Annual Reviews
www.annualreviews.org/aronline
Copyright1972. All rights reserved
THE OPERON REVISITED
Annu. Rev. Genet. 1972.6:133-156. Downloaded from arjournals.annualreviews.org
by University of Wisconsin - Madison on 03/29/07. For personal use only.
WILLIAM
S. REZNIKOFF
Departmentof Biochemistry,Collegeof Agricultural and Life Sciences,
University of Wisconsin, Madison,Wisconsin
INTRODUCTION
Simple observation of living organisms allows one to conclude that the entire
genetic potential of any given cell is never expressed at any given moment.In
bacteria this phenomenonis termed genetic adaptation and it occurs in two apparently contradictory ways. The synthesis of enzymesresponsible for a catabolic
process generally appears to be turned on (induced) by the substrate (1) while
formation of anabolic enzymes appears to be turned off (repressed) by the end
product of each given pathway (2). These observations have led to the question-what mechanismsare responsible for genetic adaptation ?
In 1961 Francois Jacob and Jacques Monodaddressed themselves to this
problem and, in an attempt to describe the mechanisms of genetic adaptation
(and hopefully howgene regulation occurs in all organisms), they presented the
"operon model" (3, 4). This model in fact described two phenomena: (a)
structural genes (those coding for proteins) express themselves, and (b) how
expression is regulated.
Some parts of the operon model have been so consistent with experimental
results that they have becomearticles of faith for most biologists. These will be
described only briefly. Other aspects of the model have either had to be modified
with time or have been found to apply to only a few systems. These eases will be
discussed in greater detail. This review will be restricted almost entirely to
Escherichia coli and Salmonella typhimurium. The reader is strongly advised to
search out other recent reviews (5-17).
THE EXPRESSIONOF STRUCTURAL
(~ENES
Summaryofmodel.--Structural genes function by serving as templates for the
synthesis of messenger RNA(mRNA).It is the mRNA
that, in association with
the protein synthesizing machinery (the ribosomes, translation factors, aminoacylatcd transfer RNAs,etc.), determines the primary structure of proteins. This
mRNA
is degraded soon after its synthesis. Structural genes coding for the
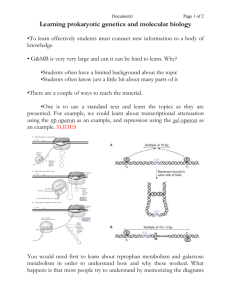
proteins of a particular biochemical pathway are often clustered on the chromosome(e.g., the genes controlling lactose (lac) utilization shown in Fig. 1).
Such a group of genes is expressed as a unit in a sequential polarized fashion.
The synthesis of mRNA
initiates at one end of the operon and progresses through
the structural genes (lac z, lac y, and laca in the case of lac) to the end of the
133
3038
Annual Reviews
www.annualreviews.org/aronline
1.34
REZNIKOFF
Annu. Rev. Genet. 1972.6:133-156. Downloaded from arjournals.annualreviews.org
by University of Wisconsin - Madison on 03/29/07. For personal use only.
p
I~I
i
t:po
I~_|.I I
y
a
FIG.1. Thelac regionof the E. coli chromosome.
Thisis composed
of twounits of
transcription,the lae i gene,whichcodesfor the lae repressor(REP),andthe lac operon
whichincludesthree genes,lae z, lac y, andlae a, codingfor/%galactosidase
(B-Gz),
galactosidepermease(B-P) and thioglactosidetransacetylase (TA)respectively.
polymerase(RNP)binds to and initiates mRNA
synthesis at the promoter, p. The
RNP-p
interaction for the lac operonis dependenton the presenceof the catabolite
geneactivator protein (CAP)and cyclic AMP
(A). Transcriptioncontinuesuntil
RNPreachesthe terminator (0. TheREPpreventstranscription of the lac operonby
bindingto the operator, o. TheREP-ointeraction is preventedby the inducer(D.
operon. Such a unit of transcription was defined as an "operon" by Jacob &
Monod(3, 4).
The messenger, unstable RNA.--By1961 it was believed (a) that proteins
were not produceddirectly on the chromosome
but rather on the ribosomes, (b)
that RNAwas an intermediate "messenger"template in protein synthesis, (c)
that ribosomesdid not provide the necessary specific messengerinformation,
and (d) that there existed a metabolically unstable species of RNA,whichcould
be found associated with the ribosomes,whoseproperties moreaccurately fitted
the expected properties of "messenger-RNA"
(3, 4, 18-20). Subsequentlyit was
discoveredthat specific inhibitors of RNA
synthesis wouldpromptlyblock protein synthesis (21-23) and that the unstable RNA
contained species that could
form specific molecularhybrids with DNA
knownto be active in the ceils from
whichthe RNA
was isolated (24, 25).
