Lesson 3: Species in the environmental complex
advertisement

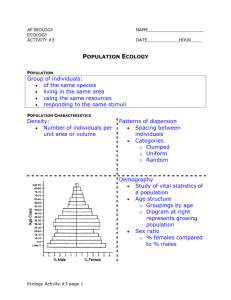
Lesson 3: Species in the environmental complex • Environmental factors and plant distribution – Liebigs law of the minimum – Shelfords law of tolerance – Billings Holocoenotic concept • Ecotypic variation within species: – Morphological variation – Ecophysiological variation – Genetic variation Why are organisms absent some places and abundant in others? Why are organisms absent some places and abundant in others? • Abiotic Forces: plants have certain limits regarding environmental tolerances. –Temperature, moisture, sunlight, pH, substratum, salinity, atmosphere, etc • Biotic Forces: Organisms compete for space and resources, and some eat others. –Competition, predation, symbiosis, nest sites, habitat modification. • Opportunity (history): organisms are absent where there is no geographical access. –Sometimes when they are introduced into areas where they were previously absent, havoc ensues. –introduced (exotic) species are the second most important cause of extinction. Habit destruction is the first. –Examples: kudzu, fire ants, killer bees, zebra mussels, Asiatic clams, Japanese honeysuckle, water hyacinth, Hydrilla, Egeria, Japanese beetles, honeybee mites, giant slugs, Brazilian pepper, tamarisk, thistles, Australian pine, privet. Justis Liebig (1803-1873) Liebig’s Lab in Giessen, 1840 Liebigs Law of the Minimum • • • Liebig recognized that plant growth is controlled by plant nutrients, and whichever nutrient is in most limited supply controls the plants growth. This concept was later expanded to cover other environmental factors such as water, light, temperature. He noted that plants had varying ranges of tolerance for a given factor. • “The growth or distribution of a plant is dependent on the one environmental factor most critically in demand.” Shelfords Law of Tolerance • • • • Shelford in 1913 noted a weakness in Liebigs general law which came to be known as the Law of Tolerance. And this in turn was modified by Ronald Good, a plant geographer: “Each and every plant species is able to exist and reproduce successfully only within a definite range of environmental conditions. Good rated climatic factors above edaphic factors. Some Examples: – Salinity tolerance in plants (intertidal zonation) – Thermal constraints on activity (sparrow microclimates, shown in the diagram from Smiths Ecology and Field Ecology.) Shelfords Law of Tolerance • From Smith’s Ecology and Field Ecology. The distribution of a species along an environmental gradient generally approximates a Gausian distribution, with the optimal occurrence somewhere near the midpoint of the distribution. Steno- and Eury• • These prefixes describe the width of the ecological niche in relation to a given environmental factor. For example, A plant with a narrow range of tolerance for temperature is termed stenothermic. Plants with a wide thermal tolerance are termed eurythermic. -phile and -phobe • • These suffixes are used to describe plants that favor one extreme of an environmental gradient. For example, a plant favoring snowy habitats is called a chionophile, and a plant that grows only in windblown, snowfree habitats would be called a chionophobe. Plants operate within the tolerance limits for all the critical factors for their growth • • Add Fig. 3-1 (BBPGS) No single factor controls the distribution of a plant. • Good thought that climate factors were most important controls, edaphic (soil) factors were next most important, and biotic factors, such as competition, were least important. Biotic Controls: Competitive exclusion in animals: Gauses experiment using protozoans (Paramecium) • .Competition with other species can strongly influence how a species will respond to an environmental factor. • The concept of what defines a species niche is one of the most hotly disputed topics in ecology. This famous experiment by the Russian biologist G.F. Gause (1934) first demonstrated the principal of “one species, one niche”. He did the experiment using small animals. Biotic Controls: Competitive exclusion in animals: Gauses experiment using protozoans (Paramecium) • P. aurelia • • P. caudata Grown separately Grown together Based on Gause 1934 In separate but identical bacterial cultures. Each species showed a similar growth rate in the absence of competition. When placed together, P. aurelia was able to compete successfully for the bacterial food, and