Lichens : a chemically important biota
advertisement

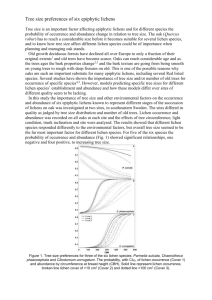
J. Natn.Sci.Foundation Sri Lanka 2005 33(3): 169-186 REVIEW LICHENS: A CHEMICALLY IMPORTANT BIOTA VERANJA KARUNARATNE", KARUNANANDA KATHIRGAMANATHAR and VINITHA M. THADHANI BOMBUWELA, SELVALUXMY Department of Chemistry, University of Peradeniya, Peradeniya. (Accepted: 03 October 2005) Abstract: Lichens produce a wide variety of secondary metabolites which assist them in the colonization of terrestrial h a b i t a t s . S u c h compounds a r e i m p o r t a n t i n t h e chcmotaxonomy of lichens. Furthermore, lichen substances exhibit antiherbivore, antimicrobial and antineoplastic activities. This review highlights the different types of lichen substances and their activities, uses and t h e possible ecological role. Lichens a r e symbiotic organisms of fungi (mycobiont) and algae (photobiont) comprising about 17,000 species. Lichens are unique because they look and behave quite differently from their component organisms. They are classified and named according to their fungal partner, with many thousands of species of lichens sharing a much smaller number of photobiont species. Lichens show a world-wide distribution commonly growing on rocks and poorly developed soils such as those of arid lands and boreal-arctic regions or as epiphytes on trees and shrubs. Primary metabolites of lichens, which are intracellular, are proteins, amino acids, polyols, carotenoids, polysaccharides and vitamins. Lichens produce a wide array of secondary metabolites (extracellular). There are over 700 lichen substances reported to date1and many are restricted to t h e lichenised s t a t e . Broadly speaking, there are three major types of lichen substances based on their biosynthetic pathway^.^ (a) Depsides, depsidones and dibenzofurans formed by the acetate-malonate pathway. The most important of these are the esters and the oxidative coupling products of simple phenolic units related to orcinol and p-orcinol. Most depsides and depsidones are colourless compounds which occur in the medulla of the lichen. However, usnic acids, yellow cortical * Corresponding author compounds formed by the oxidative coupling of methylphloroacetophenone units are found in the cortex of many lichen species (Figure 1). Anthraquinones, xanthones and chromones, are all pigmented compounds which occur in the cortex, which are also produced by the acetate-malonate pathway by intramolecular condensation of long folded polyketide units rather than the coupling of phenolic units (Figure 2). (b) The shikimic acid biosynthetic pathway produces two major groups of pigmented compounds,which occur in the cortex: pulvinic acid derivatives and terphenyl quinones (Figure 3). Although most pulvinic acid derivatives lack nitrogen, they a r e biosynthesized through phenyl alanine. Nitrogen is strongly limiting to metabolic activities in most lichens, and nitrogen rich metabolites such as alkaloids are unknown among lichen substances. (c) Terpenes and steroids are produced by the mevalonic acid pathway. These include compounds unique to lichens and many that occur in higher plants as well (Figure 4). Chemotaxonomy of lichens The application of chemical discriminators to lichen taxonomy is widely accepted. The secondary metabolites in about 5000 species (33 % of all known species) have been studied and the data are used in the routine identification of lichens than in any other group of organism^.^^^ Cortical chemistry: It has now been recognized by many lichenologsts that some cortical substances Veranja Karunaratne et al. Alectorialic acid (Benzyl depside) Lecanoric acid (para-Depside) Sekikaic acid (meta-Depside) Gyrophoric acid (Tri depside) Micareic acid (Diphenyl ether) Didymic acid (Dibenzofuran) Usnic acid Virensic acid (Depsidone) Picrolichenic acid (Depsone) Contortin (Biphenyl) Figure 1: Structures of typical acetyl-polymalonyl lichen products derived from two or more polyketide chains. Lichens: a chemically important biota Aliphatic acids H02C, H ,C02H (CH2)11CH3 H3C (+)-Protolichesterinicacid H - " Roccellic acid Mononuclear phenolic compounds Orsellinic acid Siphulin (Chromone) Haemoventosin (Naphthaquinone) POrsellinic acid Lichexanthone (Xanthone) Averythrin (Anthraquinone) Figure 2: Structures of some acetyl-polymalonyl lichen products derived from a single polyketide chain. Veranja Karunaratne et al. 172 Pulvinic acid derivatives Pulvinic acid Calycin Terphenyl quinones Polyporic acid Indolyl-3-acetic acid Thelephoric acid Ambewelamide A, 1 R = (CH,),CH, Ambewelamide B, 2 R = (CH,),CH, Figure 3: Structures of some lichen products derived from the shikimic acid pathway. Terpenoids (-)-p-Thujone(Monoterpenoid) (-)-Sandaracopimaricacid (Diterpenoid) Fukinanolide (Sesquiterpenoid) Hopan-Ga,7a,22-triol (Triterpenoid) Lichens: a chemically important biota Ergosterol (Steroid) y-Carotene (Carotenoid) Figure 4: Structures of typical lichen products derived from the mevalonic acid pathway. are correlated with higher taxonomic ranks. For example, vulpinic acid in Letharia (at the genus level) or anthraquinones such as parietin in Teloschistaceae (at the family l e ~ e l ) . ~ Medullary chemistry Variations in medullary constituents are used mainly as discriminators at the species level. Most morphologically defined species have a constant chemistry, regardless of their geographical location, substrate or ecology. Within a complex of morphologically similar species, three common patterns of chemical variation can be discerned: (a) replacement compounds, (b) chemosyndromic variation and (c) accessory type compound^.