Lecture Notes for Animal Behavior. Part II
1-30-03
Cleavage. Purposes:
1. Increases cell number
2. Decreases individual cell volume. If a cell gets smaller, the ratio of cytoplasm to nucleus goes
down. This has implication for gene action.
3. Allows cells to acquire differences
4. Arranges cells.
As cells divide, they have a modified cell cycle. A typical cell goes through 4 stages: S, G1, G2 and M. In
early embryonic development, G1 and G2 are not there. These are the growth stages. This causes the
reduction of size.
Anytime you look at cell division you have to explain 2 phenomenon:
1. Nuclear division (karyokinesis)
2. Cytoplasmic division (cytokinesis).
-
The mitotic spindle causes the division of the nucleus and the contractile ring causes the
division of the cytoplasm.
Tubulin is the major protein involved in karyokinesis. It makes up microtubules. Actin
and myosin are in the contractile ring.
The proteins for karyokinesis is centrally located and those for cytokinesis is cortical.
Nocodazole is a drug that disrups karyokinesis and cytochalasin B disrupts cytokinesis.
Major Cleavage patterns: Within the embryo we have an animal and a vegetal pole. There is an axis along
the animal to vegetal pole.
1. Complete (holoblastic) vs incomplete (meroblastic). Complete is cleavage all the way through a
cell. Incomplete is partial cleavage. Yolk is the culprit in meroblastic cleavage.
2. If you are cutting top to bottom, you are cutting regular cleavage (parallel/perpendicular). If you
are cutting at an angle, it is spiral (oblique). This is at an angle relative to A-V axis due to the
orientation of mitotic spindle.
3. Another way to look at cleavage is what are the sizes of the daughter products. If the two daughter
cells are the same size, that is equal cleavage. If it isn’t, this is unequal. This is due to displaced
mitotic spindle.
4. This is a specific way to compare the 2 types of regular cleavage. Medidional (parallel) is top to
bottom. Equatorial is perpendicular to A-V axis. This also has to do with the orientation of the
mitotic spindle.
Sea Urchin Cleavage:
Sea urchins are radially symmetric. The first division is a regular cleavage. More specifically meridional.
The mitotic spindles are at a right angle to the cleavage plane. It is also an equal cleavage. The second is
the same. When you go from 4 to 8 cells, you have an equatorial cleavage. Whenever you have an
equatorial cleavage you double the amount of layers. When you cut the bottom tier at the 8-cell stage, it is
en unequal cleavage, forming micromeres and macromeres. Something pushed the mitotic spindle closer to
the vegetal hemisphere so you end up with tiny cells. In the top half we get an equal meridional cleavage
that lead to the formation of mesomeres.
2-4-03
Continuing sea urchin cleavage.
Holoblastic, equal/unequal, regular cleavage patterns. The ball of cells (product of early cleavage) is called
a blastula. Notice that as cleavage occurs, initially there is no growth of daughter cells. The blastula is the
same size of the beginning egg.
Frog Cleavage
In the frog, it is fairly similar. The divisions are regular, equal and unequal. The difference is that the
presense of macromeres is a result of more yolk in the vegetal pole making it harder to divide.
*Be familiar with examples of cleavage and terms*
Chick Cleavage.
In the chick, there is a lot of yolk and a small island of cytoplasm. If you were to look down at the egg, you
would see an ocean of yolk and a little disk that cleaves, leading to a cluster of cells on top of the yolk. This
is called meroblastic cleavage. The resulting cells are about the same size (equal). The cleavage is also
regular. The result of cleavage in the chicken is a disk of cells so it is referred to discoidal cleavage. The
underlying yolk is food for the developing animal. The disk of cells is called the blastoderm. There is a
subgerminal space between the blastoderm and the yolk. There are some of the cells that are in direct
contact with the yolk. The ones that are found above the space are refered to as area pellucida (when you
shine light, this part lights up more). The area opaca is not as clear because there is no space between the
cells and the yolk.