eventually eliminated P. caudata. In another experiment P. caudata was grown with P. bursaria. Both species fed on the bacteria, but in this case, P. bursaria fed on the bacteria at the bottom of the tub, while P. caudata fed on the bacteria in suspension-Coeexistance because of different niches. Competitive exclustion in plants: Harpers experiment using duckweed (Lemma) Adapted from Harper 1961 Effects of competition: shift of ecological optimum • Ellenberg showed that the growth of two species of mustard have very similar response to pH when grown separately. • When grown together in mixed culture, they both shift their optimum pH a bit. Spergula favors a lower pH, and Raphanus favors a higher pH. • In most cases it is unlikely that they will have exactly the same niche, where one totally excludes the other. Solid line: Spergula arvensis (Spurry) Dashed line: Raphanus raphanistrum (a wild mustrard) From Ellenberg 1958 The ecological (or realized) niche for a species may shift or be truncated in the presence of competition. • Add Fig. 3-3 (BBPGS) Another view of fundamental vs. realized niches From Smith 1977 • The presence of other species in portions of the range of characteristics, can eliminate Species A from parts of its fundamental niche, so that its realized niche is smaller. Dwight Billings • Dwight Billings helped to crystallize the study of the relationship of plants to the environment. He was one of the major proponents of using an ecophysiological approach. • He believed that the best way to study these relationshps was through detailed autecological studies of plant species and how they react to changes in their environment. Why do plants grow where they do? The holocoenotic environmental complex W.D. Billings, 1952. The environmental complex in relation to plant growth and distribution. Quaterly Review of Biology 27: 251-265. • A complex, indvisible whole system consisting of the plant and its multitude of environmental influences. • Holocoenosis = ecosystem? • Compare to Tansleys concept of the “ecosystem”, which also included time and change. • Billings concept is therefore an ecosystem at a moment in time. Billings: Groups of factors in a terrestrial plant environment: – – – – – – Climate Edaphic Geographic Topographic Pyric Biotic Groups were subdivided into: factors subfactors and aspects. Examples: Climate was divided into the factors Radiation, Temperature, Water, Atmospheric gases. Edaphic was divided into Parent material, Soil; Geographic was divided into Gravity, Rotational Effects, Geographic Position, Vulcanism, Ditrophism (folding, faulting), Erosion and Deposition, Topography, etc. The factor Radiation was divided into the subfactors: Solar radiation, cosmic radiation, and terrestrial radiation. The subfactor Solar radiation was subdivided in the aspects, wavelenths, intensity, photoperiod and other cycles. Billings identified a total of 64 environmental aspects, but this was by no means a comprehensive list. It only served to illustrate how complex the plant environment is and how the aspects interact among themselves and the plant. Some conclusions from Billings paper: Factors of a terrestrial plant environment: • Environment of a plant is holocoenotic (forms a complete system in combination with the plant). • For a given species, limiting factors can be different in different parts of its range. • The total environment is dynamic and varies both space and time. • Vegetation can be used as an indicator of the total environment if the tolerances of its characteristic species are known. What is a species?: The biological species concept • A group of natural populations that are morphologically, genetically, and ecologically similar. – This definition involves • Appearance (morphology) • Breeding behavior (genetics) • Habitat distinctiveness (ecology) • They may or may not be interbreeding, but they are reproductively isolated from other species. The biological species concept • Classical taxonomists have focused most on morphology. This approach has been until recently the only tool to determine the relationship of one species to others. • Newer approaches to the species focus on the genetic aspects -how population of plants maintain their distinctiveness through genetic isolation. • These isolating barriers may be due to: – breeding behavior (time of flowering, type of pollinator), – habitat, or geographic isolation, or – inability to form fertile hybrids. Ecotype Concept • Many