^ With replacement compounds, cogeneric chemotypes show simple replacement of one substance by another. Morphologically these lichen populations are sometimes indistinguishable, but possess well-defined and constant variations in chemical compositions. For example, Psuedovernia furfuraceae has three chemical races. One race contains olivetoric acid and is found in Northern Europe, a second is found in Southern Europe and Northern Africa and contains physodic acid and third containing lecanoric acid is found i n North America. Biogenetically the first two compounds are closely related because one can be derived from the other by one single step. But lecanoric acid is not biosynthetically related to these two compounds. I t is therefore now recognized t h a t when biogenetic demarcations complement biogeographic variations, such taxa should be recognized as separate species. Thus, the North American taxon is named as P. c o n s ~ c i a n s . ~ However, it has been suggested that the best way to recognize whether chemical variation is under genetic control or not is through sympatric chemical races which maintain their chemical integrity even when growing side by side." Chemosyndrome refers to a group of biosynthetically related lichen substances and the major metabolite is accompanied by several minor constituents which are biosynthetically relatede4 The compounds which are major constituents in one species may occur as minor constituents in related taxa and vice versa. These chemotypes can also be considered as sibling species5, where reproductive isolation of a species often leads to ecological differentiation. This is a common result of evolution in animals and vascular plants. The occurrence of accessory compounds may tend to complicate the above arguments. These compounds are found at certain times in species, usually i n addition to normal constituents, a n d have relation to the morphological or distributional variation. Thus, these compounds a r e not accorded chemotaxonomic significance. Such compounds Veranja Karunaratne et al. often occur as accessory compounds in more than one species and often vary in quantity. The following spot tests and microchemical methods will be used to garner taxonomical details of lichen specimens: (a) Chemical (colour) tests conducted on lichen thalli The common spot test reagents not only indicate where particular compounds are located in sectioned thalli, but may also indicate the chemical nature of the substance. Reagents for thalline spot tests: K = 10 % aqueous KOH solution a. Turns yellow to red with most o-hydroxy aromatic aldehydes b. T u r n s b r i g h t red to deep p u r p l e w i t h anthraquinone pigments C = saturated aqueous Ca(OCl), or common bleach NaOC1 solution a. Turns red with m-dihydroxy phenols, except for those substituted between the hydroxy groups with a -CHO or -CO,H. b. Turns green with dihydroxy dibenzofurans. KC = 10 % aqueous KOH solution followed by saturated aqueous Ca(OCl), or common bleach (NaOC1) solution. a. Turns yellow with usnic acid. b. Turns blue with dihydroxy dibenzofurans. c. Turns red with depsides and depsidones which, undergo rapid hydrolysis to yield mdihydroxy phenolic moiety. PD = 5 % alcoholic p-phenylenediamine solution a. Turns yellow, orange or red with aromatic aldehydes. (b) Microchemical methods by the position relative to the standards. Data are then sorted to find all compounds with the same R,classes. Of these possibilities, those with similar spot characteristics (colour, fluorescence etc.,) are compared chromatographically w i t h t h e unknown. Additional solvent systems are used for compounds that do not separate well in the initial analysis. Two-dimensional TLC methods are used to evaluate structurally similar compounds. Before spraying the dried plates are examined in day light for pigments and for fluorescence or quenching u n d e r s h o r t (254 n m ) a n d long wavelength (365 nm) ultraviolet (UV) light. Subsequently the plates are sprayed with 10 % sulfuric acid and heated a t 110" C on a hot plate for 10 minutes to develop the spots. The following solvent systems are used: Solvent A = Toluene: dioxane: acetic acid (180: 45: 5) Solvent B = Hexane: methyl tert. butyl ether: formic acid (140: 72: 18) Solvent C = Toluene: acetic acid (170: 30) Additional solvents: Solvent E = Cyclohexane: ethyl acetate (75: 25) Solvent G = Toluene: ethyl acetate: formic acid (139: 83: 8) (ii) High performance liquid chromatography (HPLC): Samples are dissolved in methanol and eluted through the appropriate partition column by using a suitable solvent or solvent system u n d e r high pressure. The substances t h a t separate are detected in a UV detector. The retention time R, and peak intensity are recorded by a chart recorder. HPLC is also used to measure either absolute or relative concentrations of lichen compounds. Gradient elution methods are used. The RI value of an unknown peak is calculated as follows: RI = R, of Peak - R, of benzoic acid xlOO R, of Solorinic acid silica gel plates a n d t h r e e solvent systems (designated A, B & C) and two internal controls RI values are very stable during the (atranorin and norstictic acid) are ~ s e d . All ~,~ Rf, ~ lifetime of a column. Substances which are eluted values are compared. An acetone extract of the a t t h e s a m e time often belong to different lichen is spotted on the plate and subsequently substance classes and can be distinguished by eluted in each solvent system. For each solvent their UV spectra. system, a spot is assigned to a R,class determined (i) Standardized TLC methodology: Commercial Lichens: a chemically important biota (iii) Gas Chromatography and Mass Spectrometry: Xanthones, anthraquinones, dibenzofurans, terpenes a n d pulvinic acid which lack thermolabile ester groups can be studied by gas chromatography coupled with mass spectrometry (GCMS). Xanthones in lichens were studied by injecting a lichen extract directly into a mass spe~trometer.