Earth and Planetary Science Letters 361 (2013) 429–435
Contents lists available at SciVerse ScienceDirect
Earth and Planetary Science Letters
journal homepage: www.elsevier.com/locate/epsl
Constraints on Early Triassic carbon cycle dynamics from paired organic
and inorganic carbon isotope records
K.M. Meyer a,n, M. Yu b, D. Lehrmann c, B. van de Schootbrugge d, J.L. Payne a
a
Department of Geological & Environmental Sciences, Stanford University, 450 Serra Mall, Building 320, Stanford, CA 94305, USA
College of Resource and Environment Engineering, Guizhou University, Caijiaguan, Guiyang 550003, Guizhou Province, PR China
c
Department of Geoscience, Trinity University, One Trinity Place, San Antonio, TX 78212, USA
d
Institute of Geosciences, Johann Wolfgang Goethe University Frankfurt, Altenhöferallee 1, Frankfurt am Main D-60438, Germany
b
a r t i c l e i n f o
abstract
Article history:
Received 5 June 2012
Received in revised form
22 October 2012
Accepted 26 October 2012
Editor: J. Lynch-Stieglitz
Available online 5 December 2012
Large d13C excursions, anomalous carbonate precipitates, low diversity assemblages of small fossils,
and evidence for marine euxinia in uppermost Permian and Lower Triassic strata bear more similarity
to Neoproterozoic carbonates than to the remainders of the Permian and Triassic systems. Middle
Triassic diversification of marine ecosystems coincided with the waning of anoxia and stabilization of
the global carbon cycle, suggesting that environment-ecosystem linkages were important to biological
recovery. However, the Earth system behavior responsible for these large d13C excursions remains
poorly constrained. Here we present a continuous Early Triassic d13Corg record from south China
and use it to test the extent to which Early Triassic excursions in d13Ccarb record changes in the d13C of
marine dissolved inorganic carbon (DIC). Regression analysis demonstrates a significant positive
correlation between d13Corg and d13Ccarb across multiple sections that span a paleoenvironmental
gradient. Such a correlation is incompatible with diagenetic alteration because no likely mechanism
will alter both d13Corg and d13Ccarb records in parallel and therefore strongly indicates a primary
depositional origin. A simple explanation for this correlation is that a substantial portion of the
preserved Corg was derived from the contemporaneous DIC pool, implying that the observed excursions
reflect variation in the d13C of the exogenic carbon reservoir (ocean, atmosphere, biomass). These
findings support existing evidence that large d13C excursions are primary and therefore strengthen the
case that large-scale changes to the carbon cycle were mechanistically linked to the low diversity and
small size of Early Triassic fossils. Associated sedimentary and biogeochemical phenomena further
suggest that similar associations in Neoproterozoic and Cambrian strata may reflect the same
underlying controls.
& 2012 Elsevier B.V. All rights reserved.
Keywords:
Permian–Triassic boundary
extinction
stable carbon isotopes
China
carbon cycle
1. Introduction
Some of the largest d13C excursions of the Phanerozoic are
recorded in Lower Triassic limestones, deposited during the
aftermath of the end-Permian mass extinction (Payne et al.,
2004). These large amplitude ( 48%) positive and negative shifts
are similar in magnitude and timescale to the enigmatic Neoproterozoic and Cambrian carbon isotope perturbations (Halverson
et al., 2005; Maloof et al., 2010). The similarities extend beyond
chemical signatures to characteristic carbonate sediment fabrics,
exceptionally low animal abundance, low taxonomic diversity,
small organism size, and evidence for widespread ocean anoxia
and euxinia (Grotzinger and Knoll, 1995; Maloof et al., 2010;
Meyer et al., 2011; Payne et al., 2004). These observations suggest
n
Corresponding author.
E-mail address: meyerk@stanford.edu (K.M. Meyer).