botanists have noted that within a given species there is often considerable morphological, physiological, or phenological variation. • Kerner in Switzerland noted such variation but thought that the various traits were plastic responses to environmental factors. • In the 1920s Göte Turesson confirmed that many of the traits were heritable. He collected samples of populations of many plant species from all over Europe and grew them in a common garden in Sweden. He noted considerable variation in a variety of factors within a given species even though the various populations were fully interbreeding. Göte Turreson: Ecotype Concept Table 3-2. From Barbour et al. 1999. • Turreson grew plants of the same species from all over Europe in his transplant garden in Åjarp, Sweden. • He noted within one species of hawkweed collected from woodlands, fields, and dunes, there was consistent variation in leaf morphology, pubescence, and autumn dormancy. Turresons observations of Betula . ALPINE TUNDRA MID-ELEVATION FOREST COASTAL BLUFFS LOW TEMP (SPRING) MOD TEMP (SUMMER) MOD. TEMP (SUMMER) PROSTRATE TINY LEAVES • TREE LARGE LEAVES SHRUB FLESHY LEAVES THESE DIFFERENCES PERSISTED IN THE COMMON ENVIRONMENT GARDEN LEADING GOTE TURESSON TO LABEL EACH POPULATION AN ECOTYPE OF ITS SPECIES Key aspects of ecotypes according to Turresons concept • Wide-ranging species are differentiated into different hereditary groups that are genetically based (ecotypes). They are discrete entities with clear differences separating them from other ecotypes. • The genetic differences are adaptations to the different habitats. • Ecotypes occur in distinctive habitats. In a given habitat populations of different species often exhibit similar morphological and developmental characteristics. Distinctiveness can be morphological, physiological, and/or phenological. (e.g. (1) life form, (2) timing of growth, (3) tolerance of frost, (4) tolerance of salt, and (5) tolerance of shade.) • They are potentially interfertile with other ecotypes of the same species. Clausen, Keck, and Heisey (1940) • • • • Fig. 3.5. Barbour et al. 1999. Ecotypes along an elevation transect in California. Initially, they had many transplant gardens located along this transect and were transplanting about 180 species from different areas into as many gardens as possible. Reduced the number of gardens to the three at Stanford, Mather, and a Timberlilne site, and 60 species. Transplanted local species from each site into the other gardens. Summary of environmental conditions at the transplant gardens Table 3.3. Barbour et al. 1999. Three ecotypes (subspecies) of Potentilla glandulosa and their appearance in each of the transplant gardens Ecotype • Photos along the diagonal show the species as it grows at its native site. • All of the populations were shown to be interfertile. • They concluded, as Turesson did, that species are really composed of genetically distinct groups of ecotypes which are best suited their specific environment. Nevadensis reflexa typica Fig. 3-6 from Barbour et al. Mooney and Billings (1959): Ecophysiological variation in ecotypes of Oxyria digyna (Mountain sorrel) • Took the concept one step further with detailed examination of the physiological responses of different population. Morphological, biochemical, and phenological differences between the arctic and alpine populations of O. digyna • Insert Table 3-6 from BBPGS Physiological response of O. digyna ecotypes: Photosynthetic response Temperature varies Light intensity varies Fig. 3-8, Barbour et al. 1999 • CO2 uptake of two ecotypes at different temperature and light conditions. • The alpine plants had higher photosynthetic response to both temperature and light, reflecting the generally warmer and sunnier conditions in the alpine environment. • The optimum point was also higher in each case for the alpine plants (i.e., higher light intensity and higher temperature. • Ecophysiological differences have now been documented at much more local scales, between geographically close populations, consistent with idea of ecoclines. Ecocline concept • Langlet (1959) examined the growth of Pinus sylvestris at 580(!) sites in Sweden and concluded that species really formed a cline, or continuum of variation. • Quinn (1987) showed that every population of Danthonia caespitosa had distinctive growth characteristics (phenology, morphology, physiology). He concluded that each population was in some sense individualistic. McNaughton (1966): enzymatic connection in Typha latifolia • • http://plants.usda.gov/ • • • • • Fig. 3-9 , Barbour et al. 1999 Another step closer to the demonstrating actual genetic control for ecotypic variation of physiological differences. Point Reyes: foggy coastal site. Red Bluff: hot Sacramento Valley. Collected dormant rhizomes from each site and placed in common greenhouse. Made plant extracts and collected 3 enzymes. Subjected to the enzymes to heat stress of 50 ˚C for up to 30 min. One of the critical enzymes (malate deydrogenase) from Red Bluff showed much higher activity with higher temperature. Other studies the helped demonstrate differences in ecotypes at the genetic level • Smith and Pham (1996) using molecular biology showed high genetic variation in separate populations of wild onion. • McCauley et al. (1996) examined spatial patterns of chloroplast DNA in Silene alba, and found the most similar genotypes were those in close proximity to each other. • Rejmanek (1996) found major differences in the amount of nuclear DNA in different pine species. Aggressive invaders of pines had small amounts of nuclear DNA, which correlated with a shorter time for cell division, more rapid growth. • Techniques of molecular biology have now been used to identify differences in specific genes. Rapid evolution of heavy metal tolerance in bent grass (Danthonia tenuis). Anthony Bradshaw and students (1996): • Studied mines in Wales abandoned about 100 yr ago, and contaminated with high levels of Cu, Fe, Zn, and Pb. Heavy metals cause proteins to precipitate leading to death. • Bent grass was one of the few species growing on mines and away from mines. • Bradshaw grew plants in concentrations of 20 and 2000 ppm of heavy metals. Plants on the mine sites grew in both solutions, but plants on other sites grew only in 20 ppm solution. • Tolerance is present in 0.3% of natural population but is strongly selected for on the mine sites. • Plants with the tolerance make specific proteins to bind the heavy metals, and this takes energy away from growth of leaves and roots, making these plants less competitive in natural populations. http://www.planning.sa.gov.au/ See: 1. Bradshaw, A.D. and McNeilly, T. 1996. Evolution and pollution. London: Edward Arnold. 2. Salt, D.E. et al. 1995. Bio/Technology 13: 468-474. 3. Adler, T. Science News. 1996. 150:4243. McGraw and Antonovics study of Dryas octopetala • • Examined the causes of ecotypic variation in ssp. octopetala and ssp. alaskana An integrated approach to study plant populations using: – – – – – – Growth chamber studies Field transplants Competition trials Pollination ecology Photosynthesis measurements Determination of photosynthate allocation patterns in plant organs McGraw, J.B. 1985b. Experimental ecology of Dryas octopetala ecotypes. III. Environmental factors and plant growth. Arctic and Alpine Research, 17: 229239. Summary • Environmental factors and plant distribution – – – • Liebigs (1840) law of the minimum Shelfords (1913) law of tolerance Holocoenotic concept of the the plant environment (Billings 1952) Ecotypic variation within species: – – – – Morphological variation (Turesson 1920s, Clausen, Keck and Heisey 1940, Langlet 1959, Quinn 1987) Ecophysiological variation (Mooney and Billings 1961, McNaughton 1967; Björkman 1968) Genetic linkage identified (e.g., Rejmanek 1995) Rapid selection for genes and adaptation to highly toxic environments (Bradshaw and students) Literature for Lesson 3 • • • • • • • Billings, W.D. 1952. The environmental complex in relation to plant growth and distribution. Quarterly Review of Biology 27: 251-265. Björkman, O. 1968. Carboxydismutase activity in shade-adapted species of higher plants. Physiologia Plantarum 21:1-10. *McGraw, J.B. and J. Antonovics. 1983. Experimental ecology of Dryas octopetala ecotypes. Ecotypic differentiation and life-cyle stages of selection. J. Ecol.: 879-897. *McGraw, J.B. 1985b. Experimental ecology of Dryas octopetala ecotypes. III. Enviornmental factors and plant growth. Arctic and Alpine Research, 17: 229-239. McNaughton, S. J. 1966. Thermal inactivation properties of enzymes from Typha latifolia L. ecotypes. Plant Physiology 41: 1736-1738. *Mooney, H.A. and W.D. Billings. 1961. Comparative physiological ecology of arctic and alpine populations of Oxyria digyna. Ecological Monographs 31: 1-29. Rejmanek, M. 1996. A theory of seed plant invasiveness: the first sketch. Biological Conservation 78: 171-181.