P1: GDL
April 12, 2001
9:48
Annual Reviews
AR129-12
Annu. Rev. Plant Physiol. Plant Mol. Biol. 2001. 52:315–33
c 2001 by Annual Reviews. All rights reserved
Copyright °
THE PLASTID DIVISION MACHINE
Katherine W Osteryoung and Rosemary S McAndrew
Department of Plant Biology, Michigan State University, East Lansing, Michigan 48824;
e-mail: osteryou@msu.edu, smithr72@msu.edu
Key Words chloroplast, FtsZ, MinD, bacterial cell division, organelle fission,
Arabidopsis
■ Abstract Plastid division is essential for the maintenance of plastid populations
in cells undergoing division and for the accumulation of large chloroplast numbers in
photosynthetic tissues. Although the mechanisms mediating plastid division are poorly
understood, ultrastructural studies imply this process is accomplished by a dynamic
macromolecular machine organized into ring structures at the plastid midpoint. A key
component of the engine that powers this machine is the motor-like protein FtsZ, a
cytoskeletal GTPase of endosymbiotic origin that forms a ring at the plastid division
site, similar to the function of its prokaryotic relatives in bacterial cytokinesis. This
review considers the phylogenetic distribution and structural properties of two recently
identified plant FtsZ protein families in the context of their distinct roles in plastid
division and describes current evidence regarding factors that govern their placement
at the division site. Because of their evolutionary and mechanistic relationship, the
process of bacterial cell division provides a valuable, though incomplete, paradigm for
understanding plastid division in plants.
CONTENTS
INTRODUCTION . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
MORPHOLOGICAL ASPECTS OF PLASTID DIVISION . . . . . . . . . . . . . . . . . .
EVOLUTIONARY ORIGIN OF THE DIVISION APPARATUS . . . . . . . . . . . . . . .
FTSZ FUNCTION IN PLASTID DIVISION . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
Bacterial FtsZ . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
Phylogenetic Distribution of FtsZ Genes . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
Evolutionary and Functional Diversity Among Plant FtsZ Proteins . . . . . . . . . . . .
Proposed Functions of FtsZ1 and FtsZ2 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
COMPARISON BETWEEN PLANT FTSZ1 AND FTSZ2 PROTEINS . . . . . . . . . .
Structural Domains . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
Posttranslational Modification Motifs . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
PLACEMENT OF THE PLASTID DIVISION MACHINERY . . . . . . . . . . . . . . . .
Selection of the Cell Division Site in Bacteria . . . . . . . . . . . . . . . . . . . . . . . . . . .
Role of MinD in Plastid Division Site Selection . . . . . . . . . . . . . . . . . . . . . . . . .
ADDITIONAL COMPONENTS OF THE PLASTID DIVISION APPARATUS . . . .
FUTURE CHALLENGES . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
1040-2519/01/0601-0315$14.00
316
316
317
318
318
318
318
321
322
322
324
324
324
325
326
326
315
P1: GDL
April 12, 2001
316
9:48
Annual Reviews
OSTERYOUNG
¥
AR129-12
MCANDREW
INTRODUCTION
It is perhaps premature given the sparse state of our knowledge to refer to the
protein components that make up the plastid division complex as a “machine.”
Nevertheless, plastid division, like other cellular processes involving remodeling
of organelles, is clearly orchestrated by a macromolecular complex composed of
numerous proteins that coordinate the mechanical activity required to constrict
the plastid. This review focuses specifically on the molecules that are known or
postulated to make up the plastid division apparatus in photosynthetic eukaryotes.
Because only a few components of the divison complex have been identified to
date, their functions are emphasized. However, information from other systems,
most prominently bacteria, is included in the context of its potential relevance
to understanding plastid division. For literature related to the developmental and
regulatory aspects of plastid division, the reader is referred to other reviews (9,
48–50, 85a–88).
MORPHOLOGICAL ASPECTS OF PLASTID DIVISION
A brief discussion of the morphological and ultrastructural changes that accompany progression through a plastid division cycle is in order at the outset because
functional analysis of the protein participants must be interpreted within the context
of what is actually observed. More detailed treatments of this subject can be found
in the following references: (46, 47, 51, 86, 121). Most of the information derived
from light microscopy has concentrated on observations of green chloroplasts because they are easily viewed, but ultrastructural evidence and recent molecular and
genetic data suggest that the division process and associated machinery are likely
to be similar for all plastid types.
At the level of light microscopy, the first visible indication that chloroplast
division has commenced is the formation of a constriction located at the plastid
midpoint, perpendicular to the longitudinal axis (48, 51). In land plants, division is
accompanied by a lengthening and narrowing of the chloroplast, and at late stages
a narrow, twisted isthmus joining the two daughter plastids is sometimes observed.
At the ultrastructural level, formation of the central constriction is frequently associated with the appearance of an electron-dense annular structure termed the plastid
dividing (PD) ring (11–13, 23, 38, 39, 45, 47, 51, 64, 67, 68, 79, 80, 86, 114). Such
structures have been described in chloroplasts from diverse plant and algal taxa, as
well as in amyloplasts and proplastids from various tissues (tabulated in 47). In the
red algae Cyanidioschyzon merolae and Cyanidium caldarium, in which the ultrastructure of plastid division has been investigated in detail, the PD ring is visible
early in the division process and becomes thicker as the constriction narrows, but
in land plants the PD ring does not become visible until constriction is well under
way. In many electron micrographs, the PD ring can be resolved into two concentric
rings, one associated with the stromal surface of the inner envelope membrane and
P1: GDL
April 12, 2001
9:48
Annual Reviews
AR129-12
PLASTID DIVISION
317
the other associated with the cytosolic surface of the outer envelope membrane.
More recent work has revealed a third electron-dense component of the PD ring
localized in the intermembrane space in C. merolae (66). In some studies, thin
filaments encircling the constriction have also been described (13, 47, 79, 116). It
is generally assumed that the PD ring functions in the division process by constricting the membranes, but the molecular composition of these rings and how
they function remains to be established.
EVOLUTIONARY ORIGIN OF THE DIVISION APPARATUS
Like many plastid-associated processes, plastid division has its evolutionary origin in the cyanobacterial endosymbiont that gave rise to chloroplasts (82). The
possibility that the plastid division apparatus might have components in common
with those involved in prokaryotic cell division was recognized by Possingham &
Lawrence (86) and more recently by Suzuki et al (114). The latter investigators
specifically suggested a possible relationship between the PD rings in C. merolae
and the bacterial cell division protein FtsZ, an essential cytoskeletal component of
the bacterial cell division apparatus that forms a contractile ring during cytokinesis
(described below). This relationship was confirmed with the discovery of a nuclear
gene from Arabidopsis thaliana that encoded a protein with over 40% amino acid
identity to many bacterial FtsZ proteins and over 50% identity to the cyanobacterial
FtsZs. The further demonstration that the Arabidopsis FtsZ protein was synthesized as a precursor and posttranslationally targeted to the chloroplast by virtue
of a cleavable chloroplast transit peptide strongly suggested its involvement in
chloroplast division (84).
A definitive role for plant FtsZ proteins in plastid division was subsequently
confirmed in the moss Physcomitrella patens and in Arabidopsis. In the moss,
homologous recombination was used to create a knockout mutation in one of two
FtsZ genes present in this organism (111). Instead of having multiple chloroplasts
as in wild-type plants, cells in the deletion mutants appeared to contain only a
single “macrochloroplast” in every tissue examined, consistent with a defect in
plastid division. In Arabidopsis, an antisense approach was employed to generate
transgenic plants in which accumulation of AtFtsZ1-1 (formerly called cpFtsZ), the
chloroplast-targeted FtsZ protein described above, was greatly reduced (83, 110).
In this case as well, cells from mesophyll tissue, instead of containing the wildtype complement of approximately 100 chloroplasts (Figure 1A), contained as
few as one very large chloroplast (Figure 1B). These studies demonstrated an
essential role for FtsZ genes in plastid division in both nonvascular and vascular
land plants, indicating that a portion of the prokaryotic cell division apparatus was
recruited during the evolution of chloroplasts from their cyanobacterial ancestors to
function in plastid division. Subsequent studies described later in this review have
revealed that this conservation extends to other prokaryotic cell division genes as
well.
P1: GDL
April 12, 2001
318
9:48
Annual Reviews
OSTERYOUNG
¥
AR129-12
MCANDREW
FTSZ FUNCTION IN PLASTID DIVISION
Bacterial FtsZ
A brief overview of the structure and function of bacterial FtsZ is important for
understanding how its eukaryotic counterparts might function in plastid division.