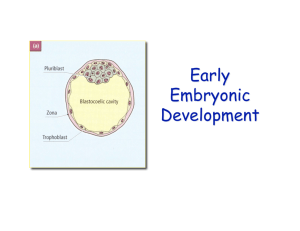
Mammalian Cleavage:
Instead of having a blastula, we have a blastocyst that is analogous to the blastula. As cleavage occurs, the
initial cleavages occur still surrounded by the zona pellucida. Eventually the ZP breaks down. The
blastocyst is a cluster of cells with 2 regions: The inner cell mass, which is surrounded by a hollow sphere
of cells called the trophoblast. The inner cell mass is what the actual animal is going to develop from. The
trophoblast forms a portion of the placenta, which feeds the developing mammal. Part of what happens
during cleavage to generate the blastocyst is the phenomenon of rotational cleavage. This means that the
axis of cleavage of the cell rotates in relationship to the cleavage plane of another cell. In the frog or sea
urchin, the cell divides down the middle so you get 2 cells. Each of these divide down the middle so you
have 4 cells side by side. However, if we were to go to rotational cleavage, one of these planes rotates to 90
degrees. So when the second plane divides you end up with a top layer and a bottom layer as opposed to
just a “left and right” layer. Some of the resulting cells’ cleavage planes also rotate in relation to other cells.
Snail Cleavage:
This is a good example of spiral cleavage. Here we have 2 cells that are side by side and equal in size. Now
we have a spiral division. If you look at cell “A”, you notice that the spindle is tilted. As the cell divides
you get a spiral cleavage. 1a and 1A are a the products of A. Through a series of tilted cleavages you end
up stacking cell layers upon cell layers that are offset. Each new layer that is established adds another
twist. As you do this, you can trace the coiling and eventually you have the left- or right-handed coiled
shell.
Fruitfly Cleavage:
This is an example of meroblastic cleavage. The yolk is evenly distributed. Energids are the nuclei of the
insect egg with the associated cytoplasm. First you have karyokinesis until there are about 6000 nuclei. The
nuclei starts off inside the yolk, then start to migrate, leave the yolk, and enter the cytoplasm. As the nuclei
accumulate, the stage is the development of syncitial blastoderm. This is a multinucleated condition. This is
a temporary condition. As the nuclei leave the yolk and are found only in the cytoplasmic periphery, when
the number reaches 6000, there is a wave of cellularisation dividing the cytoplasm into 6000 cells. This
layer is then called the cellular blastoderm. This is what the fly is built from. This is called superficial
cleavage because only the outermost part of the egg is being cleaved.
Embryonic polarity
How do we build a body plan from a cluster of cells. The first thing we need to explain is how do we
impose an axis of symmetry. The answer to this is that we have a few clues. We will start with the frog as
one example.
Frog
In the frog, the sperm comes in. The male nucleus is meeting up with the female pronucleus. The act of
fertilization causes a shift in the cortical cytoplasm. This rotation occurs towards the point of sperm entry.
The cortical cytoplasm shifts 30 degrees towards that point. Some areas that were clear now have dark
pigment, and some areas that were pigmented are now clear. The gray crescent is formed as a result of this
opposite to where the sperm entered. It turns out that the gray crescent marks the dorsal side. In addition to
knowing that this is the future dorsal side, it seems to be a region that activates proteins that have the power
to direct cell movements within the embryo. This polarization is happening even before cleavage.
Gastrulation is the processes of cell movement that establish the 3 germ layers:
- Ectoderm: Becomes epidermis and nervous system (generally)
- Mesoderm: Becomes muscle, bone, and connective tissue (generally)
- Endoderm: Becomes gut organs and glands (generally)
We have to have an organizer – something to direct all the cell movement. The presence of a primary
organizer does this. This region tells other cells what to do. In the frog, the gray crescent area is that region.
These cells become the dorsal lip of the blastopore. If you transplant the cells into a new embryo, the new
transplant will start to build a secondary body axis forming a twin. The properties of the cells in the dorsal
lip of the blastopore are the conductors.
This phenomenon is shown in other animals also. In the chicken, there is a region called Hensen’s node. If
you graft it onto a new embryo, you end up with 2 animal axes forming from the 1 blastoderm. It
eventually aborts, but the beginnings can be seen. This is a cross species behavior. The transported
Hensen’s node came from a duck.