0012-821X/$ - see front matter & 2012 Elsevier B.V. All rights reserved.
http://dx.doi.org/10.1016/j.epsl.2012.10.035
intimate relationships between carbon cycle behavior, biodiversity, and ecosystem function for both Neoproterozoic–Cambrian
and Early Triassic intervals (Maloof et al., 2005, 2010). However,
the causes of very large and rapid ( o1 My) carbon isotope
excursions in marine limestones remain poorly understood and
several of the proposed explanations are related to diagenesis or
spatial variation in carbonate mineralogy rather than disturbances of the global carbon cycle (Derry, 2010; Swart and
Eberli, 2005). Consequently, constraining the causes of large
variations in the d13C of inorganic carbon in carbonate rocks is
central to understanding the mechanistic links, if any, between
the Early Triassic sedimentary, paleobiological, and biogeochemical records.
Early Triassic excursions in the d13C of carbonate rocks
13
(d Ccarb) occur across the Tethys (Atudorei, 1999; Horacek
et al., 2007; Payne et al., 2004; Richoz, 2004) and in records from
Panthalassa (Horacek et al., 2009) and have therefore been widely
interpreted to result from variation in the d13C of Early Triassic
430
K.M. Meyer et al. / Earth and Planetary Science Letters 361 (2013) 429–435
107° E
Guiyang
26° N
30°N
Yangtze Block
Guanling
Great Bank
of Guizhou
N
Nanpanjiang
Basin
110°E
tze
fo
lat
rm
Zhenfeng
P
Gu
g
n
Ya
izh
Luodian
ou
a
Gu
i
x
ng
S
Nanpanjiang Basin
N
S
M. Triassic
L.Triassic
U. Permian
Bianyang
Guandao
Dawen
250 m
Shelly limestone
Tubiphytes Reef
1 km
Oolitic limestone
Peritidal cyclic limestone
and dolomite
Pelagic limestone
and allodapic breccias
Shale
Stratigraphic section
Thrombolite-bearing
cyclic limestone
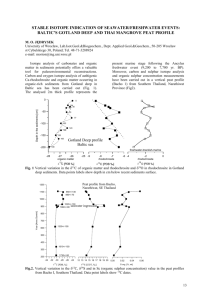
Fig. 1. (A) Paleogeographic map of the Early Triassic GBG (Lehrmann et al., 1998). Inset: The stippled pattern indicates the Nanpanjiang Basin and the brick pattern
represents the Yangtze Block. (B) Schematic cross section of the GBG, modified after Lehrmann et al. (1998). The vertical bars illustrate the locations of the stratigraphic
sections within the GBG.
dissolved inorganic carbon (DIC) in the oceans. Carbon isotope
stabilization early in the Anisian (Middle Triassic) has been
interpreted to reflect either the end of environmental forcing of
carbon cycle perturbations or a stabilization of the carbon cycle
caused by the recovery of global ecosystems (Payne et al., 2004).
Like their Early Triassic counterparts, many Precambrian–
Cambrian isotope excursions can be correlated over vast distances
but the underlying causes remain widely debated. They have been
variously interpreted to reflect perturbations of the global carbon
cycle, unusual configurations or sizes of carbon reservoirs in the
oceans (Grotzinger et al., 2011; Rothman et al., 2003; Tziperman
et al., 2011), or diagenetic overprints (Derry, 2010; Knauth and
Kennedy, 2009). Global processes, such as sea level change,
impact the distribution of carbonate mineralogies and depositional environments and are of particular relevance to the Early
Triassic because the d13Ccarb record observed on widely separated
carbonate platforms could show similar trends in the absence of a
change in the d13C of marine DIC (Swart and Eberli, 2005).
Similarly, global changes in the relative importance of diagenetic
processes such as methanogenesis could also cause an observed
shift in d13Ccarb without a similar change in the d13C of the DIC
reservoir (Hayes and Waldbauer, 2006). Thus, the observation
of globally correlated isotope excursions does not unequivocally
establish that the d13Ccarb variations record secular changes in the
d13C of the marine DIC reservoir.