The reader is also referred to other recent reviews (7, 25, 58, 62, 76, 104). FtsZ,
like numerous other genes involved in bacterial cell division, was identified genetically in a screen for temperature-sensitive mutants of Escherichia coli that
formed long filaments at the restrictive temperature due to defects in cell division,
and hence they were termed fts mutants (filamentation temperature-sensitive)
(59). The protein is encoded by an ancient and highly conserved gene found in
most prokaryotes, usually in a single copy. FtsZ is now known to be a structural
homologue and very likely the evolutionary precursor of the eukaryotic tubulins
(26, 30, 54, 77). Like tubulin, FtsZ is a self-polymerizing, filament-forming GTPase (8, 15, 28, 72–74, 99, 101, 123), and it functions during bacterial cell division
by assembling into a ring structure at the division site on the interior surface of the
cytoplasmic membrane (5). Mutant analysis has shown that FtsZ ring assembly is
required for the subsequent midcell localization of all other components of the cell
division apparatus (1, 62). The FtsZ ring remains associated with the leading edge
of the division septum throughout cytokinesis, then it disassembles immediately
following cell separation before rapidly reassembling at the center of the newly
formed daughter cells (1, 5, 19, 113). The in vivo structure of the FtsZ ring and how
it effects bacterial cytokinesis are not yet known, but recent data suggest that GTP
hydrolysis triggered by FtsZ polymerization may generate the force and induce
the curvature necessary for constriction (55).
Phylogenetic Distribution of FtsZ Genes
Genes encoding FtsZ have been identified in nearly all prokaryotes, including
archaeal species, cyanobacteria, and the wall-less mycoplasmas as well as in diverse eukaryotes, including unicellular algae and other protists, moss, and vascular
plants (reviewed recently in 2, 33). Most of the eukaryotic FtsZ genes identified
to date are related to those in cyanobacteria and are presumed to be involved in
chloroplast division. Recently, a role for FtsZs in mitochondrial division has also
been uncovered in the chromophyte algae and in red algae, which, in addition to
their cyanobacterial-like FtsZs, contain FtsZ genes closely related to those of the
α-proteobacterial ancestors of mitochondria (3, 115). However, FtsZ genes have
not been found in fungi or animals, organisms in which mitochondrial division is
apparently accomplished by another self-assembling GTPase, dynamin (27, 81).
Evolutionary and Functional Diversity Among
Plant FtsZ Proteins
A significant number of FtsZ sequences from land plants have been submitted to
the gene databases in the past few years. In phylogenetic analyses, these proteins,
P1: GDL
April 12, 2001
9:48
Annual Reviews
AR129-12
PLASTID DIVISION
319
Figure 2 Phylogenetic analysis of FtsZ proteins from land plants performed using Clustal
W (117) at the Biology WorkBench 3.2 website (http://workbench.sdsc.edu). The tree was
rooted with a cyanobacterial sequence (accession P45482; not shown). Accession numbers
are shown in parentheses. Asterisks indicate partial sequences.
all encoded in the nucleus, fall into two major groups that have been designated
the FtsZ1 and FtsZ2 families (83) (Figure 2). Expressed sequence tag collections
suggest that both families are represented in most angiosperms and are encoded by
small gene families. Arabidopsis has one FtsZ1 and two FtsZ2 genes; but multiple
FtsZ1 genes are present in tobacco, and a partial sequence from a fern groups with
the FtsZ1 family (note Neottopteris in Figure 2), suggesting that FtsZ1 and FtsZ2
genes are present in all vascular plants. The only two FtsZ sequences currently
available from a nonvascular plant are from Physcomitrella, and both of these fall
into the FtsZ2 family.
The FtsZ1 and FtsZ2 gene products are more closely related to the cyanobacterial FtsZs than to those in other prokaryotes, consistent with an endosymbiotic
origin in both cases, yet they differ in their overall sequence relatedness and in
their predicted subcellular localizations. All members of the FtsZ1 family for which
full-length sequences are available are predicted by the program TargetP (24) to
contain cleavable chloroplast transit peptides at their amino-terminal ends that
target them to the stromal compartment (Table 1). These predictions have been
verified in in vitro chloroplast import assays for Arabidopsis AtFtsZ1-1 (84) and
for an FtsZ1 family member from Pisum sativum (32). In contrast, most members of the FtsZ2 family are not predicted by TargetP to bear chloroplast transit
peptides, at least not in flowering plants. Consistent with the low TargetP score
P1: GDL
April 12, 2001
320
9:48
Annual Reviews
OSTERYOUNG
¥
AR129-12
MCANDREW
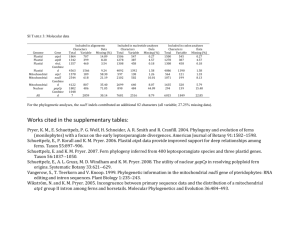
TABLE 1 TargetP predictions for the presence of an N-terminal chloroplast transit peptide for
all full-length FtsZ sequences from land plants currently represented in the public databases
Organism
Accession
number
Family
grouping
TargetPa
scoreb
Chloroplast
import assay
results
Reference
number
Arabidopsis thaliana
Q43545
FtsZ1
.974
Imported
83, 84
—
Nicotiana tabacum
AJ133453
FtsZ1
.934
NPDc
Nicotiana tabacum
AJ271749
FtsZ1
.916
NPD
—
Nicotiana tabacum
AF205858
FtsZ1
.968
NPD
—
Pisum sativum
Y15383
FtsZ1
.945
Imported
32
Tagetes erecta
AF2513460
FtsZ1
.938
NPD
69
Arabidopsis thaliana
AAD21440
FtsZ2
.161d
Not importedd
83
Arabidopsis thaliana
CAB89236
FtsZ2
.374
NPD
83
Gentiana lutea
AAF23771
FtsZ2
.555
NPD
—
Nicotiana tabacum
AJ271750
FtsZ2
.131
NPD
—
Lilium longiflorum
AB042101
FtsZ2
.712
NPD
—
Physcomitrella patens
AJ249140
FtsZ2
.782
Imported
43a
Physcomitrella patens
AJ249138
FtsZ2
.871
Imported
43a, 111
a
(24); http://www.cbs.dtu.dk/services/TargetP/
b
c
Calculated using N-terminal 130 amino acids of predicted open reading frame.
No published data.
d
Based on predicted open reading frame.
of 0.161 calculated for AtFsZ2-1, an Arabidopsis FtsZ2 family member (Table 1,
accession number AAD21440), this protein failed to undergo import into isolated
chloroplasts under the same conditions in which AtFtsZ1-1 was imported (83).
However, the higher TargetP score for an FtsZ2 protein from Lilium longiflorum
suggest that some higher plant FtsZ2 family members could be imported into
the chloroplast, as recently noted for the two FtsZ proteins from Physcomitrella
(43a) (Table 1). This ambiguity underscores the limitations of the available computational resources for predicting the presence or absence of chloroplast transit
peptides, and it emphasizes the need for verifying the localization predictions
experimentally, particularly in cases where the scores are equivocal. A further
caveat to use of TargetP and related programs is that the input sequences used in
the analyses are often taken from the open reading frames (ORFs) identified by
gene prediction programs, which may not accurately predict the true translational
initiation sites. For example, recent database submissions have revealed that the
ORF predicted from the genomic and cDNA sequences of AtFtsZ2-1 (Table 1,
accession number AAD21440) is shorter at its 50 end than the ORFs predicted
for several other FtsZ2 genes. However, inspection of the AtFtsZ2-1 gene sequence reveals a second potential in-frame start codon 243 nucleotides (81 codons)
P1: GDL
April 12, 2001
9:48
Annual Reviews
AR129-12
PLASTID DIVISION
321
upstream of the predicted start site that corresponds closely to the position of the
predicted start codon for a second Arabidopsis FtsZ2 gene, AtFtsZ2-2 (Table 1,
accession number CAB89263). This suggests the possibility of misidentified or
perhaps multiple translational initiation sites among FtsZ2 genes. Analysis of the
longer AtFtsZ2-1 ORF by TargetP changes the chloroplast targeting score from
0.161 to a more ambiguous 0.555 and suggests that the corresponding polypeptide
might be imported into chloroplasts if tested in an in vitro import assay, contrary
to the results obtained using the shorter ORF (Table 1) (83). In addition, targeting
prediction programs may be less reliable for proteins from taxonomic groups that
are not well represented in the datasets used in their construction. Nevertheless,
these programs have been invaluable for revealing a potentially important distinction between FtsZ1 and FtsZ2 proteins in flowering plants, though whether both
groups are represented in all land plants, and if so whether the apparent correlation
between family grouping and presence or absence of a transit peptide will apply
in all cases, remains unknown.