~ More recently, the main terpenoid components of t h e lichens of t h e family Pyxinaceae have been studied by GCMS." Ecological role of lichen substances A significant number of lichen substances are not present in non-lichenised fungi. Many genera of lichens also do not produce any of t h e characterized products. Thus the presence of these compounds are not required to maintain the symbiotic state. Lichen substances are found deposited in either the cortex or the medulla as crystals or on the outer surface of hyphae. The commonest cortical compounds are usnic acid and atranorin. Anthraquinones, pulvinic acid derivatives and xanthones also occur in the cortex to a lesser degree. All these compounds with the exception of atranorin are pigmented. The majority of lichen substances a r e deposited i n t h e medulla. Concentrations of lichen substances are variable and typically it is around 1% but values of 5-6 % a r e not uncommon. Once formed, lichen substances are very stable. In lichens, there is no correlation between growth r a t e s a n d concentration of lichen substances, although high concentrations are found at actively growing lobe or branch tips as well as soralia and apothecial margins. There appear to be several roles of lichen substances which determine t h e relative ecological success of individual lichen species and the structure and diversity of natural lichen communities: (a) light-screening compounds, (b) chemical withering compounds, (c) biologically active compounds, (d) anti-herbivore defense compounds and (e) Allergenic compounds:l (a) Light-screening compounds Many cortical lichen substances show variation in concentration along light gradients. Evidence suggests that these compounds act as lightscreens, regulating the light intensity reaching the algal layer in the upper cortex. Usnic acid, a yellow cortical pigment is the most widespread lichen substance found in thousands of lichens. Lichen populations having usnic acid which are exposed to the sun are yellow, while the shade populations are grey-green, indicating a lower concentration of the depside. Cladonia subtenuis and Parmeliopsis ambigua show concentrations of usnic acid varying along concentration gradients.ll Anthraquinones such as parietin ( a reddish cortical pigment) appear to act like usnic acid acting as a light filter. This compound is characteristic of species within Teloschistaceae. These species occur in exposed habitats. In extreme desert environments, species of Caloplaca may be the only saxicolous lichens present, whileXanthoria and Caloplaca dominate highly exposed coastal areas. Pigmented xanthones in the outer cortex of many crustose lichens may also play a role in regulating the light intensity reaching t h e algal layer. These xanthones which are yellow or brown in colour occur in many species of Lecanora, Pertusaria, Melanaria, Lecidea and Buellia in habitats with high light intensities. Unlike fungal xanthones, lichen xanthones contain one or more chlorinated substituents, suggesting that the availability of chloride in the environment may affect the production of these compound^.^ (b) Chemical withering compounds Lichen substances such as phenolic compounds with carbonyl functional groups play an important role in the withering of rock due to their ability to complex metal ions.12 This process leads to soil formation. Low but significant amounts of these substances dissolve in water. Solubility is determined by the nature and the number of polar groups present. The metal complexation i~ advantageous to many crustose and foliose lichens growing in close proximity to rock substratez Since lichen substances commonly occur as crystals on the outer surfaces of hyphae, it puts them in direct contact with the substrate where biochemical withering could aid attachment. 176 Veranja Karunaratne et al. (c) Biologically active compounds The biological activity of lichen compounds very likely gives them a n ecological advantage in colonization of terrestrial areas. Such compounds have been used by man as well. Lichens have been used in medicine by the ancient Chinese and Egyptians.l"14 The medicinal use of lichens is not prevalent at present, particularly since the testing of the atom bomb and the Chernobyl accident in 1986 which have increased the concentrations of radionuclides (137Cs a n d 40K) to a l a r m i n g prop~rtions.~%ccording to recent investigations, lichens possess a variety of biological activities: ( i ) A n t i m i c r o b i a l a c t i v i t y : Although lichen substances are used as antibiotics, their activity is found to be low compared to those isolated from microorganisms. Furthermore, lichen antibiotics are not adequately water soluble for therapeutic use. Depsides, depsidones and usnic acid are active against Gram-positive microorganism^.^^ Sodium salt of usnic acid was used as a drug in Russia.17 Antiseptic creams such as "Usno" and "Evosin" contain usnic acid. Usnic acid acts by uncoupling phosphorylation and is effective because animal cells are far less permeable to usnic acid than the microorganisms. In S r i Lanka, lichens h a d remained completely unexplored (both chemically and taxonomically) until 1996, when a systematic study to harness their biological and medicinal v a l u e was i n i t i a t e d a t t h e D e p a r t m e n t of Chemistry, University of Peradeniya. Roccella montagnei is a fruticose lichen, which is pendulous or shrubby. It is sparsely to moderately branched and has shades of grey, specially lilac grey. (Figure 5).lDifferent Roccella lichen species a r e used to prepare complex mixtures of pigments for the preparation of litmus paper (pH 4.5 red; pH 8.3 blue). Two compounds Erythrin namely, erythrin and methylorsellinate