2-6-03
Types of cell movement during gastrulation: Gastrulation is the orchestrated cell movements into
endoderm, mesoderm and ectoderm. There are certain conductors that organize how cells move. We will
look at 6 ways cells move:
- Surface Movements: Their job is to spread on a surface. They are shaping layers of cells.
1. Epiboly: Epi refers to above. This is seen on the surface of the embryo. A layer of cells flatten out.
They start out cuboidal and flatten out. It is kind of squashing a cell layer.
2. Intercalation: 2 layers interdigitates and become 1 layer.
3. Convergent extensions: We start out with multiple layers that are repacked into fewer layers. This
differs from intercalation because intercalation involves repackaging in a pattern. Convergent
extension is more random.
- Internalization movements: Cells move from an exterior position to in interior position.
4. Invagination: A layer of cells get pushed in to make an impression. You are taking cells from the exterior
and pushing it to the interior.
5. Ingression: The movement of individual cells to an internal position.
6. Involution: Where you have a layer or multiple layers of cells that push in and tuck under the overlying
cells.
Sea Urchin Gastrulation:
The early sea urchin embryo is a single layer of cells with a hollow cavity (blastocoel) on the inside,
surrounded by a hyaline layer. The hyaline layer is kind of like an epithelium. The cell layer lies on a basal
lamina. The first thing that happens during gastrulation is a flattening of the lower half of the embryo
(vegetal pole) to form the vegetal plate. Ingression happens next. The cells that are breaking free are called
the primary mesenchyme cells. These cells come from the vegetal plate. The cells develop different cell
affinities. This cause those cells to be released since they are no longer “attracted” to their neighboring cells
and hyaline layer. Invagination then happens to develop a cavity that is to become the future gut
(archenteron). Invagination starts it, but the elongation is believed to be convergent extension and
intercalation.
Summary:
1. Ingression to establish primary mesenchyme cells.
2. Invagination to initiate development.
3. Intercalation and convergent extension to elongate developing gut.
* The cells from the portion of the vegetal plate that invaginates will repack so that we go from multiple
cells to maybe 8 or 10. *
There are actually two events of ingression. There is the initial PMC ingression. Then there are secondary
mesenchyme cells that ingress from the top of the developing archenteron.
The sea urchin is the simplest form of gastrulation that we are looking at. It only has 1 layer of cells
initially.
Frog gastrulation:
The frog has much more yolk. Any kind of vegetal movement is kind of inhibited by the yolk. Initiation of
cell movement is in the marginal zone (midsection of the embryo). Another factor is that we have multiple
layers to talk about. There are surface and subsurface movements.
The first movement deals with invagination to form the blastopore. As you see, when you push in, it starts
to deform some cells. These elongated cells are called bottle cells. These are some of the first evidences of
cell movement inside the embryo. Involution then happens. Cells that use to be on the surface start to move
under the surface cells. These are future mesoderm cells. During this movement, we are now creating the
archenteron. The way we built archenterons in the frog involves both invagination and involution. This
displaces cells inside and the blastocoel gets smaller and smaller until it disappears. The bottle cells are
some of the first to get pushed ahead and become part of the embryo that lines the gut. At this point in frog
gastrulation we have basically set up the 3 germ layers. The blastopore takes up the appearance of a plug
(yolk plug). These cells are not moving. Epiboly helps spread cells across the surface.
Early movements:
There is an initial dimple at the marginal zone that deforms cells. Cells (surface and subsurface) curve
under and involute (bottle cells). Involution is basically the major way in which frog cells become
internalized.
Chick Gastulation:
The chicken has a lot of yolk that prevents involution. Ingression is the major movement. So we have a
single layer of cells that is called the blastoderm. Cells break free from this layer and enter the subgerminal
cavity. They are joined by cells from the area opaca (Koller sickle region). They divide and form a new
layer called the hypoblast. This layer will eventually seal off a new cavity called the blastocoel. The top
layer is then called the epiblast and the chicken is built from this layer. The hypoblast cells form different
things important to the development of the chicken. The next events happen in the epiblast. As cells
converge through convergent extension, the thickening of the region of the blastoderm (Koller’s sickle) that
thickens and grows out to form the developing body axis. It stretches out until you have an elongated line.