Although the localization predictions suggest that many FtsZ2 proteins are not
targeted to the stromal compartment, an essential role for them in plastid division
was established by the finding that Arabidopsis plants expressing an AtFtsZ21 antisense transgene exhibited defects in chloroplast division identical to those
observed in the AtFtsZ1-1 antisense plants (83) (Figure 1C). The demonstration
that AtFtsZ1-1 protein levels were unaffected in the AtFtsZ2-1 antisense plants
and vice versa (110) confirmed that FtsZ1 and FtsZ2 proteins, though postulated
to be localized in different subcellular compartments, each played essential but
functionally distinct roles in plastid division.
Proposed Functions of FtsZ1 and FtsZ2
A consistent feature of recent models depicting the macromolecular configuration
of the plastid division apparatus is inclusion of the stromal and cytosolic PD rings
observed in ultrastructural studies (47, 65, 79). Osteryoung et al (83) incorporated
the results of the AtFtsZ1-1 and AtFtsZ2-1 chloroplast import and antisense experiments described above into these models by postulating that, in higher plants,
FtsZ1 and FtsZ2 proteins are components of the stromal and cytosolic PD rings,
respectively, functioning together on the two envelope surfaces to constrict the organelle. In partial support of this model, recent immunofluorescence localization
studies have confirmed that FtsZ1 and FtsZ2 proteins colocalize to rings at the plastid midpoint in Arabidopsis and other plants (Figure 3) (S Vitha, RS McAndrew,
& KW Osteryoung, submitted). Based on the combination of targeting predictions
and in vitro chloroplast import experiments (32, 84), it can be reasonably concluded that the FtsZ1-containing ring is localized inside the chloroplast, though
whether it represents the same structure as the stromal PD ring remains to be
established. However, higher resolution techniques are needed to determine precisely where with respect to the envelope membranes the FtsZ2-containing ring
is situated. Though its proposed localization on the cytosolic surface of the outer
P1: GDL
April 12, 2001
322
9:48
Annual Reviews
OSTERYOUNG
¥
AR129-12
MCANDREW
envelope is consistent with the position of the cytosolic PD ring, these two rings
may represent distinct structures, and the FtsZ2 ring may be positioned along with
FtsZ1 inside the stromal compartment. If this were true, it would suggest the possibility that FtsZ1 and FtsZ2 form heterodimers in vivo, as do α- and β-tubulin (78).
Alternatively, FtsZ2 could be localized in the intermembrane space, a possibility
suggested by the recent description of a third PD ring positioned between the envelope membranes in the red alga C. merolae (66). Complicating this issue further
is the fact that it has not yet been determined whether there is functional overlap
between AtFtsZ2-1 and AtFtsZ2-2. The arrangement of the FtsZ1 and FtsZ2 rings
remains a major outstanding issue with important implications for understanding
how the plastid division complex as a whole is organized and how its component
proteins function together to achieve organelle constriction.
COMPARISON BETWEEN PLANT FTSZ1
AND FTSZ2 PROTEINS
Although the predicted presence or absence of a chloroplast transit peptide in
higher plant FtsZ proteins is correlated with their assignment to the FtsZ1 or
FtsZ2 family, respectively, the phylogenetic analyses responsible for these assignments (Figure 2) are based on sequence alignments that exclude the amino-terminal
portions of the proteins, which are highly variable and thus phylogenetically uninformative. It is therefore instructive to consider structural properties in addition
to the transit peptides that distinguish FtsZ1 and FtsZ2 proteins from one another.
This is best accomplished by comparing the plant proteins with their well-studied
bacterial counterparts.
Structural Domains
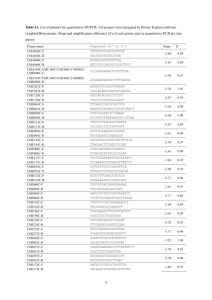
In general, plant FtsZ proteins share most of the structural features common to
the bacterial proteins (26, 53, 54). These features are highlighted in Figures 4, 5,
and 6, which represent the primary (excluding the extreme amino-terminal end),
secondary, and tertiary structures, respectively, of FtsZ1 and FtsZ2. All FtsZs can
be divided into two major structural domains: a highly conserved N-terminal domain and a less well-conserved C-terminal domain. The N-terminal domain forms
a Rossmann fold (53, 103), typical of the GTPase domain of Ras and other Gproteins. This structure contains the “tubulin signature motif,” GGGTG(T/S)G,
required for GTP binding in FtsZs and tubulins (15, 25, 56, 73, 99) (note double
underline in Figure 4) as well as residues that contact the guanine nucleotide
(black bars) and are essential for GTP-hydrolysis (red diamonds) (120). These
residues are completely conserved in the plant sequences with one exception.
In FtsZ1 proteins, an alanine residue essential for GTP-binding and hydrolysis
in the bacterial proteins has been replaced by a threonine residue (circled red
diamond). This may be important for its function in the chloroplast stroma, as
just prior to this substitution there is a helical structure (H3a) (Figure 5B) not
present in FtsZ2 proteins that corresponds to the only region in the N-terminal
P1: GDL
April 12, 2001
9:48
Annual Reviews
AR129-12
PLASTID DIVISION
323
domain in which FtsZ1 and FtsZ2 differ significantly in their hydrophobicity
(Figure 5A). Otherwise, the secondary (Figure 5B) and tertiary (Figure 6) structures of the N-terminal domains in FtsZ1, FtsZ2, and the bacterial FtsZ proteins
(26, 53) are nearly identical. In addition, there is a stretch of amino acids at the
amino-terminal end of E. coli FtsZ, protruding from the otherwise globular structure, that forms a small helix and is required for FtsZ ring assembly in vivo (120).
This helix is not evident in the plant structures (Figure 5B), but the residues at the
amino-terminal ends of the FtsZ1 and FtsZ2 proteins are also predicted to protrude
(Figure 6) and may be involved in assembly of the chloroplast FtsZ rings in plants.
The C-terminal domain of FtsZ is more variable among proteins from different
organisms and appears to play a more regulatory role. However, some conserved
features can be distinguished. Among these are three highly conserved “synergy”
residues that regulate GTP hydrolysis and are essential for cell division in E. coli,
although the C-terminal domain is dispensable for GTPase activity and FtsZ polymerization per se (120). These synergy residues are completely conserved in the
plant FtsZ1 and FtsZ2 families (Figure 4, red dots; Figure 6, stars). Also present in
the C-terminal domain are loop structures that have been implicated in the binding
of calcium (53), which is thought to stabilize FtsZ polymer networks formed in vitro
(29, 75, 109, 113, 123). Similar loop structures are predicted in FtsZ1 and FtsZ2,
and they may participate in the assembly of plant FtsZs into multimers, which
has been reported for an FtsZ1 protein from pea (32). The carboxyl end of the
C-terminal domain contains a high proportion of hydrophilic residues exposed at
the surface of the protein (Figure 5A), and it resembles the microtubule-associated
protein (MAP) binding regions of tubulin (20). Within this region is a small, highly
conserved sequence, D/E-I/V-P-X-F/Y-L, termed the C-terminal core domain (60),
which is required for direct interactions between FtsZ and two other essential cell
division proteins in E. coli, ZipA and FtsA (21, 37, 52, 60, 70, 71, 120, 122). ZipA
is an integral membrane protein with homology to MAPs that stabilizes the FtsZ
ring structure and may be involved in anchoring it to the bacterial cell membrane
(35–37, 98). FtsA is a peripheral membrane protein related to the actin/HSP70
superfamily of ATPases and also functions in part to support the FtsZ ring structure (4, 35, 61, 106). Interestingly, the C-terminal core domain and associated secondary structure are conserved in the plant FtsZ2 proteins (Figure 4, red box;
Figure 5B, H11; Figure 6, H11), but they are missing from FtsZ1. This suggests
that proteins similar to ZipA and/or FtsA may interact specifically with FtsZ2,
and it reveals a structural distinction between the two protein families that may be
relevant to their distinct functions in plastid division.
The N-terminal and C-terminal domains are connected and clearly delimited
by a long central helix (Figure 5B, H7; Figure 6, H7). In bacteria, this helix
contains additional residues that contact the guanine nucleotide (53). The linker
helix and associated GTP-binding residues are also conserved in both FtsZ1 and
FtsZ2 (Figure 4, black bars in unboxed region). The presence in both plant proteins
of all structural elements required for FtsZ activity in bacteria implies that the
FtsZ1 and FtsZ2 proteins catalyze similar reactions even though they have distinct
functions and may be localized in different subcellular compartments.