have been isolated from the dichloromethane extract of Roccella montagnei collected from Bamunakotuwa in Kurunegala district. I n g e n e r a l , e r y t h r i n showed lower antifungal activity when compared with benomyl (Benlate- standard funpcide) in the TLC bioassay technique. Apart from somewhat reasonable activity shown a g a i n s t C o l l e t o t r i c h u m gleosporioides, erythrin exhibited moderate activity against four fungi namely, Colletotrichum musae, Cladosporium cladosporioides, Curvularia trifolii, and Monacrosporium ambrosium. These results were corroborated by a spore germination a s s a y a g a i n s t C o l l e t o t r i c h u m m u s a e . The germination was 70.1 %with erythrin at 10 ppm, while for the untreated control germination was 74.2 %. Methylorsellinate on the other hand, showed significantly higher antifungal activity as compared to erythrin (Table 1).Methylorsellenate, when tested against Colletotrichum gleosporioides, Colletotrichum musae, Cladosporium cladosporioides and Curvularia t r i f o l i i , showed g r e a t e r activity t h a n t h e commercial fungicide Benlate. Methylorsellinate showed lesser activity against t h e fungus, Monacrosporium a m b r o s i u m as compared to Benlate. Based on the diameter of inhibition zones, methylorsellinate was significantly more active than erythrin against C. cladosporioides. The antifungal activity of methylorsellinate was also confirmed by a spore germination assay using Colletotrichum musae. The germination percentage with methylorsellinate was 29.9 %while in the control it was 74 %. In order to show that methylorsellinate was not produced as a n artifact in the methanol Methylorsellenate Lichens: a chemically important biota Figure 5: Roccella montagnei lichen growing on the trunk of a coconut tree, Cocos nucifera Table 1: Antifungal activity of erythrin and methylorsellinate Inhibition zone diameter 1 mm Fungus Erythrin(41) Methylorsellinate(42) Benlate Colletotrichum musae 25 50 Colletotrichum gloeosporioides 20 44 25 Cladosporium cladosporioides 10 50 31 Curvularia trifolii 16 55 25 Monacrosporium ambrosium 10 36 48 extract by the methanolysis of erythrin, the lichen Roccella montagnei was extracted into ethyl acetate. Upon examination by TLC and co-TLC it was evident that this extract also contained both erythrin and methylorsellinate, proving that the latter was indeed a natural product isolated from Roccella montagnei and not an artifact. is found in large amounts in the lichen (4.9 % of dry weight) compared with methylorsellinate (0.3 % of dry weight). It is not uncommon that lichen substances are accumulated in amounts exceeding 20 % of the dry weight of the thallus. These compounds are considered to impart a protective role against invading pathogens and herbivores.18 Both erythrin and methylorsellinate are almost identical in functionality and in general, phenolic compounds a r e known to exhibit antifungal properties. In fact the total hydrolysis of erythrin would yield two molecules of orsellinic acid and such the latter could be considered as the biosynthetic precursor of the former. Erythrin Interestingly, in Roccella montagnei the storage of hydrolytically labile erythrin in large amounts (with its relatively low antifungal activity) leads us to reason that the lichen may be converting erythrin to the strongly antifungal methylorsellinate when required. Veranja Karunaratne et aL. Figure 6: Usnea species from Ambewela growing on decaying Acacia decurrens Ambewelamide A, R = (CH2),CH3 Ambewelamide B, R = (CH2),CH3 (ii) Anti tumour and anti mutagenic activity: (-1Usnic acidlg, protolichesterinic and nephrosteranic acids20,physodalic acid20~21, lichen g l u c a n ~and ~ ~lichenin derivatives" have shown anti tumour and anti mutagenic activity. Lichen polysaccharides, the class of compounds to which the latter two belong are particularly interesting due to their anti tumour activity. The mechanism of action of these lichen polysaccharides are not completely known, b u t a p p e a r s to be host mediated: such compounds a r e believed to g e n e r a t e lymphoid cells, p l a s m a cells, macrophages i n t h e vicinity of t h e grafted tumo~r.~~ It was found that extracts of the lichen Usnea sp., collected from Ambewela, exhibited potent in vitro cytotoxicity (Figure 6 ) . Bioassay guided fractionation of the Usnea sp. extracts led to the isolation of ambewelamides A (1)and B (21, whose structures have been elucidated via a combination of single crystal X-ray diffraction and spectroscopic a n a l y s e ~ . ~ ~ ~ ~ ~ Ambewelamides A (1)and B (2)are new members of a family of highly modified diketopiperazines that include fungal metabolites such as a r a n ~ t i n ~ ~ , apoaranotinZ7,emethallicinsZ8,exserohiloneZ9and SCH64847.30There are reports of anti allergenic 28,anti viralz7,EGF receptor binding 30,phytotoxic 29 activity for t h e members of t h e family of diketopiperazines, but the ambewelamides are the first to show potent cytotoxicity. Ambewelamides are the only members of the family having ring Al Lichens: a chemically important biota E epoxides and a diketopiperazine epidisulphide bridge, suggesting that these functionalities might be necessary for cytotoxicity. The ambewelamides are the first examples of the diketopyperazine family isolated from a lichen. (d) Anti-herbivore compounds herbivores. Since these compounds are bright yellow in colour, their cortical position indicates a n example of warning coloration. The medullary position of pulvinic acid derivatives i n t h e Stictaceae and two species ofParmelio. suggest a defensive role against small, nonvisual1y.