An important question is whether all mRNA
species are equally unstable
and, ff not, what factor(s) determine the degree of mRNA
instability. Some
mRNA
species in E. coli appear to be quite stable. Analmost trivial exampleis
the RNAmolecule injected into E. coli by f2 and other RNAphages. This
moleculeis both the chromosome
and the messengerof the phage (26). Another
exampleis the RNA
codedfor by the coliphageT7 (27). The relative stability
T’/mRNA
is, at least in part, an inherent property of the mRNA
itself. Measurements of the stability of tryptophan (trp) mRNA
and T7 mRNA
in T7 infected
cells indicate that the two species of mRNA
maintaintheir distinctly different
half lives (98). WhetherT7 in addition produces a regulatory agent which
"stabilizes" certain mRNA
species (amongwhichtrp mRblAis not represented)
is still an open question. The original studies by Summers
suggestedthat bulk
Annual Reviews
www.annualreviews.org/aronline
THE OPERON REVISITED
13 5
Annu. Rev. Genet. 1972.6:133-156. Downloaded from arjournals.annualreviews.org
by University of Wisconsin - Madison on 03/29/07. For personal use only.
E. coli mRNA
is more stable after T7 infection (27) and, in addition, T4 early
mRNA
seems to be more stable in T7 infected cells (R. Haselkorn, personal
communication).
The operon,functional polarized grouping of structural genes.--Jacob & Monod
(3, 4) were struck by an observation made by Demerec(29) that genes coding
biochemically related functions are frequently linked together on both the E. coli
and S. typhimurium chromosomes.They postulated that this linkage has a functional aspect--the linked genes are expressed sequentially as a group via the
synthesis of polygenic mRNA.The experimental evidence that supports this
hypothesis includes: (a) The observation that the genes of the histidine (his)
operon (and other ope~ons) are expressed coordinately (i.e., the repression/induction ratios of all the enzymescoded for by the operon are identical) (30), (b)
existence of polar mutations (mutations in one gene which decrease the expression
of neighboring genes; these effects are always manifested in a unidirectional
manner, e.g. lac z--~lae a) (31L34), (c) The existence of regulatory mutations
located at one end of the gene group in the operator region. These mutations
affect the expression of all the linked genes (3, 4, 35), (d) Theisolation of pleiotropic transcription defective mutations in the promoter region located at the
same end of the operon as the operator (36), (e) The detection of polygenic
mRNA
corresponding to the lac operon (37, 38), the his operon (39), and the
trp operon (4,0), and (f) The observation that induction and repression
quentially affect both the synthesis of the enzymescoded for by an operon (41)
and the synthesis of the RNAmessenger (40, 42-44).
In some cases the genes corresponding to a given pathway are not linked.
Somepeople feel that this observation conflicts with the operon model. In fact,
Jacob & Monodrecognized and accounted for these cases by suggesting that in
such a system there may exist multiple operons each composed of one or more
structural genes and that all related operons could be controlled by one regulatory gene in a similar but not necessarily coordinant fashion. The arginine (arg)
system (45-47) was the example used by Jacob & Monod(3, 4). Maas & Clark
(48) coined the term "regulon" to describe any system regulated by a single
regulatory gene regardless of how manyoperons are involved. Twoother regulons
that are composedof more than one operon are the arabinose (ara) system (132)
and the trp system (164).
There are special conditions where the synthesis of the enzymescoded for by
the his operon occurs in a simultaneous rather than sequential fashion (49, 50).
These conditions are those that produce high intracellular
concentrations of
Nl°-formyltetrahydrofolate. The mechanistic basis of these observations is unknownbut they may represent an exception to the generally held hypothesis of
sequential polarized expression.
The promoter, transcription initiation ~ site.--The expression of an operon via
the polarized, sequential synthesis of polygenic mRNA
implies a specific genetic
t Theterm "transcription initiation" refers to all the steps prerequisite to the formation of the first mRNA
phosphodlester bond.
Annual Reviews
www.annualreviews.org/aronline
Annu. Rev. Genet. 1972.6:133-156. Downloaded from arjournals.annualreviews.org
by University of Wisconsin - Madison on 03/29/07. For personal use only.
