047 Rosas, A., et al, 2006, “Paleobiology and comparative morphology of a late Neandertal sample from El Sidrón, Asturias, Spain”, Proceedings of the National Academy of Sciences of the USA, vol.103, no.5, 19266 -19271 Rossano, M., 2007, “The Evolution of Conscious Experience: Ritual, Altered States And The Origins of Religion”, in “New Developments in Consciousness Research” (edited by V.W. Fallio), New York, Nova Science Publishers Inc. Ryle, G., 1949, “The Concept of Mind”, Harmondsworth, Penguin Books, Sahly, A., 1969 « Le problème des mains mutilées dans l’art préhistorique ». Thèse de Doctorat ès Lettres (Toulouse). Tunis 79 Saver, J.L., and J. Rabin, 1997, “The Neural Substrates of Religious Experience”, Journal of Neuropsychiatry and Clinical Neurosciences, vol.9, no.3, 498-510 Schjødt, U. et al, 2008, “Rewarding Prayers”, Neuroscience Letters 443 (2008) 165–168 Schnall, E., et al, 2010, “The relationship between religion and cardiovascular outcomes and allcause mortality in the women’s health initiative observational study”, Psychology and Health, vol.25, No.2, 249-263 Shariff, A., and A. Norenzayan, 2007, “God Is Watching You: Priming God Concepts Increases Prosocial Behavior in an Anonymous Economic Game”, Psychological Science, Vol. 18.Number 9, 803-809 Shermer, M., 2000,”How we Believe: science scepticism and the search for God”, New York, Henry Holt and Company Singer, P., 1981 “The Expanding Circle”, Oxford, Oxford University Press Snyder, S., 2006, “Commentary on: Psilocybin can occasion mystical-type experiences having substantial and sustained personal meaning and spiritual significance by Griffiths et al”, Psychopharmacology, vol. 187, 287-288 Sosis, R., and E. Bressler, “Cooperation and Commune Longevity: A Test of the Costly Signaling Theory of Religion”, Cross-Cultural Research, Vol. 37 No. 2, May 2003, 211-239 Sosis, R., and B. Ruffle, 2003, “Religious Ritual and Cooperation: Testing for a Relationship on Israeli Religious and Secular Kibbutzim”, Current Anthropology, 713-22 Sosis, R., 2006, “Religious Behaviours, Badges and Bans”, “Where God and Science Meet”, vol. 1, (Ed. by P. McNamara), Westport, Connecticut, USA Praeger, 61-86 Stallings, M., et al 1996, “Genetic and Environmental Structure of the Tridimensional Personality Questionnaire: Three or Four Temperament Dimensions?” Journal of Personality and Social Psychology, vol. 70, No. 1, 127-140 Steadman, S., 2009, “Archaeology of Religion: cultures and their beliefs in worldwide context”, Walnut Creek, California, Left coast Press Inc. Stringer, C., 2011, “The Origin of Our Species”, London, Allen Lane, Penguin Books Smirnov, Y., 1989, “Intentional Human Burial: Middle Paleolithic (Last Glaciation) Beginnings”, Journal of World Prehistory, vol.3, no. 2, 199-233 Trivers, R.L., 1971, "The evolution of reciprocal altruism", Quarterly Review of Biology 46: 35– 57 Ucko, P., 1969, “Ethnography and Archaeological Interpretation of Funerary Remains”, World Archaeology, Vol. 1, No. 2, 262-280 Uffe Schjoedt, et al, 2011, “The power of charisma—perceived charisma inhibits the frontal executive network of believers in intercessory prayer”, Social Cognitive and Affective Neuroscience 6(1): 119-127 80 Vanhaeren, M., et al, “Middle Paleolithic Shell Beads in Israel and Algeria”, 2006 Science, vol. 312, 1785-88 Van Rossum, J. 1967, “Neuropsychopharmacology, Proceedings Fifth Collegium Internationale Neuropsychopharmacologicum”, Brill H, Cole J, Deniker P, Hippius H, Bradley P B, editors. Amsterdam: Excerpta Medica. pp. 321–329 Verhoeven, M., 2004, “Beyond Boundaries: Nature, Culture and a Holistic Approach to Domestication in the Levant”, Journal of World Prehistory, Vol. 18, No. 3, 179-282 Vyse, S. A., 1997, “Believing in Magic: the psychology of Superstition”, Oxford, Oxford University Press Walter, M. and E. Fridman, 2004, “Shamanism: an encyclopedia of world beliefs, practices, and culture, Volume 2”, Santa Barbara, California, ABC-CLIO Way, B., and M. Lieberman, 2010, “Is there a genetic contribution to cultural differences? Collectivism, individualism and genetic markers of social sensitivity”, Scan, 5, 203-211 Wengrow, D., 2011, “Cognition, materiality and monsters: the cultural transmission of counterintuitive forms in Bronze Age societies”, Journal of Material Culture”, 16 (2) 1-9 West, S., 2001, “Testing Hamilton's rule with competition between relatives”, Nature, vol. 409, 25, 510-513 West, S., et al, 2010, “Sixteen common misconceptions about the evolution of cooperation in humans”, Evolution and Human Behavior, 31, 1-32 Whitehouse, H., 2004, “Modes of Religiosity: A Cognitive Theory of Religious Transmission”, Walnut Creek, California, Altamira Press Whitehouse, H., and R. McCauley, (Editors), 