This region is called the primitive streak and is develops before the chicken lays the egg. If you look
closely at this streak you see a groove called the primitive groove. The very end of it is the Hensen’s node
(organizer). The primitive streak is a temporary structure that forms to allow cells to become internalized.
Once the primitive streak is formed a variety of process start to happen at the same time. As time goes on,
the primitive streak shrinks. As it shrinks, different things develop where it was (notochord somites etc). As
the primitive streak shortens, the notochord lengthens.
Cells converge at the streak. These cells move at this primitive groove. Cells can dip subsurface and enter
the blastocoel. As they move in, they establish the 3 germ layers. Some of them move to the very bottom
and displace the hypoblast cells and set up the endoderm. Others don’t move as deep and set up the
mesoderm (by ingression). The cells that do not ingress are going to develop into the ectoderm. The basic
way we set up the 3 germ layers in the chicken is ingression. The primitive streak is really just a temporary
structure that allows the cells to move. No involution or invagination happens in chick gastrulation. The
only other type of movement that seems to play is epiboly of the epiblast.
Missing:
- 2-11-03 (Get from someone)
2-13-03
Sometimes when we classify animals, we use parts of their body that appear in development. There is a
group called the chordates. These are called chordates because they have a notochord that is involved in the
development of the body plan. There is another group called the amniotes. These posses a structure in
development called the amnion. We are amniotes. Birds and reptiles also fall into this category. What we
are looking at now is what is the amnion. This will be compared between the chicken and the human.
Chicken
Extraembryonic membranes form as extension of the embryonic germ layers. In the early embryo the germ
layers are on top of the yolk. These extraembryonic germ layers. The layers split into a somatic layer and a
splanchnic layer separated by extraembryonic coelom. These membranes enclose things.
The way the embryo gets enclosed is through folding of the extraembryonic germ layers. One is called a
head fold and there is a tail fold. These folds thrust up, grow higher and higher and the two tips fuse
making a roof that encloses the animal. These 2 layers that are fusing are the ectoderm and the somatic
mesoderm (keep track of these). While this is happening the yolk gets enclosed also. Endoderm starts to go
around it, and the mesoderm follows. This is called the yolk sac, which is made of inner endoderm and
splanchnic mesoderm. As these membranes fuse a multimembrane based system is made outside of the
membrane.
When you take the endoderm and the splanchnic mesoderm together, they are called the splanchnopleure.
The counterpart that involves ectoderm and somatic mesoderm is called the somatopleure.
The outermost membrane that form is called the chorion and is made up of somatopleure.
The amnion is also built from somatupleure. The Yolk sac is made of splanchnopleure. The outgrowth from
the hindgut is the allantoic membrane and is made of splanchnopleure.
The amnion encloses the amniotic cavity. The yolk is enclosed by the yolk sac. The chorion encloses the
extraembryonic coelom, the allantois encloses the allantoic membrane.
If you go through the whole thing, you go through the chorion (first ectoderm, then mesoderm), then the
extraembryonic coelom, then you go through the allantoic membrane, which is made up of mesoderm and
endoderm. You then pass through the allantoic cavity, then through endoderm, mesoderm, extraembrionic
coelom, mesoderm, ectoderm, amniotic cavity.
So you start out with ectoderm and you end with ectoderm.
As the chicken grows, the yolk sac shrinks as the yolk is used up, the animal expands and eventually the
extraembryonic coelom is gone. The allantois actually fuses with the overlying chorion.
Why do we have the 4 layers:
1. The yolk has a nutritional role.
2. The chorion is important for respiratory exchange.
3. The allantois stores waste that is secreted. Where it fuses with the chorion it also serves as
respiratory exchange.
4. The amnion protects the embryo (fluid filled).
Human placenta:
The placenta is made up of extraembryonic membranes. To trace back where it comes from we have to
look at the structure of the blastocyst.