-oriented invertebrate herbivores.ll In vascular plants, several studies have Both vertebrate and invertebrate herbivores shown that sesqui- and diterpenoids may act as consume lichens. The reasons for the low observed defense restricting h e r b i ~ o r yThe . ~ ~nutrient rich levels of herbivory may be due to low nutrient species of Pseudocyphellaria, Sticta a n d content of lichens, the difficulties in metabolising lichens, and the sparse nature of lichens as a Nephroma show a surprising lack of herbivory indicating that terpenes may play an important resource. Lichens comprise a major source of role in protecting these species with high nitrogen winter food supply of' caribou and reindeer in content. l1 boreal a r e a s i n t h e Northern Hemisphere. Although lichens are a poor source of proteins (2Fumarprotocetraric acid is a b i t t e r 3 %), they are a n easily accessible carbohydrate source when higher plant foliage is not a ~ a i l a b l e . ~ ~depside occurring in many lichens which appear to deter herbivory. For example, neither Caldonia In addition, herbivory on lichens by insects, mites, arbuscula nor C. rangiferina (both contain molluscs are i m p ~ r t a n t .Despite ~ ~ . ~ the ~ ~ lichens ~~ fumarprotocetraric acid) are eaten by reindeer, being a potential food source for herbivores, while they s r e attracted to C. alpestris which has feeding on lichens is not common. In temperate no fumarprotocetraric acid.39Concentrations of evergreen forests, there is little evidence of f'umarprotocetraric acid vary greatly between v e r t e b r a t e or i n v e r t e b r a t e herbivory. T h e younger and older thallus parts in Cladonia, with resistance of individual lichens toherbivory may t h e g r e a t e s t concentration occurring in be due to a variety of factors. Lichens with metabolically active growing tips." Species gelatinous sheaths covering their surface are containing depsides and depsidones such as relatively immune to herbiv01-y~~ as seen in some diffractic acid, physodic and physodalic acids and species of Collemataceae. Thick cortical structure salazinic acid which are bitter are avoided by with heavily agglutinated hyphae may also Caribou in Canada.41 provide some protection from small invertebrate herbivores. Inorganic chelate agents present in P a r i e t i n , a n a n t h r a q u i n o n e lichen many saxicolous crustose lichens are toxic to substance appears to have a defensive role against numerous insects.36 F o u r groups of lichen herbivore^.^^ Lichen species of Teloschistaceae substances that have possible defensive functions contain relative high contents of ~ a r i e t i n . ~ ~ in lichens are pulvinic acid derivatives, terpenes, depsides and depsidones and anthraquinones. (+)-Usnicacid isolated from a n Usnea sp. i n t h e Ambewela a r e a 4 4showed significant Several studies support t h e primary antitermite activity (80 % mortality at 10 mg) defensive role of pulvinic acid derivatives. Lichens against Glyptotermes dilatatus, which is a major containing these compounds have relatively high live-wood tea-termite a t low elevations. Although nitrogen content thus making them potentially a usnic acid is a well-known antimicrobial valuable food source. Yet, feeding on these taxa compound16 t h e r e is only one report of t h e are virtually unknown. Vulpinic acid, the most insecticidal activity of usnic acid against the widespread of these compounds is toxic to both polyphagous herbivorous insect Spodoptera vertebrates and invertebrate^.^^ The cortical l i t t o r ~ l i s . 'Currently, ~ the control methods of G. presence of most of the pulvinic acid derivatives dilatatus are focused on t h e development of is unusual since they show no variation along light r e s i s t a n t clones a n d t h e r e a r e no reported gradients. B u t t h i s histological position is chemicals which can be used in controlling the consistent with their defensive role against 180 Veranja Karunaratne et al. pest, and as such search for such compounds are imp~rtant.~" (e) Allergenic activity Contact d e r m a t i t i s a m o n g forestry a n d horticultural workers has been attributed to the exposure to lichen substances. Among the many lichen substances responsible for such allergenic action a r e usnic acid, evernic acid, fumarprotocetraric acid, stictic acid and atranorin. A t r a n o r i n a n d stictic acid a r e capable of photosensitizing human skin as well as being contact allergen^.^^ Atranorin and other p-orcinol type depsides absorb UV radiation and this may account for their photo toxicity. Chemical ecology of lichens Sequestering of plant chemicals by insects is a well-known phenomenon. Specifically, butterflies of the family Lycaenidae are known to sequester plant chemicals.47Larvae of Monarch butterflies (Nymphalidae) a c c u m u l a t e toxic cardiac glycosides in mill. weeds, which are used by the a d u l t s to d e t e r predators.48 Although t h e sequestration of lichen compounds by the moths of the families Arctiidae and Lithosiinae are known, butterflies are not known to sequester lichen compounds. Many species of invertebrates live on and among lichens, using them for concealment, shelter or food. Over half of the orders of insects have associations with lichens.49 Moths belonging to the family Lepidoptera have the richest association with li~hens.