The carbon isotope composition of organic matter (d13Corg) has
the potential to better constrain the causes of the large Early
Triassic C isotope excursions. Most Phanerozoic paired d13Ccarb
and d13Corg records display the consistent coupling that is
expected when marine organic matter is produced from carbon
in isotopic equilibrium with contemporaneous dissolved inorganic carbon (DIC), especially when variation in d13C of DIC is
greater than any variability in e (fractionation between d13CDIC
and d13Corg during carbon fixation). Any alteration of the d13Corg
record would result from processes different from those known to
diagenetically alter d13Ccarb (Knoll et al., 1986). For example,
thermal maturation of organic matter decreases the total organic
carbon (TOC) composition of rocks and tends to shift residual TOC
to more enriched d13C values (Hayes et al., 1999). This process
would impact the d13Corg values but not the trends (Des Marais
et al., 1992). Migration of hydrocarbons or contamination by
detrital organic Corg from rock weathering could also affect d13Corg
K.M. Meyer et al. / Earth and Planetary Science Letters 361 (2013) 429–435
values. Again, these processes would not impact the d13Ccarb
record, and could be evaluated by examining d13Corg values by
facies or in comparison to TOC. Therefore, paired d13Ccarb and
d13Corg records can reveal the likelihood that perturbations in
either one are primary. If paired d13Ccarb and d13Corg measurements are correlated, the record is likely driven by changes in the
exogenic carbon cycle. If the isotope measurements are not
correlated, the perturbations may be related to diagenetic controls (Derry, 2010; Knauth and Kennedy, 2009) or non-steady
state carbon cycle behavior and the buildup and oxidation of a
large organic carbon reservoir (Rothman et al., 2003; SwansonHysell et al., 2010). If the Early Triassic records are correlated and
the d13Ccarb excursions represent changes in the contemporaneous carbon cycle, then a steady state interpretation of C cycle
dynamics requires extreme shifts in the ratio of organic to
inorganic carbon burial (Maloof et al., 2010; Payne and Kump,
2007), a substantial change in riverine d13Ccarb, or a change in the
fractionation between d13Ccarb and d13Corg.
Here we use a paired carbon isotope approach to evaluate the
extent to which the observed perturbations reflect changes in the
Early Triassic ocean chemistry and to gain insight into the nature
of the carbon cycle during biotic recovery from mass extinction.
2. Field setting and methods
We measured the d13C composition of limestone and associated organic matter from hand samples from the Great Bank of
Guizhou (GBG), an exceptionally exposed carbonate platform in
the Nanpanjiang Basin of south China (Fig. 1). A syncline dissects
the GBG, providing cross-sectional exposure of thick Upper
Permian through Middle Triassic stratigraphic sections along
an inferred paleobathymetric gradient from the shallow-water
(intertidal to tens of meters) platform interior to deep-water
(hundreds of meters) basin margin environments (Lehrmann
et al., 1998). Separate sections are correlated based upon field
stratigraphic relations and conodont and foraminiferan biostratigraphy (Lehrmann et al., 1998; Meyer et al., 2011; Payne et al.,
2004).
Geochemical and stable isotope analyses were performed in
the Stanford University Stable Isotope Biogeochemistry Laboratories. Limestone samples were powdered using a 0.8 mm dental
drill bit. Micrites were selectively sampled, avoiding calcite veins
and fossils. Dolomitized intervals of Laolaicao and Dajiang were
also sampled, but at a lower density, for completeness. Methods
for d13Ccarb analysis are fully described in Meyer et al. (2011). For
d13Corg analysis, hand samples were powdered with a masonry
bit, with special care taken to avoid weathering fronts and veins.
1–3 g of dried, powdered limestone was dissolved in 3 N hydrochloric acid until reaction was complete to remove carbonate
minerals, typically less than 1 h. After centrifugation, the remaining supernatant was removed and each sample was rinsed 3 times
with 18.2 MO water. d13Corg values of the dried, powdered
residues were then determined using a Costech elemental analyzer coupled to a Thermo Finnegan Deltaplus XL isotope ratio mass
spectrometer. Isotopic composition is reported in standard delta
notation relative to the VPDB standard. Analytical precision was
typically better than 0.2% based on replicate measurements of an
internal laboratory standard.
