Cell-to-cell signaling is important to both multicellular organims and
advertisement

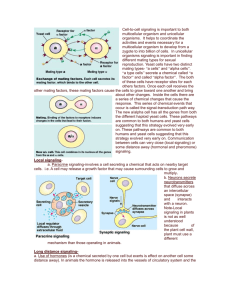
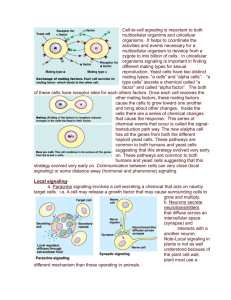
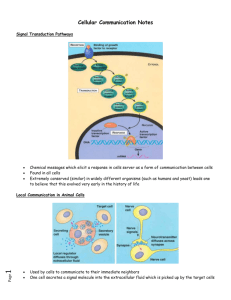
Cell-to-cell signaling is important to both multicellular organims and unicelluar organisms. It helps to coordinate the activities and events necessary for a multicellular organism to develop from a zygote to into billion of cells. In unicellular organisms signaling is important in finding different mating types for sexual reproduction. Yeast cells have two distinct mating types- “a cells” and “alpha cells”. “a type cells” secrete a chemical called “a factor” and called “alpha factor”. The both of these cells have receptor sites for each others factors. Once each cell receives the other mating factors, these mating factors cause the cells to grow toward one another and bring about other changes. Inside the cells there are a series of chemical changes that cause the response. This series of chemical events that occur is called the signal-transduction path way. The new a/alpha cell has all the genes from both the different haploid yeast cells. These pathways are common to both humans and yeast cells suggesting that this strategy evolved very early on. These pathways are common to both humans and yeast cells suggesting that this strategy evolved very early on. Communication between cells can very close (local signaling) or some distance away (hormonal and pheromone) signaling. Local signalinga. Paracine signaling-involves a cell secreting a chemical that acts on nearby target cells. i.e. A cell may release a growth factor that may cause surrounding cells to grow and multiply. b. Neurons secrete neurotransmitters that diffuse across an intercellular space (synapse) and interacts with a another neuron. Note-Local signaling in plants is not as well understood because of the plant cell wall, plant must use a different mechanism than those operating in animals. Long distance signalinga. Use of hormones (is a chemical secreted by one cell but exerts is effect on another cell some distance away). In animals the hormone is released into the vessels of circulatory system and the molecules travel to target cells. In plant cells, the hormones may travel in vessels or diffuse the the air as a gas. Hormones may vary in their size and molecular structure. Types of Animal Hormones Modified amino acids- i.e. epiniphrine is a modified tyrosine. Proteins-i.e. insulin is a large protein Steroid Hormones- i.e. testoterone and estrogen are steroids. b. Pheromones are chemicals secreted by one animal to affect the behavior of another animal.Direct contact- Cells have cell junctions which is in contact of the cytoplasms of adjacent cells occur. This allows for signaling substances in the cytosol to pass feely between the adjacent cells. Animals cells can communicate via direct contact between molecule on their surface of their cell membranes. This is common in embryonic development and the immune system (secretion of interleukin when macrophage and virgin-T cell make a match). The three stages of cell signaling are 1. Reception 2. Transduction 3. Response1. Reception- A large portion of the reception of a signal begins with the signal interacting with a receptor site located on the outside surface of the plasma membrane and will never enter the cell itself. Quite often the signal is a ligand (a small molecule that attaches to a large one). When the ligand attaches it will cause a change in the shape of the receptor site. Receptors are usually proteins inserted into the plasma membrane. Three major types of membrane receptors are a. G-protein-linked receptors b. tyrosine-kinase receptors c. ion-channel receptors a. G-protein-linked receptors vary in their binding sites and for recognizing different G-proteins. Most have seven alpha helices penetrating the plasma membrane. When a G-protein-linked receptor binds with a signal or ligand, it becomes activated and interacts with a G-protein. Yeast mating factors, epinephrine, neurotransmittors and other hormones use G-protein-linked receptors. G-protein-linked receptors activate G proteins. G proteins are also found in the plasma membrane. They are activated when the G-proteinlinked receptors causes GTP to replace GDP on the G-protein. Once this happens the activated Gprotein now moves laterally to interact with a given enzyme also located in the plasma membrane which causes a certain biochemical pathway to be activated. Once this occurs, the Gprotein cleaves a phosphate group from GTP and makes it GDP and thus the G-protein becomes inactivated. These biochemical pathways include developmental pathways, sensory perception in vision and smell. NoteBacteria that often cause disease by secreting toxins. These toxins may interfere with G-protein functions. 