~lThere are many examples of moth names reflecting the resemblance to lichens, such as the "pale lichen moth" (Crambidia pallida) and the "powdered lichen moth" (Bruceiapuluerina). A noctuid moth, Zanclognatha theralis, in Tennessee, USA, was discovered to both feed on Usnea stigosa and look remarkably like the lichen.52 The experiments with polyphagous larvae of the moth Spodoptera littoralis (Noctuidae) demonstrated pronounced acute toxicity and feeding deterrency for frequently occurring lichen compounds such as (+)- and (-)-usnic acids and vulpinic acid a t concentrations comparable to or even below those found in many lichens.53 Hesbacher et a1." have shown that the lichen phenolics such as parietin, atranorin as well as the hydrolytic cleavage product of atranorin (methyl-2, 4-dihydroxy-3, 6-dimethylbenzoate) among other lichen compounds were detected in 11different moth species of the family Arctiidae. COOMe OH Parietin Methyl-2, 4-dihydroxy-3, 6-dimethylbenzoate Atranorin Lichens: a chemically important biota (-)-Usnic acid a-collatolic acid Vulpinic acid The moth larvae of Lithosiinae feed on epiphytic algae of trees or on the algal component of lichens.48A recent survey for the presence of sequestered lichen compounds i n several Lithosiinae species in Australia and Germany, demonstrated t h a t t h e presence of lichen compounds is variable but wide spread in wildcaught imagines.48 Many snails and slugs (class: Gastropoda) feed on lichens. In Israel, two species of snails that eat lichens growing under the surface of lime stone rocks were discovered to be converting rock to soil a t a n amazing rate. The snails pass significant amounts of rock through their digestive tracts in the process of consuming the lichen. In addition, the snails were taking the nitrogen from the lichen and leaving it behind in the new On the other hand, many lichen species contain bitter compounds that may discourage feeding by invertebrate^.^^ Although the ecological role of the sequestered compounds for the herbivores remains unknown, it is thought that they may be utilized for chemical defense against predators or pathogen^.^^ ~ i g & e7: T. nyseus butterfly landing on B. calycinum Veranja Karunaratne et al. Three species of lichen-grazing snails, Baleaperversa, Chondrina clienta and Helicig-ona lapicida were found to sequester lichen compounds when feeding on the crustous lichen species Aspicilia calcarea, Caloplaca atra and Xanthoria parietina. The lichen compounds such as (+)-usnicacid or a-collatolic acid were detected in the feces of these snails, suggesting selective uptake of lichen compounds by the snails.57 Lycaenidae is t h e largest family of butterflies in Sri Lanka and consist of 40 genera, comprising 8 1 species.50Some species of Lycaenidae live on plants, a few live or, scaleinsects, others feed on the larvae and pupae of the large red ant Oecophylla smaragdina, while many are actually tended by ants in their nests. Some are cannibalistic, devouring larvae or the pupae of their own hnd.50Fruit-eating forms are also found in the Lycaenidae. Talicada nyseus Guerin (red Pierrot) which belongs to the family Lycaenidae is distributed from Sri Lanka to Indo-China. The Sri Lankan population belongs to the nominotypical race, which flies through out the island below 4000 feet. The butterflies love human settlement being equally abundant in domestic gardens as it is in the It is plentiful all over the island all the year round and flies slowly near the ground and settles f r e q ~ e n t l y .The ~ ~ natural food source of its larvae is Bryophyllum calycinum (Sinhala: akkapana; Tamil: malai-kalli) (Figure 7). During our studies of Sri Lankan lichens, we discovered the sequestration of lichen products belonging to the crustaceous lichen Leproloma sipmanianum Kurnmerl & Leukert by t h e butterfly T. nyseus Guerin (red pierrot) (Lycaenidae) in Beragala (80" 54' 30" E, 6"45' 30" N), Uva province, Sri Lanka.58T. nyseus was found flying close to the extensive lichen thallus, growing on road side proterozoic rocks of gneiss and quartz schist which are exposed at road-edges, and roosting on it periodically (Figure 8). Pupae of the butterflies were also located anchored to the thallus surface. T. nyseus were collected from Beragala between the period from September 1996 and March 2001. Out of seven butterfly collections made, on three occasions, five compounds namely, zeorin 1, psitosterol 2, a long-chain fatty acid ester 3, atranorin 4, and (+)-usnic acid 5, were present in wild-caught imagines (Figure 9). The butterf ies which emerged from the pupae also contained these compounds. The presence of these compounds in the butterfly was confirmed by comparative TLC studies a n d GC-MS with authentic lichen substances isolated from t h e lichen L. sipmanianum. The butterflies found at the University of Peradeniya and Kandy contained only psitosterol Figure 8: T.nyseus butterfly landing on L. sipmanianum lichen Lichens: a chemically important biota CH3(CH2)4ICH20CCH2CH2CH2CH3 Me H 3 CHO OH 0 0 Figure 9: Structures of lichen compounds present in L. sipmanianum and T. nyseus: 1. Zeorin, 2. BSitosterol, 3. Fatty acid ester, 4. Atranorin, 5. (+)-Usnicacid. 2 (common plant sterol also found in the host plant B. calycinum) with no trace of other lichen compounds. It was concluded that the presence of lichen products of L. sipmanianum in adult T. nyseus butterflies indicated that its larvae may feed on the lichen although the natural food source is B. calycinum. Out of the seven butterfly collections made from Beragala, all five compounds, (1) - (5) were present in adult butterflies on three occasions. Importantly, on two occasions, the butterflies which emerged from laboratory-reared pupae also contained compounds (1) - (5). (+)-Usnicacid (5) was present only on three occasions and atranorin (4) four times. T. nyseus collected during all the field visits contained zeorin (I), 0-sitosterol (2) and the fatty acid ester (3). In order to expand the scope of the study and investigate the presence of lichen products in T. nyseus in other areas of the island, butterflies were collected from four other locations: foot hills of Adam's peak in Central Province (80"30' 50" E, 6" 50' N), Peradeniya University premises (80"35' 30" E, 7" 15' N), from a domestic garden in Aniewatta, Kandy (80"38' E, 7" 17' 30" N), both in