The trophoblast gives support to the developing embryo. Implantation starts at around day 6. The blastocyst
moves to a sub surface position. The blastocyst gets swallowed up and the cells of the trophoblast
proliferate and invade deeper and deeper into the endometrium. This invading part is called the
syncytiotrophoblast. They will eventually hit capillaries and the blood will still out as the wall of the
capillaries are broken down. The blood will bathe the cells of the syncytiotrophoblast. Any nutrients in the
blood can be taken up. As time goes on, the syncytiotrophoblast set up blood pools called lacunae where
the blood is enclosed by the tissue. This blood interaction is a primitive “placenta”. Implantation has
happened. The second evidence of developing placenta is found in cells in the trophoblast that form
finderlike extension and reach out to the pools of blood and forms a chorionic villus. The cells are called
the chorion cells. The job of these cells are to selectively proliferate in rays that form an interface between
the fetal and maternal circulatory system. They house the fetal circulation but will be bathe by the mom’s
blood so nutrients can move along with antibodies etc. There is a barrier though (placental barrier) so that
the blood doesn’t mix. Three weeks into development, the chorionic vesicle (made up of chorionic villi) is
formed. In the mature placenta, capillaries fill the chorionic vesicle. The placenta really has two parts:
1. The chorionic villi (developed by the fetus)
2. The part from mom that makes the room.
Implantation occurs on the back side of the uterus in the upper half. In the woman, the endometrium is
called the deciduas. This is going to be shed during the birth experience. As the baby develops, you can see
3 regions of the deciduas:
1. The part that is overlying the child is called the decidua capsularis.
2. The part that is underneath the child is called the decidua basalis
3. Everything else that is not directly touching the child is the decidua parietalis.
When the baby gets bigger and bigger, the capsularis layer fuses with the parietalis layer so that there is no
more cavity. As this happens, the pressure of the baby pushing out crushes the chorionic villi and they die.
They take on a smooth region called the chorion laeve. Just the opposite is happening on the other side of
the baby. The chorionic villi are proliferating and is called the chorion frondosum and are part of the
placenta. It interfaces with the decidua basalis from the mother and form the placenta. The fetal component
is the chorion frondosum, the mom’s component is the decidua basalis. When the baby is born, you have to
rupture the chorion. The bag of water breaks and the baby comes out. The placenta will not come out until
you cut the chord. Then the whole plate of chorionic villi and decidua basalis will then come out.
If not all the placenta comes out, the piece can interfere with appropriate contraction to deliver the placenta
and cause a lot of excess bleeding.
The Cellular Basis for Morphogenesis:
Morphogenesis is the group of process that builds the body plan. It’s the shaping of the embryo. It is going
to involve a lot of different things. We are going to look at it from a cellular point of view. The emphasis
here is on the cytoskeleton.
Elements of the cytoskeleton:
1. Microtubules: These have a diameter of 24 nm. They are build from a polymer of dimers called
tubulin. In most of our cells, microtubules play movement type of roles. This is the thickest of the
filaments.
2. Microfilaments: 8nm in diameter. These are polymers of G-actin. It is polymerized in a twisted
helix of two chains of the actin to make the filaments. This is the thinnest of the filaments. You
can distinguish an endedness. There is a + end, which grows the fastest during polymerization and
a –end. Actin is important in contraction.
3. Intermediate filaments: They are 10nm in diameter. 4 protofibrils make up an intermediate
filaments. In each protofibrils, we have protofilaments. These are believed to play a role in
mechanical strength of cells. It has been shown that people who take anabolic sterioids, levels of
intermediate filaments increase in their cells. There are different kinds of intermediate filaments in
different kinds of tissue:
a. Lamins: Make up the nuclear lamina
b. Neurofilaments: found in neurons
c. Vimentin: found in bone/cartilage
d. Keratins: found in epithelial layers.
Intermediate filaments have a structural role while actin tends to have a functional role (contraction etc.)
Cellular movement as epithelia.