60% of all medicines exert their effect by influencing G-protein pathways. b. Tyrosine-Kinase receptors-are receptors that when activated can activate more than one signal-transduction pathway at one time. This is important when an event like cell reproduction requires a number of biochemical pathways to be activated at once. The tyrosine-kinase receptor in the inactive form is two separate polypeptide chains forming alpha helices that span the membrane. Inside the cell, the two polypeptides have tyrosines on tails that extend into the cytoplasm. Both have a receptor site for a signal or ligand. Once both receptors are activated, the polypeptides move toward one another and bond with one another to form a dimer. Now the dimer is phosphorylated at every tyrosine by ATP. Now the receptor is activated and can trigger with a number of transduction-signal pathways. This may include the transfer of a phosphate to another enzyme. Triggering a number of these pathway is the main difference bewteen this receptor and the G-protein-linked receptor.c. Ligand-gated ion channels. These channels are protein pores on the plasma membrane that open or close in response to a chemical signal allowing or preventing the flow of ions into or out of the cell. Examples include Na+ or Ca++. Once the signal attaches to the gated channel, the protein changes shape and a pore opens. This allows for an immediate change in the concentration of ions. d. Intercelluar receptorsSome signal receptors are NOT found in the plasma membrane. Instead they are dissolved in the cytoplasm. This means the signal must be lipid soluble to make into the cytosol. These signals are usually steroids hormones, thyroxine, and nitric oxide. i.e. Testosterone enters the cell and binds with a receptor protein found in the cytoplasm. This complex now moves to the nucleus where is acts a transcription factor and helps transcribes certain genes. Transduction-After reception, the cell needs to process the signal. This is called tranduction and is a mult-step process. Usually the molecules that are processing the signal are proteins that are activated by phophorylating them. Protein kinases are proteins that transfer phosphate groups from ATP to another protein. Serine and threonine of the two amino acids that receive the phosphate group on the receiving protein. A biochemical pathway usually has a cascading effect where one protein kinase will activate another protein kinase by transferring a phosphate group on to it. These protein kinases are extremely important. Approximately 1% of all our genes code for various protein kinases. Protein phosphatase is a protein that will remove a phosphate group from proteins and can deactivate a protein kinase. Between the receptor molecule that is activated and signal-transduction pathway that is to be activated there may be a second messenger. Note-The the hormone or signal that attaches the to the receptor site is called the first messenger. The most common secondary messengers include a. cyclic AMP (cAMP) b. Calcium ions and inositol triphosphate. cAMP-i.e. When epinephrine binds to its receptor site, it is found that there is a corresponding increase in the amound of cAMP found in the cell. cAMP is a derivative of ATP. An enzyme (adenlate cylase) in the plasma membrane causes this reaction to occur. In this example, epinephrine activates a receptor site, which in turns activates the Gprotein. The G protein now activates adenlyate cyclase, which then makes cAMP from ATP. cAMP activates the epinephrine pathway which ultimately increases the amount of glucose in the blood stream. Calcium ions and Inositol Triphosphate (IP3) Some signal molecules in animals induce responses by increasing the amount of Ca++ in the cytosol. This is used in the muscles and nerves and certain hormones. Most of the time Ca++ in the cytosol is low because it is pumped into the E.R., the mitochondria and also outside of the cell. The release of Ca++ from E.R. reservoirs involves diacylglycerol (D.A.G.) and inositol triphosphate (IP3). These two messengers are made by the cleavage of certain kind phospholipid in the plasma membrane. This is done when the G protein acitvates an enzyme phospholipase C. Phospholipase C cleaves a plasma membrane in two to make D.A.G. and IP3. IP3 will then move to the E.R. and bind with a IP3-gated calcium channel to allow Ca++ to flow out. Types of cellular responses 1. Ultimately activating an enzyme 2. Synthesis of a particular enzyme or protein 1. Activating an enzymeUsually when cAMP, Ca++ or IP3 is activated then it will interact with a specific enzyme to start the cascading effect •Scaffolding proteins are large relay proteins to which other relay proteins are attached •Scaffolding proteins can increase the signal transduction efficiency by grouping together different proteins involved in the same pathway Termination of signal •Inactivation mechanisms are an essential aspect of cell signaling •When signal molecules leave the receptor, the receptor reverts to its inactive state