the Central Province and in Ukgalkalthota (80" 52' 30" E, 6" 39' 30' N) in the Uva Province. Butterflies were subjected to the same analysis protocol as described above using both TLC and GC-MS. Interestingly, the butterflies found at the University of Peradeniya and Kandy contained only 0-sitosterol with no trace of other compounds. However, those collected in Ukgalkalthota contained atranorin (4) and 0-sitosterol while butterflies a t the foot-hills of Adam's peak contained (+)-usnic acid (5) along with 0sitosterol. Examination of the extracts (hexane, dichloromethane and methanol) of air-dried plant material (140 g) of BryophylLum calycinum by TLC and MPLC did not reveal the presence of lichen compounds (I), (3), (4) and (5). However, the dichloromethane extract (1.50 g) whose TLC showed the presence of p-sitosterol upon MPLC on silica gel (eluent: s t e p gradient from dichloromethane: hexane (2: 3) to methanol: dichloromethane (1:79)) yielded fl-sitosterol as a colourless crystalline solid (0.021 g) confirming that B. calycinum contains p-sitosterol 2 and it could be the source of the butterfly. It may be concluded that the presence of lichen products found in L. sipmanianum in adult Veranja Karunaratne et al. T. nyseus butterfly indicates that its larvae can feed on the lichen although its natural food source is B. c a l y c i n u m . In t h e case of T. n y s e u s individuals contained only P-sitosterol (Peradeniya and Kandy), the larvae probably used the natural food source as their only diet. On the other hand, the presence of atranorin (4) and (+)usnic acid ( 5 ) , compounds which are found only in lichens, in the butterflies in Ukgalkalthota and at the foot-hillsof Adam's peak, respectively, lends credence to the evidence accumulated in Beragala that the larvae of T. nyseus may indeed sequester lichen products. References 1. 2. Huneck S. & Yoshimura I. (1996). Identification of L i c h e n S u b s t a n c e s , S p r i n g e r , B e r l i n Heidelberg, New York. Elix J. A. (1996). Biochemistry and secondary metabolites. In: Lichen Biology (Ed. Nash T. III), pp. 154-180. Cambridge University Press. 3. Culberson C. F. (1969). Chemical and Botanical Guide to Lichen Products. Univ. North Carolina Press, Chapel Hill. 4. Culberson C. F . & Culberson W. L. (1976). Chemosyndromic variation i n lichens. Systematic Botany 1: 325-339. 5. Culberson C. F. (1986). Biogenetic relationships of t h e lichen substances in t h e framework of systematics. Bryologist 89: 123.131. 6. Culberson C. F . (1972b). Improved conditions and new d a t a for t h e identification of lichen products by a standardized thin-layer chromatographic method. Journal of Chromatography 72: 113-125. 7. C u l b e r s o n C . F . & A h m a d j i a n K. (1979). Standardmethode zur Dunnschicht c h r o m a t o g r a p h i von F l e c h t e n s u b s t a n z e n . Herzogia 5: 1-24. 8. Culberson C. F., Culberson W. L. & Johnson A. (1981).A Standardized TLC analysis of b-orcinol depsidones. Bryologist 84: 16-29. 9. Santesson J. (196933). Chemical s t u d i e s on lichens. 10 Mass spectrometry of lichens. Ark Kemi 30: 363-377. A p r o o t A. ( 1 9 8 7 ) . T e r p e n o i d s i n t r o p i c a l Pyxinaceae (Lichenized fungi). XNInternational Botanical Congress Abstracts, Berlin: 504 -507. Rundel P . W. (1978). T h e ecological role of secondary lichen s u b s t a n c e s . Biochemical Systematics and Ecology 6: 157-170. W e s s e l s D . C. & S c h o e m a n n P. ( 1 9 8 8 ) . Mechanisms and rate of whithering of Clarens sandstone by a n endolithic lichen. South African Journal of Science 84: 274-277. Brightman I?. H. (1960). Antibiotics from lichens. Biology and H u m a n Affairs 26: 1-5. S m i t h A. L . ( 1 9 2 0 ) . L i c h e n s . C a m b r i d g e University Press, London. Heinrich G., Gries A., Muller H. & Oswald K. (1989). Ruckblick. Drei J a h r e nach Tschernobyl. Mitt Naturwiss Ver Steiermark 119: 5-13. Ingolfsdottir K., Bloomfield S. F. & Hylands P. J . (1985). I n vitro evaluation of the antimicrobial activity of lichen metabolites a s potential preservatives. Antinzicroial Agents Chemother 28: 289-292. Savicz V. P., Litvinov M. A. & Moissejeva E. M. (1960). E i n Antibiotikum a u s Flechten als Arzneimittel. Planta Medica 8: 192-202. I r w i n M. B. ( 1 9 7 3 ) . T h e L i c h e n s . ( E d s . Ahmadjian V . & Hale M.), pp. 25. Academic Press, New York. Takai M, Uehara, Y. & Beisler J . A. (1979). Usnic acid derivatives a s potential antineoplastic agents. Journal of Medicinal Chemistry 22: 1380-1384. Hirayama T., Fujikawa F., Kasahara T., Otsuka M., Nishida N. & Mizuno D. (1980). Anti-tumor activities of some lichen products a n d their degradation products. Yalzugaku Zasshi 100: 755-759. Shibamoto T. & Wei C. I. (1984). Mutagenicity of l i c h e n c o n s t i t u e n t s . E n v i r o n m e n t a l Mutagenesis 6: 757-762. Hirabayashi K., Iwata S., Ito M., Shigeta S., Narui T., Mori T. & Shibata S. (1989). Inhibitory effect of a lichen polysaccharide sulfate, GE-3- Lichens: a chemically important biota S , on t h e replication o f h u m a n immunodeficiency virus ( H I V ) i n vitro. Chemistry Pharmaceutical Bulletin 37: 2410-2344. Demleitner S., K r a m s J . & Franz G . (1991). S y n t h e s e and Antitumoraktivitat v o n Licheninderivaten. Pharmazie Unserer Zeit 20: 120. Peake J . F., & James, P.W. (1967).Lichens and mollusca. Lichenologist 3: 425. Gerson U . (1973).Lichen arthropod associations. Lichenologist 5: 434-443. Sowter F.A. (1971). Mites (Acari)ans lichens. Lichenologist 5: 176. T o k u z e n R. (1971).Comparison o f local cellular reaction t o t u m o r grafts i n mice treated w i t h some plant polysaccharides. Cancer Research 31: 1590. Grandjean F. (1950).Observations ethologiques sur C a m i s i a s e g n i s ( H e r m . ) e t P l a t y n o t h r i s peltifer ( K o c h ) ( A c a r i e n s ) . BuLletin o f the American Museum of Natural History 22: 224. Williams D. E., Bombuwela K., Lobkovsky E., De Silva E.D., Karunaratne V., Allen T . , Clardy J . & Andersen R. (1998).Ambewalamides A and B , A n t i n e o p l a s t i c Epidithiapiperazinediones Isolated from t h e Lichen Usnea sp. Tetrahedron Letters 39: 9579-9582. S a x e n a P. N . & S a x e n a S . (1989).Insecticidal activity o f some penta- and hexa-coordinated heterocyclic P-diketone derivatives o f diorganotin ( I V ) against t h e stored product pest Rhizopertha (Coleoptera: Bostrichidae). Applied Organometallic Chemistry 8: 279-281. Karunaratne V., Bombuwela K . & De Silva E. D. ( 1 9 9 8 ) . A m b e w a l a m i d e s A and B , Novel Antineoplastic Compounds f r o m S r i L a n k a n Usnea sp. Proceedings of the Award Winning Papers, U n i v e r s i t y of Peradeniya Research Sessions 3 A , 83. Slansky F. (1979).Effect o f t h e lichen chemicals atranorin and vulpinic acid upon feeding and growth o f larvae o f t h e yellow-stripped armyworm, Spodoptera ornithogalli. Enviornmental Entomology 8: 865-868. Neuss N., Nagarajan R., Molloy B. B. & Huckstep L. L. (1968). Aranotin and related metabolites from Arachniotus aureus ( E i d a m )Schroeter. IV. Fermentation, isolation, structure elucidation, biosynthesis. Antimicrobial Agents Chemother 8: 213-219. Kawahara N . , Nozawa K., Y a m a z a k i M., Nakajima S . & Kawai K. (1990). Structures o f novel epipolythiodioxopiperazines, emethallicins B , C , and D , potent inhibitors o f h i s t a m i n e release from Emericella heterothallica. Chemistry Pharmaceutical Bulletin (Tokyo)38: 73-78. Sugawara K., Sugawara F., Strobe1 G . A., Fu. Y . , He C . H . & Clardy J . (1985).Exserohilone, a novel phytotoxin produced b y Exserohilirtm holmii. Journal of Organic Chemistry 50: 5631. Hegde V . R., Pate1 M., Das P.R. & Puar M.S. ( 1 9 9 7 ) . Novel t h i o d i k e t o p i p e r a z i n e f u n g a l metabolites as epidermal growth factor receptor antagonists. Tetrahedron Letters 38: 911. Pulliainen E. (1971). Nutritive value o f some lichens used as food b y reindeer i n northeastern Lapland. Annales Zoologici Fennici 8: 385-389. W r i g h t A . B. ( 1 9 9 3 ) . Peterson first guide to caterpillars o f N o r t h America. Houghton Mifflin Co., Boston. L a w r e y J . D . ( 1 9 8 0 b ) . Correlations b e t w e e n lichen secondary chemistry and grazing activity b y Palliferavaria. Bryologist 83(3): 328-334. Laundon J . R. (1971a). Lichen communities destroyed b y psocids. Lichenologist 5: 177. Ahti T . (1977). Lichens i n t h e boreal coniferous zone. In: Lichen ecology. (Eds. Seaward M. R. D.), pp. 145.181. Academic Press, London. Yom-Tov Y . & Galun M. (1971).Note o f feeding h a b i t s o f t h e d e s e r t snails Sphincterochila b o i s s i e r i C h a r p e n t i e r and T r o c h o i d e a (Xerocrassa) seetzeni Charpentier. T h e Veliger 14: 86. Maass W . S . G . (1975b). Lichen substances. V . M e t h y l a t e d d e r i v a t i v e s o f orsellenic acid, l e c a n o r i c a c i d , a n d g y r o p h o r i c acid f r o m Pseudocyphellaria crocata. Canadian Journal of Botany 53: 1031-1039. Veranja Karunaratne et al. Kathirgamanathar S., Williams D. E., Andersen R. J., BombuwelaK., De SilvaD. & Karunaratne V. (2005). b-orcinol depsidones from the lichen Lrsnea sp. Natural Products Research 19 (7): 695701. lichen compounds against the polyphagous herbivorous i n s e c t S p o d o p t e r a l i t t o r a l i s . Phytochemistry 33: 1389-1394. 54. Hesbacher S., Giez I., Embacher G., Fiedler K., Max W., Trawoger A,, Turk R., Lange 0. L. & Proksch P. (1995). Sequestration of lichen compounds by lichen-feeding members of the Arctiidae (Lepidoptera). Journal of Chemical Ecology 21(12): 2079-2089. 55. Hesbacher S. B., Baur A. Bauer & P. Proksch (1995). Sequestration of lichen compounds by three species of terrestrial snails. Journal of Chemical Ecology 21(2): 233-246. 56. Jones C. G. & Shachak M. (1990). Fertilization of the desert soil by rock-eating snails. Nature 346: 839-841. 57. Lawrey J. D. (1986). Biological role of lichen substances. Bryologist 89(2): 111-122. 58. Orange A., Wolseley P . , Karunaratne V. & Bombuwela K. (2001). Two Leprarioid Lichens New to Sri Lanka. Bibliotheca Lichenologica 78: 327-333. 59. Karunaratne V., Bombuwela K., Kathirgamanathar S., Kumar V., Karuaratne D. N. R a n a w a n a K. B., Wijesundara D. S . A.,Weerasooriya A. & E. D. De. Silva. (2002). An association between the butterfly Talicada nyseus and the lichen Leproloma sipmanianurn as evidenced from chemical studies. Current Science 83 (6): 742-745. 60. Bergwein I. K. (1972). Moss fragrance in modern perfumery. Dragoco Report 3: 43-48. Sivapalan P., Kulasekeran S. & Kathiravetpillai S . (1986). Hand Book on T e a , Tea Research Institute, Sri Lanka. Benzera C., Ducombs G., Sell Y. & Foussereau J. (1985). Plant Contact Dermatitis (Ed. Brian C . Decker), pp. 50-51. Toronto. d' Abrera B. (1998).Butterflies o f Ceylon. pp. 188. Welpac P r i n t i n g & P a c k a g i n g P v t . Ltd., Singapore. Rawlins J . E. (1984). Mycophagy in Lepidoptera. In Fungus-Insect Relationships. (Eds. Wheeler Q . & Blackwell M.), pp. 382-423. Columbia University Press, New York. Richardson D. H. S . (1977). Lichens a n d vertebrates. In: Lichen Ecology. (Ed. Seaward M. R. D) pp. 121-144. Academic Press, London. Woodhouse L. G. 0 . (1952). The butterfly fauna o f Ceylon. (2"d Edition). pp. 57. Ceylon Govt. Press, Colombo. Gerson U. (1950).Lichen Ecology. (Ed. Seaward M . R. D) Academic Press, London 52. Sigal L. L. (1984). Of lichens and lepidopterans. Bryologist 8 7 (1): 66-68. 53. Emmerich R., Giez I., Lange 0. L. & Proksch P. (1993). Toxicity and anti feedant activity of