P1: GDL
April 12, 2001
324
9:48
Annual Reviews
OSTERYOUNG
¥
AR129-12
MCANDREW
Posttranslational Modification Motifs
Posttranslational modifications of FtsZ proteins have not been documented so far,
but they occur for tubulins. In animal systems, phosphorylation by various kinases
mediates tubulin assembly and stability, microtubule localization, and binding of
tubulin and microtubules to other proteins such as MAPs (61a). Related modifications of tubulins also occur in higher plants (108a). Analysis of FtsZ1 and FtsZ2 for
posttranslational modification consensus motifs using various online resources reveals a number of potential phosphorylation sites that are unique to, but conserved
within, each family (Figure 5B). For example, FtsZ2 proteins share one tyrosine kinase recognition site (Figure 5B, blue box) and one cGMP-dependent kinase recognition site (red box) that are not present in FtsZ1 proteins. Likewise, FtsZ1 proteins
may be uniquely phosphorylated by casein kinase II (Figure 5B, red diamond).
Although the consensus phosphorylation sites indicated in Figure 5B are derived
primarily from studies of kinases from nonplant systems, kinases with comparable
activities exist in plants (34a, 52a, 114a, 119a). Other kinds of modifications that
are not easily predicted are also possible, such as palmitoylation, which mediates
attachment of tubulin to membranes (10, 124). Although the predicted posttranslational modifications shown in Figure 5B would have to be tested experimentally
to determine whether they occur in vivo, they nevertheless provide potential clues
to mechanisms that may be important for regulating FtsZ1 and FtsZ2 function.
PLACEMENT OF THE PLASTID DIVISION MACHINERY
The many images of intact, dividing plastids present in the literature, whether obtained by conventional light microscopy (51, 68, 92, 100, 102), three-dimensional
reconstruction (31, 67), or scanning electron microscopy (12, 65), invariably show
the constriction positioned near the center of the organelle, perpendicular to the
longitudinal axis. This implies the existence of a mechanism for ensuring proper
placement of the division machinery. Recent data have revealed that this process
also has a prokaryotic origin, having evolved from the mechanism controlling the
site of cell division in many bacteria.
Selection of the Cell Division Site in Bacteria
In E. coli, placement of the FtsZ ring is governed by the minB operon, which
encodes three gene products: MinC, MinD, and MinE (reviewed in 57, 104, 105,
112). Genetic analysis has shown that the activities of these proteins exhibit a
complicated pattern of interdependency. In wild-type cells, MinC acts as a division
inhibitor by suppressing formation of the FtsZ ring at all sites except the cell center
(6, 17, 18) in part by destabilizing FtsZ polymerization at inappropriate sites (41).
This activity is dependent on MinD, which is thought to form a heterodimer with
MinC (42). Localization studies in E. coli that are based on fusions to green fluorescent protein have revealed that MinD is associated with the cell membrane but
only on one side of the cell at a time. MinD oscillates between the two cell poles
P1: GDL
April 12, 2001
9:48
Annual Reviews
AR129-12
PLASTID DIVISION
325
with a periodicity of less than one minute and is thought to carry along as cargo
the MinC division inhibitor, which exhibits a similar membrane association and
oscillatory behavior (40, 95, 96). The oscillation of MinD requires MinE, which
localizes to a ring at the midcell independently of FtsZ and in turn requires MinD
for its activity (97). The MinE ring prevents MinCD from acting at the cell center,
thereby allowing FtsZ ring assembly, and hence cell division, only at that position.
The min locus is so-named because mutations in minC or minD allow the FtsZ ring
to assemble at aberrant sites near the cell poles, resulting in the formation of minicells that lack chromosomes and cannot expand (16). Mutations in minE, on the
other hand, permit MinCD to act ectopically at the midcell, thereby preventing FtsZ
ring assembly at all sites and resulting in the formation of bacterial filaments. The
mechanisms responsible for this complex situation are not yet fully understood.
Genes encoding MinC, MinD, and MinE are also found in numerous other
prokaryotes, including the cyanobacterium Synechocystis PCC6803 (43). MinE is
absent in Bacillus subtilis, however, and exclusion of the MinCD inhibitor from
the cell center is accomplished by a different protein (104).
Role of MinD in Plastid Division Site Selection
That placement of the plastid division machinery in photosynthetic eukaryotes
is mediated by a mechanism related to that in bacteria was strongly suggested
by the discovery that homologues of MinD and MinE are encoded in the plastid
genomes of the unicellular algae Chlorella vulgaris (119) and Guillardia theta
(22). MinD is also encoded in the plastid genomes of the algae Nephroselmis olivacea (118) and Prototheca wickerhamii (accession number CAB53105) as well
as the nuclear genomes of diverse higher plants, including Arabidopsis, rice, and
marigold (14, 69). The MinD proteins from Arabidopsis and marigold bear cleavable chloroplast transit peptides that direct their import into isolated chloroplasts
(14; J Froehlich & KW Osteryoung, unpublished results). MinC genes have not
been identified in any photosynthetic eukaryote.
A role for MinD in positioning of the chloroplast division apparatus has recently
been confirmed. Using an antisense approach, Colletti et al (14) demonstrated that
reduced expression of AtMinD1, which encodes the chloroplast-targeted MinD
homologue from Arabidopsis, yielded plants that had asymmetrically constricted
plastids, indicating misplacement of the division apparatus and an abnormally high
degree of heterogeneity in chloroplast size and number, resembling the bacterial
minicell phenotype (Figure 1D). Similar phenotypes have been described in the
Arabidopsis mutant arc11 (63), which may be allelic with AtMinD1 (14). The
antisense phenotypes also had implications for the order in which components of
the plastid division apparatus are assembled. The asymmetric constriction suggested that reduced levels of plastid-targeted MinD allows misplacement of the
plastid-localized FtsZ1 ring, which in turn leads to misplacement of other division
components and hence to asymmetric division. Therefore, these results suggested
that placement of the entire plastid division apparatus is normally determined from
inside the chloroplast by the position of the FtsZ1 ring (14).
P1: GDL
April 12, 2001
326
9:48
Annual Reviews
OSTERYOUNG
¥
AR129-12
MCANDREW
ADDITIONAL COMPONENTS OF THE PLASTID
DIVISION APPARATUS
Currently, FtsZ, MinD, and MinE are the only obvious homologues of bacterial
cell division genes known to exist in photosynthetic eukaryotes, and a role for
MinE in plastid division has yet to be demonstrated. Because the functions of most
of the other bacterial cell division proteins are unknown, they provide few clues
regarding the likelihood that functional counterparts might participate in plastid
division. However, at least nine proteins localize to the division septum in E. coli
(62, 104), and the plastid division apparatus is likely to be at least as complex (82).
One potentially rich source of new plastid division genes is represented by the
Arabidopsis arc mutants (accumulation and replication of chloroplasts). These mutants, which exhibit various abnormalities in chloroplast number and size, define
at least 11 loci that are important in the control of plastid division and expansion
in plants (63, 87, 89–91, 93). The arc6 mutation, which causes drastically reduced
numbers of enlarged chloroplasts and proplastids (93, 94) resembling those in
plants expressing the AtFtsZ1-1 and AtFtsZ2-1 antisense transgenes (83; see also
Figure 1B & 1C), is not allelic with any of the FtsZ genes in Arabidopsis (KA
Pyke, personal communication) and may represent a regulatory step in plastid division (63). In most of the other arc mutants, the nature of the affected locus is not
obvious and could be either structural or regulatory. The phenotype of arc5, however, in which the chloroplasts fail to separate completely and remain permanently
constricted (102), strongly suggests that the ARC5 gene product is a late-acting
structural component of the division machinery. Cloning of this and other arc loci
should reveal additional plastid division proteins and provide new insights as to
how plastid division in plants is regulated.
Other approaches likely to yield new plastid division components include
identification of proteins that interact directly with plant FtsZ proteins and
proteomics-based strategies that rely on isolation of plastid division complexes.
Progress toward the latter goal is represented by the recent isolation of intact chloroplasts from synchronized cultures of C. merolae to which PD rings remain visibly
attached (65).
FUTURE CHALLENGES
As molecular dissection of the plastid division machinery moves forward, many of
the questions alluded to throughout this review will begin to find answers. These
include: (a) What is the composition of the plastid division apparatus? (b) How and
in what order are its component parts assembled? (c) How are they topologically
organized? (d) What are their biochemical activities and how do they affect constriction of the two envelope membranes? (e) How are these activities coordinated?
( f ) How is the division site specified and recognized? Because of the evolutionary relationship between plastid and prokaryotic cell division, bacterial models
P1: GDL
April 12, 2001
9:48
Annual Reviews
AR129-12
PLASTID DIVISION
327
will no doubt continue to inform investigation of these questions, though the finding that multiple FtsZ genes participate in plastid division already suggests some
fundamental differences between the plastid and bacterial cell division processes
(82). It will also be of interest to learn whether there are differences between the
plastid division machineries in algae and land plants that might shed light on the
evolutionary steps that gave rise to the FtsZ1 and FtsZ2 gene families during land
plant evolution (33).