There are various types of epithelia. In this example we are looking at simple columnar cell with microvilli
extending from them. This increases the surface layer. In epithelia these cells are fused through a junctional
complex.
1. Zonula occludens (tight junction): The membranes fuse at the apex of the cell. It keeps stuff from
getting in between cells.
2. Zonula adherens (adh. Junction): Associate with a band of microfilaments and runs around the
whole top of the cells.
3. Macula adherens (desmosome): On the wall of the cell membrane, there are spots where there are
attatchment. There is crosslinking between desmosomes with keratin. Knockout mice for keratin
could not make a particular keratin proteins. These mice looked fine, but died a few days later
from lack of eating. They look at their tongues and saw that they were all blistered. Apparently
without keratin, when these mice tried to nurse, just the act of nursing caused blisters, which were
so painful that the tongues did not feed.
2-18-03
Cellular movement as Epithelia.
Example: In the process of neurulation, we are dealing with the surface layer of ectoderm. There is a
contraction of the actin microfilaments in the junctional complex that is involved in the bending and
shaping the ectoderm. We believe that these microfilaments contract and the apical squishing of the cell
causes the layer to bend. The top end of the shells shrink in unison and the bottom doesn’t. There is an
assymetrical shaping of the cell layer. If you apply this to the ectoderm, looking at the neural plate. The
cells of the neural plate thickens due to microtubules elongating. Then the apical part contracts, causing the
bending and folding. This is an example of cells moving together as a layer. The whole movement is just
changing the form of where the cells are in relationship with each other, going from a flat layer to a circular
layer.
Individual cell movement:
e.g. the neural crest cells. How do cells move, what drives their movement etc. Here we are looking at
fibroblast cells. Fibroblasts are derived from connective tissue. As they move, they crawl along the surface
of the bottom of the dish and take up an interesting cell shape. They have a broaden flat extension an is the
leading edge. The back is the trailing edge. The long extension is called the lamellopodium. The leading
edge actually sticks out (lamellopodium). It sticks out farther and farther. This is due to the extension of
actin. If you stain the cell with antibodies for actin, you can see strong amounts of actin in the leading edge.
It’s kind of like a slinky. The front end stretches out and then the back end lets go of where it was
anchored. There are cycles of extension and retraction.
Fibroblast motility:
A fibroblast moves very slowly. Sometimes, pieces of the cell will break off and leaves part of the trailing
end behind.
What causes the leading edge to extend? When you look at a moving cell, the front end curious crosslinking of actin microfilaments. At some branches you can make out a protein complex. The actin
microfilaments are pushing against the leading edge. The current model is called the dendritic nucleation
model.
The Dendritic Nucleation Model:
Trying to explain how the leading edge moves. Dendritic refers to a tree and tries to explain the branching
filaments of the microfilaments. Nucleation tries to explain how the filaments are formed. There are 4 key
players (outside of actin) that need to be understood (FTQ 2):
1. WASP: This is a family of protein. One particular one is called Scar. Something (maybe a growth
factor) activates the Scar protein.
2. When WASP/Scar is active, it activates the Arp2/3 complex. It stands for actin related protein and
is involved in nucleation and branching. When it is active, it starts the polymerization of actin.
3.
4.
However actin has to have a starting point. The Arp2/3 complex binds to an actin monomer. It
acts as a starting point for polymerization. One of the ways we get microfilaments de novo is
through the activity of the Arp2/3 complex. The other action of arp2/3 is to build branches on
preexisting microfilaments. It binds to it and acts as a branch point. We explain the dendritic part
of the model by Arp2/3 binding. These branches are always 70 degrees.
Profolin: Its job is to deliver g-actin to the arp complex. When profolin binds the free actin, it can
deliver it to where it’s needed. When the actin binds to make the filament, ATP is part of the
molecule. Part of the charging of the actin (with ATP) is the job of profolin. ADP actin is not
ready to be used. There is an ADP and ATP exchange. Profolin has a supportive role.