3. Results
The Lower Triassic d13Corg records from Dawen, Guandao, and
Bianyang presented in Fig. 2 display a magnitude of negative and
positive shifts of up to 6–7%, excluding outliers. This pattern
431
follows that of the d13Ccarb record at these sections and is
consistent with the d13Ccarb records across the GBG.
The d13Corg data display much greater sample-to-sample
scatter than the d13Ccarb values and there is no discernable d13Corg
gradient across the platform (Fig. 5a). All three sections display
shifts in the d13Corg record that correspond to shifts in their
respective d13Ccarb records, though the amplitude of d13Corg
variation is damped. As a result, the D13C (d13Ccarb–d13Corg) also
exhibits shifts parallel to those seen in d13Ccarb (Fig. 2). A positive
correlation between organic and inorganic carbon isotopes is
apparent in scatterplots of d13Corg versus d13Ccarb for each section
(Fig. 3). This correlation is shown to be statistically robust
through generalized least squares regression (Table 1), which
shows that the slopes of d13Ccarb and d13Corg are statistically
different from both zero and one, ranging between 0.4 and 0.6.
4. Discussion
The d13Corg data are consistent with several other lines of
evidence indicating that the large d13Ccarb excursions are primary,
including a lack of significant correlation between d13Ccarb and
d18O or trace metals (as shown in Meyer et al. (2011)). The
analysis presented above indicates that the major excursions in
the d13Ccarb record were not significantly altered by meteoric
diagenesis, organic matter remineralization, or changes in
the mixing ratio of sediment sources and minerals across the
GBG. There is also no correlation between TOC and d13Corg,
suggesting that there is no strong diagenetic control of the Corg
record (Fig. 4; see also Johnston et al., 2012). Finally, organic
petrographic observations show that the organic matter in GBG
limestones is primarily amorphous organic matter with a likely
bacterial or marine phytoplanktonic origin. There is no evidence
for substantial contamination by modern organic matter (Fig. S1).
The positive relationship between d13Corg and d13Ccarb arising
from coeval positive and negative shifts, the global nature of the
d13C carb shifts, and geochemical tests of diagenesis suggest that
the inorganic and organic carbon records reflect variation in the
isotopic composition of contemporaneous DIC. The regression
analysis further supports the coupled nature of these measurements but do not offer a mechanism or physical explanation for
the nature of the coupling. Although the d13Corg and d13Ccarb
records are correlated, the slope of the relationship is significantly
less than unity (Table 1), which is the expected value if organic
carbon is formed from contemporaneous DIC and the fractionation associated with photosynthesis is relatively invariant. The
imperfect correlation between d13Corg and d13Ccarb could be
influenced by contamination, by a large DOC pool and consequent
time-averaging of the d13Corg signal, or by the mixing of Corg
sources. Here we discuss the relative support for these various
possibilities.
First, GBG limestones are low in TOC, and some authors have
suggested that low TOC samples may be prone to contamination
by modern organic matter. However, we find no statistically
significant correlation between %TOC and d13Corg (Fig. 4; adjusted
R^2¼0.003, p ¼0.21) or between lithofacies and d13Corg (Fig. S2).
In addition, organic petrographic analyses of selected GBG samples show no evidence for significant modern material in the
sample residues (Fig. S1). These lines of evidence suggest the
correlation less than unity in the paired d13Corg and d13Ccarb data
is not likely due to contamination by modern organic matter.
Variation in d13Corg could alternatively be damped relative to
changes in d13Ccarb if there is a large DOC reservoir due to timeaveraging of the d13Corg signal. Rothman et al. (2003) have
described a mechanism that can explain decoupling of d13Ccarb
and d13Corg through buildup and oxidation of a large DOC
432
K.M. Meyer et al. / Earth and Planetary Science Letters 361 (2013) 429–435
Fig. 2. Organic and inorganic carbon isotopic data from three stratigraphic sections across the GBG, modified after Meyer et al. (2011). The time scale is based on conodont
biostratigraphy from Guandao (Payne et al., 2004), and correlations are suggested based upon lithostratigraphic relationships (red) and carbon isotope trends (blue).