Related questions that remain largely unexplored include the following: (a) Do
active mechanisms exist for controlling plastid DNA segregation and thylakoid
membrane partitioning to daughter plastids during division? The former possibility has been suggested by the discovery of a plastid DNA binding protein associated
with the envelope membranes (107, 108). (b) Is there an active mechanism governing plastid segregation during cell division? The fact that aplastidic cells are
not observed (except in guard cells) (94) suggests this may be the case. (c) How is
total plastid compartment volume perceived and regulated? That such processes
exist is implied by the finding that decreases in plastid number are almost always compensated for by corresponding increases in plastid size and vice versa
(87, 90). (d) How are plastid division and expansion integrated into the plant’s
developmental program? A full understanding of plastid division will ultimately
require investigation of all these issues.
Visit the Annual Reviews home page at www.AnnualReviews.org
LITERATURE CITED
1. Addinall SG, Bi E, Lutkenhaus J. 1996. FtsZ
ring formation in fts mutants. J. Bacteriol.
178:3877–84
2. Beech PL, Gilson PR. 2000. FtsZ and organelle division in protists. Protist 151:11–
16
3. Beech PL, Nheu T, Schultz T, Herbert S,
Lithgow T, et al. 2000. Mitochondrial FtsZ
in a chromophyte alga. Science 287:1276–
79
4. Begg K, Nikolaichik Y, Crossland N,
Donachie WD. 1998. Roles of FtsA and FtsZ
in activation of division sites. J. Bacteriol.
180:881–84
5. Bi E, Lutkenhaus J. 1991. FtsZ ring structure associated with division in Escherichia
coli. Nature 354:161–64
6. Bi E, Lutkenhaus J. 1993. Cell division inhibitors SulA and MinCD prevent formation
of the FtsZ ring. J. Bacteriol. 175:1118–25
6a. Boffey SA, Lloyd D, eds. 1988. Division and Segregation of Organelles. Cambridge, UK: Cambridge Univ. Press
7. Bramhill D. 1997. Bacterial cell division. Annu. Rev. Cell Dev. Biol. 13:395–
424
8. Bramhill D, Thompson CM. 1994. GTPdependent polymerization of Escherichia
coli FtsZ protein to form tubules.
Proc. Natl. Acad. Sci. USA 91:5813–
17
9. Butterfass T. 1988. Nuclear control of plastid division. See Ref. 6a, pp. 21–38
10. Caron JM. 1997. Posttranslational modification of tubulin by palmitoylation: I. In
vivo and cell-free studies. Mol. Biol. Cell
8:621–36
11. Chaly N, Possingham JV. 1981. Structure
of constricted proplastids in meristematic
plant tissues. Biol. Cell 41:203–9
P1: GDL
April 12, 2001
328
9:48
Annual Reviews
OSTERYOUNG
¥
AR129-12
MCANDREW
12. Chaly N, Possingham JV, Thomson WW.
1980. Chloroplast division in spinach
leaves examined by scanning electron microscopy and freeze-etching. J. Cell Sci.
46:87–96
13. Chida Y, Ueda K. 1991. Division of chloroplasts in a green alga, Trebouxia potteri.
Ann. Bot. 67:435–42
14. Colletti KS, Tattersall EA, Pyke KA,
Froelich JE, Stokes KD, Osteryoung KW.
2000. A homologue of the bacterial cell division site-determining factor MinD mediates placement of the chloroplast division
apparatus. Curr. Biol. 10:507–16
15. de Boer P, Crossley R, Rothfield L. 1992.
The essential bacterial cell-division protein
FtsZ is a GTPase. Nature 359:254–56
16. de Boer PAJ, Crossley RE, Rothfield LI.
1988. Isolation and properties of minB,
a complex genetic locus involved in correct placement of the division site in Escherichia coli. J. Bacteriol. 170:2106–12
17. de Boer PAJ, Crossley RE, Rothfield LI.
1989. A division inhibitor and a topological
specificity factor coded for by the minicell
locus determine proper placement of the
division septum in E. coli. Cell 56:641–49
18. de Boer PAJ, Crossley RE, Rothfield LI.
1992. Roles of MinC and MinD in the
site-specific septation block mediated by
the MinCDE system of Escherichia coli. J.
Bacteriol. 174:63–70
19. Den Blaauwen T, Buddelmeijer N, Aarsman ME, Hameete CM, Nanninga N. 1999.
Timing of FtsZ assembly in Escherichia
coli. J. Bacteriol. 181:5167–75
20. Desai A, Mitchison TJ. 1998. Tubulin and
FtsZ structures: functional and therapeutic
implications. BioEssays 20:523–27
21. Din N, Quardokus EM, Sackett MJ, Brun
YV. 1998. Dominant C-terminal deletions
of FtsZ that affect its ability to localize in
Caulobacter and its interaction with FtsA.
Mol. Microbiol. 27:1051–63
22. Douglas SE, Penny SL. 1999. The plastid
genome of the cryptophyte alga, Guillardia
theta: complete sequence and conserved
23.
24.
25.
26.
27.
28.
29.
30.
31.
32.
33.
synteny groups confirm its common ancestry with red algae. J. Mol. Evol. 48:236–44
Duckett JG, Ligrone R. 1993. Plastiddividing rings in ferns. Ann. Bot. 72:619–
27
Emanuelsson O, Nielsen H, Brunak S, von
Heijne G. 2000. Predicting subcellular localization of proteins based on their Nterminal amino acid sequence. J. Mol. Biol.
300:1005–16
Erickson HP. 1997. FtsZ, a tubulin homologue in prokaryote cell division. Trends
Cell Biol. 7:362–67
Erickson HP. 1998 Atomic structures of
tubulin and FtsZ. Trends Cell Biol. 8:133–
37
Erickson HP. 2000. Dynamin and FtsZ:
missing links in mitochondrial and bacterial division. J. Cell Biol. 148:1103–5
Erickson HP, Stoffler D. 1996. Protofilaments and rings, two conformations of
the tubulin family conserved from bacterial FtsZ to α/β and γ tubulin. J. Cell Biol.
135:5–8
Erickson HP, Taylor DW, Taylor KA,
Bramhill D. 1996. Bacterial cell division
protein FtsZ assembles into protofilament
sheets and minirings, structural homologs
of tubulin polymers. Proc. Natl. Acad. Sci.
USA 93:519–23
Faguy DM, Doolittle WF. 1998. Cytoskeletal proteins: the evolution of cell division.
Curr. Biol. 8:R338–41
Gaffal KP, Arnold CG, Friedrichs GJ,
Gemple W. 1995. Morphodynamical
changes of the chloroplast of Chlamydomonas reinhardtii during the 1st round
of division. Arch. Protistenkd. 145:10–23
Gaikwad A, Babbarwal V, Pant V, Mukherjee SK. 2000. Pea chloroplast FtsZ can
form multimers and correct the thermosensitive defect of an Escherichia coli ftsZ mutant. Mol. Gen. Genet. 263:213–21
Gilson PR, Beech PL. 2000. Cell division
protein FtsZ: running rings around bacteria, chloroplasts and mitochondria. Res.
Microbiol. In press
P1: GDL
April 12, 2001
9:48
Annual Reviews
AR129-12
PLASTID DIVISION
34. Guex N, Peitsch MC. 1997. SWISSMODEL and the Swiss-PdbViewer:
an environment for comparative protein modeling. Electrophoresis 18:2714–
23
34a. Guillén G, Valdlés-López V, Noguez R,
Olivares J, Rodriguez-Zapata LC, et al.
1999. Profilin in Phaseolus vulgaris is encoded by two genes (only one expressed
in root nodules) but multiple isoforms are
generated in vivo by phosphorylation on
tyrosine residues. Plant J. 19:497–508
35. Hale CA, de Boer PAJ. 1999. Recruitment
of ZipA to the septal ring of Escherichia
coli is dependent on FtsZ and independent
of FtsA. J. Bacteriol. 181:167–76
36. Hale CA, de Boer PAJ. 1997. Direct binding of FtsZ to ZipA, an essential component of the septal ring structure that
mediates cell division in E. coli. Cell 88:
175–85
37. Hale CA, Rhee AC, de Boer PA. 2000.
ZipA-induced bundling of FtsZ polymers mediated by an interaction between C-terminal domains. J. Bacteriol.