Cofilin: This is a severing protein. It breaks the actin microfilaments making pieces of
microfilaments. If you inhibit cofilin, polymerization is compromised. There is something about
the severing that is necessary for polymerization. There is an endedness to microfilaments (+ and ). Globular actin can only be added to the plus end. When cofolin comes along and chops it up, it
creates a new end that can be used for polimerization. When you chop a microfilament in 2, you
create a new plus end, which becomes a new addition site for monomers. Then maybe Arp can
come along and reattach them. Cofilin creates new barb ends where monomers can be added.
In Vivo cell motility:
Sea urchin secondary mesenchyme cells exibit filopodia. These are long threadlike extensions. Fundulus
deep cells exibit lobopodia and filopodia. The deep cells are cells that are moving across a big body of
yolk. If you look up close to them, you see that they also have long slender extensions (filopodia) and long
sausage-shaped extension (lobopodia). So there are lamelopodia, filopodia and lobopodia, which are three
ways of moving individual cells.
Axon Growth Cones
How do cells know where to go? Do they have directional cues? We are going to look at neurons. They
work by forming synapses with other cells. They have to find a partner in order to create this connection.
The cytoskeleton seems to be involved in movement. At the end of an axon in a growing neuron you have a
growth cone that is moving and looking for partners by sensing chemicals. It has filopodia. If you look at a
moving growth cone, and put a repellent in the area, you notice that as the growth cone senses the repellent
there is a turning. It starts to move away from the repellent. The question is: why is that? First of all we
notice that the lamellopodia collapses on the side closest to the repellent. The question is: Why does it cave
in on one side? If you look at the cytoskeleton you see that there are microtubules that extend along the
axon (and into the cone) and microfilaments in the cone. The cytoskeleton gets remodeled in response to
the repellent. When they looked inside the turning growth cone, they noticed that there was a changing in
the cytoskeleton. The bundles of microfilaments gets disorganized in the collapsing part. The microtubules
also collapse. So there is a loss of microfilaments and a loss of microtubules in the collapsing end
(asymmetrically). With this loss, the whole side collapses. Where the cytoskeleton is working is the side
opposite the repellent and you end up with a turning of the cell.
2-20-03
Axon growth cone guidance:
How the Robo code helps commissural axons navigate the embryo. And axon can get very long and its job
is to find a partner and connect. The growth cone is the end that’s growing. In this model system in the
fruitfly, some axons cross from one side of the body to the other. These are called commissural axons
because they cross over. The question is: Why would the embryos crossover and what tells them where to
form a pathway of growth? On the growth cone, there are cell-surface proteins expressed that are receptors
for different signals. The main one is called robo, which is short for roundabout.
If you look at a stain of the motor axons, you see the midline and a bilateral symmetry. You see 3 bundles
on either side of the midline. Each bundle is called a fascicle. The route that is closest to the midline is
called medional, the next is intermediate and then the lateral bundle. Some axons are commissural. The
question is: What determines if an axon crossover and how does it know which bundle/fascicle to join?
It has to do with Robo and what it’s sensing. The 1st step is that the commissural axon has to be attracted to
the midline of the animal. This midline is a source of chemical cues that are expressed by the cell and
diffused out. The first cue is netrin. Cells that have the receptor for netrin are attracted to it. The receptor is
DCC. It binds the netrin and grows closer and closer to the source of netrin. As it starts to cross the midline,
it stops expressing robo. Once it crosses, it upregulated expression of robo. Robo senses a reppelent called
slit, which is released at the midline. During the crossing, the ability to sense slit is not there. Once it’s
crossed robo is expressed and it is repelled by the midline’s slit (DCC is also silenced). We don’t want the
axon to cross back over.
It has been shown that in robo mutants, it doesn’t sense the inhibitor and goes back across the midline
(roundabout mutants). With a slit mutant, the same thing happens.
So, robo is helping keep the axon on the correct side of the body. The next question is how does it know
which fascicle to join. The answer to this question was answered about 2 years ago in Vienna. The answer
is in the form of a robo code. They noticed that there is more than one kind of robo receptors (3) for slit.