Conodont first occurrences: a. Neogondolella changxingensis, b. Hindeodus parvus, c. Neospathodus dieneri, d. Ns. waageni, e. Ns. Bansoni. (For interpretation of the references
to color in this figure legend, the reader is referred to the web version of this article.)
Fig. 3. Relationship between d13Ccarb and d13Corg for Dawen, Guandao, and Bianyang limestones. The coefficients of determination (R2) for linear regressions through these
data are 0.43, 0.30, and 0.53, respectively. Full regression statistics are provided in Table 1.
Table 1
Generalized least squares regression shows that the slopes of d13Ccarb and d18Corg
are statistically different from both 0 and 1 and range between 0.4 and 0.6.
Section
Intercept
(e)
Standard
error
Slope
Standard
error
p
Bianyang
Dawen
Guandao
Upper Guandao
28.6
28.2
27.8
28.3
0.22
0.15
0.30
0.68
0.52
0.41
0.48
0.69
0.09
0.05
0.10
0.29
o0.0001
o0.0001
o0.0001
0.04
reservoir. This non-steady state approach to C cycle dynamics has
been used to interpret the Neoproterozoic Shuram excursion,
where a large carbon isotope excursion observed in d13Ccarb is
absent in d13Corg (Fike et al., 2006). Oxidation of a very large
marine DOC pool relative to DIC would cause a negative isotope
excursion in d13Ccarb without changing d13Corg. Based on visual
inspection of values calculated using Rothman et al.’s (2003)
equations, we find that the DOC:DIC ratio would have to be at
least 4 to fit the slope we observe. In addition, the relationship
between d13Ccarb and e (difference between d13Ccarb and d13Corg)
is nearly indistinguishable from the relationship shown by
Rothman et al. (2003) for 731–590 Ma. Although the extreme
scenario for the Shuram has not been proposed for the Permian–
Triassic, non-linear carbon cycle dynamics and the oxidation of
marine organic matter have been implicated in the negative
carbon isotope excursion at the P–T boundary (Rothman, 2010).
Others have suggested that a vigorous biological pump (Meyer
et al., 2011) and DOC oxidation coupled to sulfate reduction may
have impacted C and S cycling during this interval (Luo et al.,
K.M. Meyer et al. / Earth and Planetary Science Letters 361 (2013) 429–435
2010). The key question that remains for this scenario is the
uncertain timescale over which DOC buildup would have been
possible.
0.00
0.05
0.10
0.15
TOC
0.20
0.25
0.30
Alternatively, if the Corg record was originally derived from the
DIC pool, then the observed decoupling (slopes less than one in
Table 1) between the two isotope records could reflect either
covariation of photosynthetic fractionation with the d13C of DIC or
the mixing of two pools of organic carbon, one of which was
tracking the d13C of DIC with a constant offset due to photosynthetic
fractionation and one of which was invariant or varying independently of the d13C of DIC. The former possibility is consistent with
the fact that the photosynthetic fractionation varies over time due to
changes in pCO2, growth rate, cell geometry, carbon concentrating
mechanism, and other factors (reviewed by Hayes et al. (1999)).
Given the many degrees of freedom, it is difficult to either rule out
this possibility or to demonstrate clearly that it is the most likely
explanation for the observed isotopic variations. The latter possibility can be quantified and evaluated more easily, as discussed below.
Variation in the d13Corg signal may be damped relative to variation
in the d13Ccarb signal by the addition of another pool of organic carbon
with a temporally fixed d13C composition, such as recycled ancient
organic carbon (weathered TOC), marine DOC (when this reservoir is
large relative to DIC), or modern terrestrial organic matter. Recently,
Johnston et al. (2012) proposed the incorporation of recycled Corg into
the d13Corg signal as an explanation for intervals where the Neoproterozoic d13Corg and d13Corg records become uncorrelated (because
the d13Corg record shows little or no stratigraphically coherent
variation). To quantify the conditions under which this scenario
would account for our data, we performed a second regression
analysis to estimate the isotope composition and proportional contribution of a second carbon pool to the measured d13Corg record.