182:5153–66
38. Hashimoto H. 1986. Double ring structure around the constricting neck of dividing plastids of Avena sativa. Protoplasma
135:166–72
39. Hashimoto H, Possingham JV. 1989. Division and DNA distribution in ribosomedeficient plastids of the barley mutant “albostrians.” Protoplasma 149:20–23
40. Hu Z, Lutkenhaus J. 1999. Topological
regulation of cell division in Escherichia
coli involves rapid pole to pole oscillation
of the division inhibitor MinC under the
control of MinD and MinE. Mol. Microbiol. 34:82–90
41. Hu Z, Mukherjee A, Pichoff S, Lutkenhaus J. 1999. The MinC component of
the division site selection system in Escherichia coli interacts with FtsZ to prevent polymerization. Proc. Natl. Acad.
Sci. USA 96:14819–24
42. Huang J, Cao C, Lutkenhaus J. 1996. In-
43.
43a.
44.
45.
46.
47.
48.
49.
50.
51.
329
teraction between FtsZ and inhibitors of
cell division. J. Bacteriol. 178:5080–85
Kaneko T, Sato S, Kotani H, Tanaka A,
Asamizu E, et al. 1996. Sequence analysis of the genome of the unicellular
cyanobacterium Synechocystis sp. strain
PCC6803. II. Sequence determination of
the entire genome and assignment of potential protein-coding regions. DNA Res.
3:109–36
Kiessling J, Kruse S, Rensing SA, Harter
K, Decker EL, Reski R. 2000. Visualization of a cytoskeleton-like FtsZ network
in chloroplasts. J. Cell Biol. 151:945–50
Kreegipuu A, Blom N, Brunak S. 1999.
PhosphoBase, a database of phosphorylation sites: release 2.0. Nucleic Acids Res.
27:237–39
Kuroiwa T. 1989. The nuclei of cellular
organelles and the formation of daughter
organelles by the “plastid-dividing ring”.
Bot. Mag. 102:291–329
Kuroiwa T. 1991. The replication, differentiation, and inheritance of plastids with
emphasis on the concept of organelle nuclei. Int. Rev. Cytol. 128:1–62
Kuroiwa T, Kuroiwa H, Sakai A, Takahashi H, Toda K, Itoh R. 1998. The
division apparatus of plastids and mitochondria. Int. Rev. Cytol. 181:1–41
Leech RM. 1976. The replication of plastids in higher plants. In Cell Division in
Higher Plants, ed. MM Yeoman, pp. 135–
59. London: Academic
Leech RM. 1986. Stability and plasticity
during chloroplast development. In Plasticity in Plants, Vol. 40, ed. JW Jennings,
AJ Trewavas, pp. 121–53. Cambridge,
UK: Cambridge Univ. Press
Leech RM, Pyke KA. 1988. Chloroplast
division in higher plants with particular
reference to wheat. See Ref. 6a, pp. 39–
62
Leech RM, Thomson WW, Platt-Aloia
KA. 1981. Observations on the mechanism of chloroplast division in higher
plants. New Phytol. 87:1–9
P1: GDL
April 12, 2001
330
9:48
Annual Reviews
OSTERYOUNG
¥
AR129-12
MCANDREW
52. Liu Z, Mukherjee A, Lutkenhaus J. 1999.
Recruitment of ZipA to the division site
by interaction with FtsZ. Mol. Microbiol.
31:1853–61
52a. Loog M, Toomik R, Sak K, Muszynska G,
Jarv J, Ek P. 2000. Peptide phosphorylation by calcium-dependent protein kinase
from maize seedlings. Eur. J. Biochem.
267:337–43
53. Löwe J. 1998. Crystal structure determination of FtsZ from Methanococcus jannaschii. J. Struct. Biol. 124:235–43
54. Löwe J, Amos LA. 1998. Crystal structure of the bacterial cell-division protein
FtsZ. Nature 391:203–6
55. Lu C, Reedy M, Erickson HP. 2000.
Straight and curved conformations of
FtsZ are regulated by GTP hydrolysis. J.
Bacteriol. 182:164–70
56. Lutkenhaus J. 1993. FtsZ ring in bacterial
cytokinesis. Mol. Microbiol. 9:403–9
57. Lutkenhaus J. 1998. The regulation of
bacterial cell division: a time and place
for it. Curr. Opin. Microbiol. 1:210–
15
58. Lutkenhaus J, Addinall SG. 1997. Bacterial cell division and the Z ring. Annu.
Rev. Biochem. 66:93–116
59. Lutkenhaus J, Wolf-Watz H, Donachie
WD. 1980. Organization of genes in the
ftsZ-envA region of the Escherichia coli
genes map and identification of a new
fts locus ( ftsZ ). J. Bacteriol. 142:615–
20
60. Ma XL, Margolin W. 1999. Genetic and
functional analyses of the conserved Cterminal core domain of Escherichia coli
FtsZ. J. Bacteriol. 181:7531–44
61. Ma XL, Sun Q, Wang R, Singh G,
Jonietz EL, Margolin W. 1997. Interactions between heterologous FtsA and FtsZ
proteins at the FtsZ ring. J. Bacteriol.
179:6788–97
61a. MacRae TH. 1997. Tubulin post-translational modifications—enzymes and their
mechanisms of action. Eur. J. Biochem.
244:265–78
62. Margolin W. 1998. A green light for the
bacterial cytoskeleton. Trends Microbiol.
6:233–38
63. Marrison JL, Rutherford SM, Robertson
EJ, Lister C, Dean C, Leech RM. 1999.
The distinctive roles of five different ARC
genes in the chloroplast division process in
Arabidopsis. Plant J. 18:651–62
64. Mita T, Kuroiwa T. 1988. Division of plastids by a plastid-dividing ring in Cyanidium caldarium. Protoplasma 1(Suppl.):
133–52
65. Miyagishima S, Itoh R, Aita S, Kuroiwa
H, Kuroiwa T. 1999. Isolation of dividing
chloroplasts with intact plastid-dividing
rings from a synchronous culture of the unicellular red alga Cyanidioschyzon merolae.
Planta 209:371–75
66. Miyagishima S, Itoh R, Toda K, Takahashi
H, Kuroiwa H, Kuroiwa T. 1998. Identification of a triple ring structure involved
in plastid division in the primitive red alga
Cyanidioschyzon merolae. J. Electron Microsc. 47:269–72
67. Miyagishima S, Itoh R, Toda K, Takahashi H, Kuroiwa H, Kuroiwa T. 1998.
Orderly formation of the double ring structures for plastid and mitochondrial division
in the unicellular red alga Cyanidioschyzon
merolae. Planta 206:551–60
68. Modrusan Z, Wrischer M. 1990. Studies on
chloroplast division in young leaf tissues of
some higher plants. Protoplasma 154:1–7
69. Moehs CP, Tian L, Osteryoung KW, DellaPenna D. 2001. Analysis of carotenoid
biosynthetic gene expression during
marigold petal development. Plant Mol.
Biol. In press
70. Mosyak L, Zhang Y, Glasfeld E, Haney S,
Stahl M, et al. 2000. The bacterial celldivision protein ZipA and its interaction
with an FtsZ fragment revealed by X-ray
crystallography. EMBO J. 19:3179–91
71. Moy FJ, Glasfeld E, Mosyak L, Powers R.
2000. Solution structure of ZipA, a crucial
component of Escherichia coli cell division. Biochemistry 39:9146–56
P1: GDL
April 12, 2001
9:48
Annual Reviews
AR129-12
PLASTID DIVISION
72. Mukherjee A, Dai K, Lutkenhaus J. 1993.
Escherichia coli cell division protein FtsZ
is a guanine nucleotide binding protein.
Proc. Natl. Acad. Sci. USA 90:1053–57
73. Mukherjee A, Lutkenhaus J. 1994.
Guanine nucleotide-dependent assembly
of FtsZ into filaments. J. Bacteriol.
176:2754–58
74. Mukherjee A, Lutkenhaus J. 1998. Dynamic assembly of FtsZ regulated by GTP
hydrolysis. EMBO J. 17:462–69
75. Mukherjee A, Lutkenhaus J. 1999. Analysis of FtsZ assembly by light scattering and
determination of the role of divalent metal
cations. J. Bacteriol. 181:823–32
76. Nanninga N. 1998. Morphogenesis of Escherichia coli. Microbiol. Mol. Biol. Rev.
62:110–29
77. Nogales E, Downing KH, Amos LA, Löwe
J. 1998. Tubulin and FtsZ form a distinct family of GTPases. Nat. Struct. Biol.
5:451–58
78. Nogales E, Wolf SG, Downing KH. 1998.
Structure of the α/β tubulin dimer by
electron crystallography. Nature 391:199–
203
79. Ogawa S, Ueda K, Noguchi T. 1994. Division apparatus of the chloroplast in Nannochloris bacillaris (Chlorophyta). J. Phycol. 31:132–37
80. Oross JW, Possingham JV. 1989. Ultrastructural features of the constricted region of dividing plastids. Protoplasma
150:131–38
81. Osteryoung KW. 2000. Organelle fission:
crossing the evolutionary divide. Plant
Physiol. 123:1213–16
82. Osteryoung KW, Pyke KA. 1998. Plastid
division: evidence for a prokaryotically derived mechanism. Curr. Opin. Plant Biol.