2005, “Mind and Religion”, Walnut Creek, California, Altamira Press Whitson, J., and A. Galinsky, 2008, “Lacking Control Increases Illusory Pattern Perception”, Science, Vol. 322 no. 5898, 115-117 Wolpert, L., 2006, “Six Impossible Things Before Breakfast: The Evolutionary Origins of Belief”, London, Faber Wrangham, R., 2010, “Catching Fire; how cooking made us human”, London, Profile Books Zac, P., 2007, et al, “Oxytocin Increases Generosity in Humans”, PLoS ONE 2(11) Zimmer, C., 2004, “Faith-Boosting Genes: A search for the genetic basis of spirituality”, Scientific American, 291, 110-111 Zorich, Z., 2011, “A Chauvet Primer”, Archaeology, Volume 64 Number 2 81 Web sites consulted http://www.adherents.com/Religions_By_Adherents.html (Retrieved 1/10/10) http://www.apollon.uio.no/vis/art/2006_4/Artikler/python_english (Retrieved 26/04/2011) http://www.bradshawfoundation.com/clottes.page4/php (Retrieved 05/06/11) http://www.bradshawfoundation.com/hand_paintings_rock_art.php (Retrieved 14/04/11) http://www.britannica.com/EBchecked/topic/1581715/religion-Year-In-Review2009/286517/Worldwide-Adherents-of-All-Religions (Retrieved 3/10/11) 82 Appendix 1 Copy of my criticisms of Dr Wengrow’s paper Criticisms of Dr Wengrow’s paper* 1. You suggest Boyer’s explanation for the “stickiness” of counterintuitive concepts should be born out by a significant distribution of monsters in the archaeological record, but is this a misrepresentation of Boyer? Boyer talks about minimally counterintuitive concepts, not multiply counterintuitive concepts which monsters embodying several species drawn from the domain of “folk biology” would be. You admit that your definition admits of “blending elements from two or more species” but is this what Boyer has in mind? Nowhere in your paper do you qualify the notion of counterintuitive concepts with “minimally”. 2. Boyer does not argue that being counterintuitive is both necessary and sufficient for the concept to gain cultural traction; if his claim is that it is only necessary for a concept to be minimally counter-intuitive, then his claim is not inconsistent with the archaeological evidence that different combinations/permutations are discontinuously present in the artefactual record (both chronologically and geographically) This leaves open the operating of just those “technological and institutional” variables you discuss in your paper. Boyer does though seem to be maintaining that these other variables are psychological rather than material, and in this respect, your paper has proved, for me, a valuable corrective to Boyer’s rather narrow range of exclusively psychological causal factors in explaining the epidemiology of religious representations. In Religion Explained he says, “the (cultural concepts) that we find widespread in many different cultures and at different times probably have some transmission advantage, relative to several different mental dispositions.”(my emphasis) (Boyer, P. Religion Explained, 2002, Vintage, London, 57) 3. One of your main criticisms of Boyer is that there is so little evidence of composite imagery dating from the prehistoric / pre-urban/ pre-elite period which is not what would be predicted if counterintuitive representations are a function of pan-human cognitive biases. Is this contention not undermined by the fact that you have already conceded earlier in the paper that taphonomic bias can be held responsible for this distortion of our understanding? You quote Napier sympathetically in this regard on p.4 4. Am I correct in seeing the possibility of incorporating your approach into Stephen Shennan’s published work† on the demographic determinants of cultural diffusion? 5. Boyer discusses the memorability of counterintuitive beliefs as embedded components of narratives not as isolated phenomenon. Scott Atran would seem to acknowledge this where he asks why, if counterintuitive beliefs are so memorable, they are not more ubiquitous: ”An answer to this puzzle may lie in examining the memorability of an entire set of beliefs as a single unit of transmission rather than individual beliefs”(my emphasis) (Atran, 2002, “In Gods We Trust”, 101,Oxford, OUP) 83 *Wengrow, D., 2011, “Cognition, materiality and monsters: the cultural transmission of counterintuitive forms in Bronze Age societies”, Journal of Material Culture”, 16 (2) 1-9 † Powell, A., et al, 2009, “Late Pleistocene Demography and the Appearance of Modern Human Behavior”, Science, Vol. 324, 1298-1301 Appendix 2: Limitations of Shaminstic explanation for Palaeolithic “cave art” Firstly, the argument from analogy is not sound. The argument that you can infer the narrative significance of imagery in places like Chauvet from similar images in San rock art in Africa