0.35
-21
-23
-25
-27
-29
-31
-33
-35
Dajiang
Daw en
Cordev.
Late
Triassic
Carnian
Fig. 4. Relationship between d18Corg and TOC for Dawen, Guandao, and Bianyang
sections (R2 ¼ 0.09).
236
433
Laolaicao
Daw en
Guandao
Bianyang
Guandao
Bianyang
Ladinian
238
240
Long.
242
Middle
Triassic
244
246
Pelsonian
Anisian
Illyrian
A./B.
250
Early
Triassic
Spathian
248
Olenekian
Age (millions of years before present)
Fass.
Smith.
Ind.
252
Late
Permian
Dien.
Gries.
Late
Changsingian
-4
-2
0
C
2
4
6
(‰ VPDB)
8
-4
-2
0
C
2
4
6
(‰ VPDB)
8
-36 -34 -32 -30 -28 -26 -24
C
(‰ VPDB)
0
5
10
15
20
Observed diversity persample
0
1
20
30
40
50
60
Skeletal abundance as vol %
Fig. 5. Summary of Triassic C isotope and paleobiological trends. (A) The Early and Middle Triassic d13C curve from all GBG sections. (B and C) The Early and Middle Triassic
d13Corg and d13Ccarb gradients across the GBG. Plotted d13C values are from correlated samples across the slope (filled symbols) and platform interior sections (open
symbols) as in Meyer et al. (2011). The d13Ccarb gradient is about 4% in the Early Triassic. It collapses to approximately zero in the Middle Triassic interval of accelerated
biotic recovery and carbon isotope stabilization. In contrast, there is no coherent gradient in d13Corg observed across the GBG during the Early Triassic. (D and E) Box plots
of diversity and fossil abundance observed on the GBG from the Late Permian into the Late Triassic. Data from Payne et al. (2006).
434
K.M. Meyer et al. / Earth and Planetary Science Letters 361 (2013) 429–435
Assuming a constant 30% fractionation during photosynthesis, the
equation to describe the proportional contribution of two organic
carbon pools to the measured d13Corg values is:
d13 Corg ¼ ðd13 Ccarb 30Þ f contemp þ ð1f contemp Þ d13 Cother
where fcontemp represents the fraction of Corg derived from the
contemporaneous marine Corg pool and d13Cother represents the
isotope composition of carbon derived from a second pool (weathered
organic carbon, terrestrial organic carbon, modern contamination,
etc.).
Based on this model for the paired d13Corg and d13Ccarb data
across the GBG, regression analysis suggests that 45% of the
Corg measured was derived from the contemporaneous DIC pool
(for Bianyang fcontemp ¼0.52, std error¼ 0.10; for Dawen,
fcontemp ¼0.41, std error¼0.17; for Guandao, fcontemp ¼0.48, std
error¼0.10). This analysis implies that over half of the Corg must
have been from another source with a stochastically constant
composition. Such alternative sources of Corg include weathered
carbon, terrestrial inputs from a variety of sources, and a large
DOC pool with relatively invariant d13C. The predicted d13C composition of this non-DIC derived pool from regression modeling is
27%. Unfortunately, the isotope composition of the
second carbon pool does not allow us to distinguish between these
sources. Regardless of the source of this other organic carbon pool,
however, the significant correlation between the d13Corg and
d13Ccarb records supports a causal connection between carbon
isotope stabilization and enhanced biotic recovery in Middle
Triassic time; carbon isotope stabilization is associated with
increases in fossil diversity and abundance across the platform
(Fig. 5).
At present, we favor the scenario in which the bulk organic
carbon isotope record reflects the mixing of carbon from two
different sources. The extremely low TOC of our samples means
that even a small amount of organic carbon from a source other
than contemporaneous marine organisms could have a substantial impact on the bulk d13Corg record. The larger sample-tosample variation in d13Corg versus d13Ccarb is also consistent with
variation among samples in the proportion of contemporaneous
versus other organic carbon. We cannot rule out the possibility
that temporal variation in the photosynthetic fractionation of
carbon isotopes also contributed to the reduced variation in
d13Corg, but under this scenario our data require that the factors
influencing the photosynthetic fractionation covaried with
d13Ccarb with a slope of approximately one half.