1:475–79
83. Osteryoung KW, Stokes KD, Rutherford
SM, Percival AL, Lee WY. 1998. Chloroplast division in higher plants requires
members of two functionally divergent
gene families with homology to bacterial
ftsZ. Plant Cell 10:1991–2004
331
84. Osteryoung KW, Vierling E. 1995. Conserved cell and organelle division. Nature
376:473–74
85. Peitsch MC. 1996. ProMod and SwissModel: Internet-based tools for automated comparative protein modelling.
Biochem. Soc. Trans. 24:274–79
85a. Possingham JV, Hashimoto H, Oross J.
1988. Factors that influence plastid division in higher plants. See Ref. 6a, pp. 1–
20
86. Possingham JV, Lawrence ME. 1983.
Controls to plastid division. Int. Rev. Cytol. 84:1–56
87. Pyke KA. 1997. The genetic control of
plastid division in higher plants. Am. J.
Bot. 84:1017–27
88. Pyke KA. 1999. Plastid division and development. Plant Cell 11:549–56
89. Pyke KA, Leech RM. 1991. Rapid image
analysis screening procedure for identifying chloroplast number mutants in mesophyll cells of Arabidopsis thaliana (L.)
Heynh. Plant Physiol. 96:1193–95
90. Pyke KA, Leech RM. 1992. Chloroplast
division and expansion is radically altered by nuclear mutations in Arabidopsis
thaliana. Plant Physiol. 99:1005–8
91. Pyke KA, Leech RM. 1994. A genetic analysis of chloroplast division and
expansion in Arabidopsis thaliana. Plant
Physiol. 104:201–7
92. Pyke KA, Page AM. 1998. Plastid
ontogeny during petal development in
Arabidopsis. Plant Physiol. 116:797–
803
93. Pyke KA, Rutherford SM, Robertson EJ,
Leech RM. 1994. arc6, a fertile Arabidopsis mutant with only two mesophyll cell
chloroplasts. Plant Physiol. 106:1169–
77
94. Pyke KA, Rutherford SM, Robertson
EJ, Leech RM. 1994. arc6, an extreme
chloroplast division mutant of Arabidopsis also alters proplastid proliferation and
morphology in shoot and root apices. J.
Cell. Sci. 108:2937–44
P1: GDL
April 12, 2001
332
9:48
Annual Reviews
OSTERYOUNG
¥
AR129-12
MCANDREW
95. Raskin DM, de Boer PAJ. 1999. MinDEdependent pole-to-pole oscillation of division inhibitor MinC in Escherichia coli.
J. Bacteriol. 181:6419–24
96. Raskin DM, de Boer PAJ. 1999. Rapid
pole-to-pole oscillation of a protein required for directing division to the middle
of Escherichia coli. Proc. Natl. Acad. Sci.
USA 96:4971–76
97. Raskin DM, de Boer PAJ. 1997. The
MinE ring: an FtsZ-independent cell
structure required for selection of the correct division site in Escherichia coli. Cell
91:685–94
98. RayChaudhuri D. 1999. ZipA is a MAPTau homolog and is essential for structural integrity of the cytokinetic FtsZ ring
during bacterial cell division. EMBO J.
18:2372–83
99. RayChaudhuri D, Park JT. 1992. Escherichia coli cell-division gene ftsZ encodes a novel GTP-binding protein. Nature 359:251–54
100. Ridley SM, Leech RM. 1970. Division of
chloroplasts in an artificial environment.
Nature 227:463–65
101. Rivas G, Lopez A, Mingorance J, Ferrandiz MJ, Zorrilla S, et al. 2000. Magnesium-induced linear self-association of
the FtsZ bacterial cell division protein
monomer. The primary steps for FtsZ assembly. J. Biol. Chem. 275:11740–49
102. Robertson EJ, Rutherford SM, Leech
RM. 1996. Characterization of chloroplast division using the Arabidopsis mutant arc5. Plant Physiol. 112:149–59
103. Rossmann MG, Moras D, Olsen KW.
1974. Chemical and biological evolution of nucleotide-binding protein. Nature
250:194–99
104. Rothfield L, Justice S, Gracı́a-Lara J.
1999. Bacterial cell division. Annu. Rev.
Genet. 33:423–48
105. Rothfield LI, Justice SS. 1997. Bacterial
cell division: the cycle of the ring. Cell
88:581–84
106. Sánchez M, Valencia A, Ferrandiz MJ,
107.
108.
108a.
109.
110.
111.
112.
113.
Sander C, Vicente M. 1994. Correlation
between the structure and biochemical
activities of FtsA, an essential cell division protein of the actin family. EMBO
J. 13:4919–25
Sato N, Albrieux C, Joyard J, Douce
R, Kuroiwa T. 1993. Detection and
characterization of a plastid envelope
DNA-binding protein which may anchor
plastid nucleoids. EMBO J. 12:555–
61
Sato N, Ohshima K, Watanabe A, Ohta
N, Nishiyama Y, et al. 1998. Molecular characterization of the PEND protein, a novel bZIP protein present in the
envelope membrane that is the site of nucleoid replication in developing plastids.
Plant Cell 10:859–72
Smertenko AP, Lawrence SL, Hussey
PJ. 1998. Immunological homologues of
the Arabidopsis thaliana β1 tubulin are
polyglutamylated in Nicotiana tabacum.
Protoplasma 203:138–43
Sossong TM Jr, Brigham-Burke MR,
Hensley P, Pearce KH Jr. 1999. Selfactivation of guanosine triphosphatase
activity by oligomerization of the bacterial cell division protein FtsZ. Biochemistry 38:14843–50
Stokes KD, McAndrew RS, Figueroa R,
Vitha S, Osteryoung KW. 2000. Chloroplast division and morphology are differentially affected by overexpression of
FtsZ1 and FtsZ2 genes in Arabidopsis.
Plant Physiol. 124:1668–77
Strepp R, Scholz S, Kruse S, Speth
V, Reski R. 1998. Plant nuclear gene
knockout reveals a role in plastid division for the homolog of the bacterial cell
division protein FtsZ, an ancestral tubulin. Proc. Natl. Acad. Sci. USA 95:4368–
73
Sullivan SM, Maddock JR. 2000. Bacterial division: finding the dividing line.
Curr. Biol. 10:R249–52
Sun Q, Margolin W. 1998. FtsZ dynamics during the division cycle of
P1: GDL
April 12, 2001
9:48
Annual Reviews
AR129-12
PLASTID DIVISION
114.
114a.
115.
116.
117.
118.
live Escherichia coli cells. J. Bacteriol.
180:2050–56
Suzuki K, Ehara T, Osafune T, Kuroiwa
H, Kawano S, Kuroiwa T. 1994. Behavior of mitochondria, chloroplasts and
their nuclei during the mitotic cycle
in the ultramicroalga Cyanidioschyzon
merolae. J. Cell Biol. 63:280–88
Szczegielniak J, Liwosz A, Jurkowski I,
Loog M, Dobrowolska G, et al. 2000.
Calcium-dependent protein kinase from
maize seedlings activated by phospholipids. Eur. J. Biochem. 267:3818–27
Takahara M, Takahashi H, Matsunaga
S, Miyagishima S, Takano S, et al.
2000. A putative mitochondrial ftsZ gene
is present in the unicellular primitive
red alga Cyanidioschyzon merolae. Mol.
Gen. Genet. 7:245–51
Tewinkel M, Volkmann D. 1987. Observations on dividing plastids in the
protonema of the moss Funaria hygrometrica Sibth.: arrangement of microtubules and filaments. Planta 172:309–
20
Thompson JD, Higgins DG, Gibson TJ.
1994. CLUSTAL W: improving the sensitivity of progressive multiple sequence
alignment through sequence weighting, position-specific gap penalties and
weight matrix choice. Nucleic Acids Res.
22:4673–80
Turmel M, Otis C, Lemieux C. 1999. The
complete chloroplast DNA sequence of
the green alga Nephroselmis olivacea:
119.
119a.
120.
121.
122.
123.
124.
333
insights into the architecture of ancestral
chloroplast genomes. Proc. Natl. Acad.