relies on the premise that practices familiar to present day San, for example, were identical to those depicted in South African rock art. This identity would have to be established independently of the evidence for the historic practices of the San but this data is not available to us for Franco Cantabrian Palaeolithic imagery whereas it is corroborating evidence (oral tradition and artefactual) for those who study south African rock art or Australian aboriginal art. This argument from analogy is logically flawed. Steadman who is typical of those who champion this analogous approach concedes at one point that, “Without knowledge of present day Bushmen religion and ritual, the rich detail of the rock art images would certainly be lost to us.” (Steadman, 2009, 86) yet seems unwilling to infer the logically entailed corollary that because we have no comparable data from the descendants of the Chauvet cave image makers, we are not in a position to make claims about their religion and ritual. The fact that he is able to speculate about the narrative significance of San and Aboriginal rock art is because the ethnographic data is available to him from those descended from the very people responsible for the imagery; this is not the case with similar attempts to interpret the narrative significance of Franco-Cantabrian rock images. Any information from the present day inhabitants of the Franco-Cantabrian region would have absolutely no bearing on those images for reasons largely consequent from the genetic unrelatedness of the latter with the former; they are not the same people nor were the practices of the image makers of Chauvet transmitted to the ancestors of the present day inhabitants of the region. 84 Secondly, not only are the present day inhabitants of Australian and South African descended from those who created the rock art, these present day descendants inhabit a similar ecological niche to that of their forebears which makes such any interpretation they offer more likely to be the case than not. The image makers of Chauvet and the present day inhabitants of the FrancoCantabrian region do not. Thirdly, how likely is it that such radically different ecosystems would give rise to identical ritual practices? The twentieth century tribesmen inhabiting desert/savannah ecology in southern Africa are decidedly not occupying a niche identical to that of the Palaeolithic hunter gatherers in the ice age ecology of the European Palaeolithic. Fourthly, the ethnographic evidence drawn from South Africa and Australia is provided by peoples (who for several reasons) we are entitled to believe are intimately familiar with the religious and ritual significance of the creators of the rock art of the past; they practice those same rituals. This continuity is entirely absent in putative European parallels. Fifthly, the proportion of Franco-Cantabrian imagery devoted to scenes which have been held to embody shamanistic practice is very small indeed. This is unlikely to be the case if the main motivation of the Cro-Magnon cave painters was to record shamanistic practices. Anthropomorphic imagery is rare, even when entopic images are included as being typically anthropogenic in origin. If such imagery served shamanistic purposes this proportion would surely be much greater. As well as constituting a very small proportion of the total images, their depiction is often cursory and lacking the same attention to detail which depictions of animals seemed to merit. Jean Clottes himself concedes that, “They deliberately chose to represent vague humans, with few details or deformed features.” (bradshawfoundation.com/clottes.page4/php). Finally, those caves which do contain anthropomorphic imagery and have the strongest claim to represent shamanistic practices occur in spaces where there is very little evidence of 85 congregating humanity. In view of the very public context in which shamanistic activity take place, these small and relatively unfrequented locations are not what would be predicted for spaces that are centres of such activity. That the locations of these images are problematic for any theory which emphasises the role of shamans in an explanation of Palaeolithic “cave art”, is recognised by researchers such as Rossano, who finds it, “…intriguing how frequently cave artists intentionally selected tiny, tight, difficult-to-reach recesses for their work.” (Rossano, 2007, 108) The calculated affect on the audience having observed the trance state of the socalled shaman seems critical to its effective functioning but evidence of this is absent in the locations chosen to depict such rituals. Countable text (- Abstract, Figs, Acknowledgements, Bibliography and Appendices) = 16499 words 86