5. Conclusions and implications
Here we present the first paired d13Ccarb and d13Corg record for the
latest Permian and Early Triassic. Large positive and negative excursions occur in both records, but the magnitude of the shift is slightly
damped in d13Corg. Regression modeling demonstrates a significant
correlation between d13Corg and d13Ccarb, requiring that a significant
fraction of the preserved Corg formed from the contemporaneous DIC
pool. This result therefore implies that the observed excursions reflect
variation in the d13C of the exogenic carbon reservoir. In addition, this
evidence further implies that Early and Middle Triassic changes in
carbon cycle behavior, carbonate deposition, and marine ecosystem
recovery share underlying controls. Parallels between these findings
and recent results from Neoproterozoic strata also suggest that a
damped response in d13Corg relative to d13Ccarb may be typical of
low TOC rocks across high-amplitude carbon isotope excursions of
all ages.
Acknowledgments
The authors thank E. Schaal, X. Li, and H. Fu for assistance in the
field. P. Blisnuik provided laboratory assistance. Meyer acknowledges
support from the Agouron Institute. This work was supported by NSF
grant EAR-0807377-007, ACS PRF grant 45329-G8, and NGS grant
8102-06.
Appendix A. Supplementary materials
Supplementary data associated with this article can be found in
the online version at http://dx.doi.org/10.1016/j.epsl.2012.10.035.
References
Atudorei, V., 1999. Constraints on the Upper Permian to Upper Triassic Marine Carbon
Isotope Curve: Case Studies from the Tethys. Ph.D., University of Lausanne.
Derry, L.A., 2010. A burial diagenesis origin for the Ediacaran Shuram–Wonoka
carbon isotope anomaly. Earth Planet. Sci. Lett. 294, 152–162.
Des Marais, D.J., Strauss, H., Summons, R.E., Hayes, J.M., 1992. Carbon isotope evidence
for the stepwise oxidation of the Proterozoic environment. Nature 359, 605–609.
Fike, D.A., Grotzinger, J.P., Pratt, L.M., Summons, R.E., 2006. Oxidation of the
Ediacaran Ocean. Nature 444, 744–747.
Grotzinger, J.P., Fike, D.A., Fischer, W.W., 2011. Enigmatic origin of the largestknown carbon isotope excursion in Earth’s history. Nat. Geosci. 4, 285–292.
Grotzinger, J.P., Knoll, A.H., 1995. Anomalous carbonate precipitates: is the
Precambrian the key to the Permian? Palaios 10, 578–596.
Halverson, G.P., Hoffman, P.F., Schrag, D.P., Maloof, A.C., Rice, A.H.N., 2005. Toward
a Neoproterozoic composite carbon-isotope record. Geol. Soc. Am. Bull. 117,
1181–1207.
Hayes, J.M., Strauss, H., Kaufman, A.J., 1999. The abundance of 13C in marine
organic matter and isotopic fractionation in the global biogoechemical cycle of
carbon during the past 800 Ma. Chem. Geol. 161, 103–125.
Hayes, J.M., Waldbauer, J.R., 2006. The carbon cycle and associated redox processes
through time. Philos. Trans. R. Soc. B-Biol. Sci. 361, 931–950.
Horacek, M., Brandner, R., Abart, R., 2007. Carbon isotope record of the P/T boundary
and the Lower Triassic in the Southern Alps: evidence for rapid changes in storage
of organic carbon. Palaeogeogr. Palaeoclimatol. Palaeoecol. 252, 347–354.
Horacek, M., Koike, T., Richoz, S., 2009. Lower Triassic d13C isotope curve from
shallow-marine carbonates in Japan, Panthalassa realm: confirmation of the
Tethys d13C curve. J. Asian Earth Sci. 36, 481–490.
Johnston, D.T., Macdonald, F.A., Gill, B.C., Hoffman, P.F., Schrag, D.P., 2012.