Growth cones have 3 ways of sensing the same repellent: robo, robo 2 and robo 3. First they stained the
developing NS with antibodies against the 3 different robo. They noticed that robo 2 is expressed farthest
from the midline in the thinnest pathway. Robo 3 is expressed closer to the midline and overlaps with robo
3. Robo has the thickest expression closest to the midline along with robo 3 and robo 2. So lateral pathways
express all 3 robos. The ability to sense an inhibitor should be proportional to the amount of receptors you
have. The more receptors you have for slit, the more you are going to be repelled. The axons that take the
lateral position should be, and are, more sensitive to slit because they are more sensitive to it (3 robos).
Those that are medial will be least sensitive since it only expresses 1 kind of robo and will not be repelled
at much. The robo code is the combination of what particular types of robo is being expressed by a cell.
This is the theory behind why we think certain axons choose certain routes.
To test this, they did a loss of function test by knocking out the robo 2 or 3 gene. The prediction is now that
axons that use to express it should be more tolerant meaning that they can choose a pathway closer to the
slit repellent. This is exactly what they found. There was a displacement of axons closer to the slit. Axons
that were lateral now take up a medial position.
The other test is a gain of function mutation where you add extra amounts of a function genetically. They
did that with a particular axon that takes up a medional pathway. They caused it to express robo2. The
result was that it moved away from the midline. If you caused more expression of robo (eg robo3), they
moved even more away (lateral).
So the idea is that there is a gradient of slit. Where the axon picks depends on the mount of robo being
expressed. Loss of function moves axons closer to the midline and gain of function moves them away.
Last year, the same lab found out what control robo. It is called the comm. Gene. They showed that the
action of comm is to send robo proteins to the lysosome to break it down. A cell was given the robo gene
and it was showed that it is expressed everywhere. They then gave the comm gene and they saw that they
were localized in the endosomal pathway. When they added comm robo was no longer expressed
everywhere, but only where comm is. It has been shown that comm directs robo to the lysosomes in this
way and also ubiquitinates robo, marking it for destruction.
The turning off of robo expression happens when the comm gene is on in addition to the robo gene.
However, since comm is expressed, robo gets broken down. When the axons cross over, the comm gene is
turned off, and robo is then routed to a cell-surface position and can then sense slit. Ipsilateral neurons
(stays on 1 side) never turn off comm so they can never cross.
A fly that lacks the comm gene doesn’t have commissural axons. Comm is short for commissureless.
Cell adhesion in Morphogenesis.
Cells position themselves in the embryo to form the body. The ability to bind to stuff is important in this
process.
1. Adhesion to the ECM (what is it):
a. Collagen: the most abundant ECM molecule in mammals
b. Proteoglycans: such as chondroitin sulfate, hyaluronic acid
c. Fibronectin: crosslinks the ECM
d. Lamin: in basal lamina.
On the surface of cells, they have cell-surface receptors that mediate this adhesion. Sticking out of the cell
is the integrin protein, which is a dimer with an alpha and beta subunit. The alpha subunit is involved with
binding calcium. There is also a C and N terminus. The N terminus binds to an RGD (arg-gly-asp)
sequence on the fibronectin. There are also other receptors that do this sort of thing. Integrin alo binds to
the internal architecture of the cells. Fibronectin has been shown to be along migratory pathways.
2.
Adhesion to other cells. Cells can bind to each other through 2 kinds of molecules
a. Cadherins: mediate most vertebrate calcium-dependent cello-cell binding. They are
located on the cell surface. There is an N terminus outside of the cell and a C terminus
inside of the cell. Just like integrin, the inside is interacting with the actin cytockeleton.
There are proteins called catenins that mediate this connection. Cells express cadherins
on their cell surface and can bind to other cells with cadherins. Cadherins are a family.
One kind can only bind to the same kind. In neurulation in the frog, it has been shown
that as ectoderm is undergoing neurulation they start expressing different kinds of
cadherins. Those that express E-cadherin become epidermis. Those that express Ncadherin becomes neural tube. It’s been shown that if you mess up the expression, it
messes up the whole neurulation process.
b. N-Cams: calcium independent.
Copyright © 2003, Leslie Samuel Productions.
All Rights Reserved