Sci. USA 96:10248–53
Wakasugi T, Nagai T, Kapoor M, Sugita
M, Ito M, et al. 1997. Complete nucleotide sequence of the chloroplast
genome from the green alga Chlorella
vulgaris: the existence of genes possibly
involved in chloroplast division. Proc.
Natl. Acad. Sci. USA 94:5967–72
Walden R. 1998. The alphabet soup
of plant intracellular signalling: enter
cyclic nucleotides. Curr. Opin. Plant
Biol. 1:419–23
Wang X, Huang J, Mukherjee A, Cao C,
Lutkenhaus J. 1997. Analysis of the interaction of FtsZ with itself, GTP, and
FtsA. J. Bacteriol. 179:5551–59
Whatley JM. 1988. Mechanisms and
morphology of plastid division. See Ref.
6a, pp. 63–84
Yan K, Pearce KH, Payne DJ. 2000. A
conserved residue at the extreme C terminus of FtsZ is critical for the FtsAFtsZ interaction in Staphylococcus aureus. Biochem. Biophys. Res. Commun.
270:387–92
Yu XC, Margolin W. 1997. Ca2+mediated GTP-dependent dynamic assembly of bacterial cell division protein
FtsZ into asters and polymer networks
in vitro. EMBO J. 16:5455–63
Zambito AM, Wolff J. 1997. Palmitoylation of tubulin. Biochem. Biophys. Res.
Commun. 239:650–54
NOTE ADDED IN PROOF
Recent experiments based on cDNAs for the longer AtFtsZ2-1 ORF referred to on
page 321, as well as for AtFtsZ2-2, show both gene products are translocated into
isolated chloroplasts and processed upon import. These results rule out the hypothesis that FtsZ2 proteins in Arabidopsis, and probably other plants, are components
of the cytosolic PD ring.
11:57
Annual Reviews
Figure 3 Immunofluorescence localization of FtsZ1 (top) and FtsZ2 (bottom) in Arabidopsis (left) and pea (right) leaf sections. The right side
of each panel is a bright field image of the immunostained section taken with DIC optics.
April 21, 2001
Figure 1 Phenotypes of Arabidopsis leaf mesophyll cells from wild type (A) or transgenic plants expressing AtFtsZ1-1 (B), AtFtsZ2-1 (C) or
AtMinD1 (D) antisense transgenes. The inset in D shows an asymmetrically dividing petal plastid from an AtMinD1 antisense plant. Bar = 5 µm.
P1: FQP
AR129-12-COLOR
April 21, 2001
11:57
Annual Reviews
,,
,,
,,
Figure 4 Comparison of FtsZ1 and FtsZ2 proteins from higher plants. Each sequence shown is a consensus derived from several full-length FtsZ1 or
FtsZ2 sequences using the Clustal W program (117) at the Biology WorkBench 3.2 website (http://workbench.sdsc.edu). Sequences used to generate
the FtsZ1 consensus were from Arabidopsis thaliana (U39877), Pisum sativum (Y15383), Tagetes erecta (AF251346) and Nicotiana tabacum
(AJ133453, AJ271749, and AF205858). Sequences used to generate the FtsZ2 consensus were from A. thaliana (AF089738 and CAB89236), G.
lutea (AAF23771), N. tabacum (AJ271750), and Lilium longiflorum (AB042101). The highly variable extreme amino termini are not included in the
alignment. The N-terminal and C-terminal domains are enclosed in boxes. Residues identical in FtsZ1 and FtsZ2 are highlighted in gray. Residues
in FtsZ1 that differ from FtsZ2, and vice versa, are highlighted in green and blue, respectively. Similar residues, underscored by a colon, were as
specified by the default setting of the Boxshade program at the Biology WorkBench 3.2 website. Upper case letters indicate residues that are identical
in all FtsZ1 or FtsZ2 family members indicated above. Lower case letters indicate amino acids that are similar within FtsZ1 or FtsZ2 family, but
not identical. Residues that are not similar and not conserved within protein family are indicated by a dash. Thick black lines above the alignment
show residues known to contact the guanine nucleotide in M. jannaschii FtsZ (53) that are also conserved in the plant FtsZ sequences. Red diamonds
indicate residues essential for GTPase activity in bacterial FtsZ, and a change in one of these specific to FtsZ1 is circled in black (26). Red dots
,,
indicate synergy residues that do not contact GTP, but are required for the regulation of GTP hydrolysis in the bacterial protein. The tubulin
,,
,,
signature motif is underscored by a double black line. The C-terminal core domain sequence (60), specific to FtsZ2, is enclosed by a red box.
Further details are provided in the text.
P1: FQP
AR129-12-COLOR
P1: FQP
April 21, 2001
11:57
Annual Reviews
AR129-12-COLOR
Figure 5 Comparison of structural properties and sequence motifs predicted for FtsZ1
and FtsZ2 proteins from higher plants. (A) Hydropathy profiles of FtsZ1 from Arabidopsis (U39877) (red) and FtsZ2 from Gentiana lutea (AAF23771) (blue), were determined
using the Kyte-Doolittle x-1 method at the Weizmann Institute Bioinformatics website
(http://bioinfo.weizmann.ac.il/). Regions described in the text containing the N-terminal
and C-terminal domains, and the linker region connecting them (L), are separated by vertical black lines. The difference in hydrophobicity from FtsZ2 seen in the helical region H3a
of FtsZ1 also is shown. (B) Alignment between FtsZ1 and FtsZ2 consensus sequences (described in legend to Figure 4) showing predicted motifs and secondary structural features.
Numbers at the right of each line indicate the approximate residue numbers within the FtsZ1
and FtsZ2 proteins. The top two lines in the alignment indicate various phosphorylation
sites predicted for FtsZ1 or FtsZ2 proteins that are conserved within each family. Included
are those recognized by protein kinase C (yellow ovals), cGMP-dependent kinases (red
box), casein kinase II (red diamond), tyrosine-dependent kinase (blue box), and those that
may be phosphorylated by either cGMP-, cAMP-, or calmodulin-dependent kinases (green
boxes). Predictions were made using the programs PPSEARCH (http://expasy.cbr.nrc.ca,
and http://www.ebi.ac.uk), PhosphoBase (44) and NetPhos 2.0 (http://www.cbs.dtu.dk).
Consensus sites were included in the figure only if they were indicated by all three programs, were conserved in all members of the FtsZ1 or FtsZ2 family listed in Figure 4, and
received a NetPhos 2.0 score of at least 0.9. Secondary structural predictions made by
P1: FQP
April 21, 2001
11:57
Annual Reviews
AR129-12-COLOR
Figure 5 Continued
PSIPRED (http://globin.bio.warwick.ac.uk) are shown in the bottom two lines of the alignment. Regions predicted to be non-structured random coils, or loops, are indicated by wavy
black lines; α-helices or β-strands that are conserved in each protein family are indicated by
black or white boxes and labeled H or S, respectively. Hatched boxes indicate two helices,
H3a and H11, that are found only in FtsZ1 or FtsZ2 proteins, respectively. The strands and
helices, more fully described in the text, are numbered according to their relative positions
in the primary sequences of FtsZ1 or FtsZ2.
P1: FQP
April 21, 2001
11:57
Annual Reviews
AR129-12-COLOR
Figure 6 Comparison of hypothetical three-dimensional structures of FtsZ1 and FtsZ2.
Ribbon diagrams of FtsZ1 (top) and FtsZ2 (bottom) were modeled using the program RasMol 2.6 from structural coordinates determined by the Swiss-Model protein modeling server
(34, 85) (http://www.expasy.ch/) using the AtFtsZ1-1 (U39877) and AtFtsZ2-1 (AF089738)
protein sequences, respectively, and the crystal structure of FtsZ from Methanococcus jannaschii (ID1FSZ) (54). α-Helices and β-strands of the N-terminal domain are green and
blue, respectively, and those of the C-terminal domain are orange and red. The N- and
C-terminal ends, and the central helices connecting the two domains (yellow) are labeled
N, C, and H7, respectively; the helix containing the “C-terminal core domain” (60) in the
FtsZ2 structure (light blue) is labeled H11. The Rossmann fold GTP-binding structures in
the N-terminal domains of FtsZ1 and FtsZ2 are composed of alternating β-strands (S1-S6
in Figure 5B) and α-helices (H1-H5 in Figure 5B), which together form parallel b-sheets
(53, 103). The loops in the C-terminal domains contain the “synergy” residues (stars). The
FtsZ2 structure is shown from a slightly different angle than that of FtsZ1 to emphasize the
presence of the C-terminal core domain, which is not found in FtsZ1 proteins. Otherwise,
the structures are nearly superimposable. The figure is further described in the text.