Uncovering the Neoproterozoic carbon cycle. Nature 483, 320-U110.
Knauth, L.P., Kennedy, M.J., 2009. The late Precambrian greening of the Earth.
Nature 460, 728–732.
Knoll, A.H., Hayes, J.M., Kaufman, A.J., Swett, K., Lambert, I.B., 1986. Secular
variation in carbon isotope ratios from Upper Proterozoic successions of
Svalbard and East Greenland. Nature 321, 832–838.
Lehrmann, D.J., Wei, J.Y., Enos, P., 1998. Controls on facies architecture of a large
Triassic carbonate platform: the Great Bank of Guizhou, Nanpanjiang Basin,
South China. J. Sediment. Res. 68, 311–326.
Luo, G.M., Kump, L.R., Wang, Y.B., Tong, J.N., Arthur, M.A., Yang, H., Huang, J.H., Yin,
H.F., Xie, S.C., 2010. Isotopic evidence for an anomalously low oceanic sulfate
concentration following end-Permian mass extinction. Earth Planet. Sci. Lett.
300, 101–111.
Maloof, A.C., Porter, S.M., Moore, J.L., Dudas, F.O., Bowring, S.A., Higgins, J.A., Fike,
D.A., Eddy, M.P., 2010. The earliest Cambrian record of animals and ocean
geochemical change. Geol. Soc. Am. Bull. 122, 1731–1774.
Maloof, A.C., Schrag, D.P., Crowley, J.L., Bowring, S.A., 2005. An expanded record
of Early Cambrian carbon cycling from the Anti-Atlas Margin, Morocco. Can.
J. Earth Sci. 42, 2195–2216.
Meyer, K.M., Yu, M., Jost, A.B., Kelley, B.M., Payne, J.L., 2011. d13C evidence that
high primary productivity delayed recovery from end-Permian mass extinction. Earth Planet. Sci. Lett. 302, 378–384.
Payne, J.L., Kump, L.R., 2007. Evidence for recurrent Early Triassic massive
volcanism from quantitative interpretation of carbon isotope fluctuations.
Earth Planet. Sci. Lett. 256, 264–277.
Payne, J.L., Lehrmann, D.J., Wei, J.Y., Orchard, M.J., Schrag, D.P., Knoll, A.H., 2004.
Large perturbations of the carbon cycle during recovery from the end-Permian
extinction. Science 305, 506–509.
Payne, J.L., Lehrmann, D.J., Wei, J.Y., Knoll, A.H., 2006. The pattern and timing of
biotic recovery from the end-Permian extinction on the Great Bank of
Guizhou, Guizhou province, China. Palaios 21, 63–85.
Richoz, S., 2004. Stratigraphy and Carbon isotope study of the Upper Permian and
Lower Triassic in Some Localities of the Neotethyan Realm (Turkey, Oman,
Iran), University of Lausanne.
Rothman, D.H., 2010. Singular blow-up in the end-Permian carbon cycle, 2010 Fall
Meeting. AGU, San Francisco, CA.
K.M. Meyer et al. / Earth and Planetary Science Letters 361 (2013) 429–435
Rothman, D.H., Hayes, J.M., Summons, R.E., 2003. Dynamics of the Neoproterozoic
carbon cycle. Proc. Natl. Acad. Sci. USA 100, 8124–8129.
Swanson-Hysell, N.L., Rose, C.V., Calmet, C.C., Halverson, G.P., Hurtgen, M.T.,
Maloof, A.C., 2010. Cryogenian glaciation and the onset of carbon-isotope
decoupling. Science 328, 608–611.
Swart, P.K., Eberli, G., 2005. The nature of the d13C of periplatform sediments:
implications for stratigraphy and the global carbon cycle. Sediment. Geol. 175,
115–129.
435
Tziperman, E., Halevy, I., Johnston, D.T., Knoll, A.H., Schrag, D.P., 2011. Biologically
induced initiation of Neoproterozoic snowball-Earth events. Proc. Natl. Acad.
Sci. USA 108, 15091–15096.