13 6
REZNIKOFF
region associated with each operonwheretranscription initiation occurs. In the
original operonmodelthis region wasdefinedas identical to the repressor binding site--the operator (3, 4). Subsequently,whenmutationsin the operator region
werediscoverednot to havedrastic effects on the rate of maximallac expression,
the transcription initiation site waspostulatedto be a distinct geneticentity--the
promoter (51, 52). Defim’tionof the promoter region awaited the isolation
four mutations in this region by Scaife & Beckwith(36). Thesefour promoter
mutationshave these importantcharacteristics: (a) Theyare pleiotropically lac
negativein a cis-dominantfashion (linked lac z, lacy and lae a expressionare all
coordinately depressed) (36). (b) Theyare not suppressible by any known
sense or polarity suppressors(36). (c) Theyare (at least qualitatively) + (they
do not manifesta defect in repressibility) (36, 53, 54). (d) In at least onecase
lac i-lac p deletion L1) a promotermutationdecreases in vitro lac transcription
in a purified RNApolymerase-lac DNA
system (55).
Geneticlocalization of these promotermutationsresulted fromthe isolation
of lac deletions entering into or near the lac promoter region from both the
"’lac/-side" and the "lac z-side." The generation of these deletions was made
possible by the isolation of twoclasses trek80 dlac prophages(56). In both cases
the lac region carried by the prophageis located dose to the tonB locus so that
selection for resistance to bacteriophageT1 can yield lac deletions (57). Most
the tonB---lae p deletions werechosenfromthe total collection of tonB-’-lacmutations by a very arduous procedure requiring the genetic screening of all
possible candidates(53, 54, 58). Usingthese mutations,it waspossible to locate
the promoterbetweenlac i and lac o as indicated in Figure 1 (53, 54).
Anotherapproachto mappingthe promoter,vis-a-vis the operator, is to ask
whetherit is possibleto isolate lac o-lac z deletionsthat do not drastically alter
maximalexpression of the/~c operon. Suchdeletions have been found using a
techniquefirst describedby Jacobet al (51, 59). Oneof these deletionsresults in
substantial inactivation of the operator while only reducing the maximalexpression of the lac operon12%.This result indicates that the genetic material
missingin the lac o-lac z deletion S145is not necessaryfor the initiation of lac
transcription. Furthermore,in vitro experiments,to be describedlater, suggest
that at least one of the steps involvedin transcription initiation, the bindingof
the polymerase,can occurin the presenceof the lae repressor (60).
A similar delineation of the p and o regions has been found for the trp
operon(61, 62) although in other systems such as his the distinction is not so
clear (63). Theoperator and promoterare in the reverse order vis-a-vis the structural genesin the ara operon(9).
In spite of these straightforwardresults, one mightsuspect that this description of the promoteris too superficial. Transcriptioninitiation is probablya series
of sequential reactions (i.e., RNApolymerasebinding, RNApolymerasemovement, mRNA
chain initiation) each of which maywell require separate DNA
sequencesignals (64, 65). Furthermore,operator mutations do alter the maximal
level of expressionof the lac operon(66) althoughit is not dear that these are
Annual Reviews
www.annualreviews.org/aronline
Annu. Rev. Genet. 1972.6:133-156. Downloaded from arjournals.annualreviews.org
by University of Wisconsin - Madison on 03/29/07. For personal use only.
THE OPERON REVISITED
137
transcriptional
as opposed to translational
effects. Moreover, the promoter
mutants are not quantitatively as inducible as the parent wild type strain (54).
A further complication in our understanding of the lac p region arises from
the fact that this promoter is recognized efficiently by RNApolymerase only in
the presence of the catabolite gene activator protein [CAP;in other articles it is
termed CGAprotein (67) and CRP (68)], and adenosine 3’-5’ cyclic monophosphate (cyclic AMP).The expression of the lac operon, and other catabolic
systems, is depressed when the ceils are grown in the presence of glucose or
glucose-6-PO4 as opposed to glycerol. This phenomenonis called catabolite
repression. That the promoteris the target site for eatabolite repression is indicated by the work of Silverstone et al (69), Pedmanet al (70) and Beckwith et
(71). These groups have shownthat lac promoter mutations result in a loss of/ac
catabolite repression sensitivity. Furthermore, high level expression of lac that is
insensitive to catabolite repression can result either from fusing the/ac genes to a
catabolite insensitive promoter system (such as trp) (69) or by generating second
site, closely linked, revertants of promoter point mutations (72). Since the
lac Macp deletion results in total insensitivity to catabolite repression and since
the catabolite repression insensitive revertants of promoter mutations map outside of L1 (towards lac z), the region of the promoterinvolved in catabolite repression is covered by L1 (71).
The identification
of cyclic AMPand CAPas the active components in
catabolite repression came from studies showing that the intracellular level of
cyclic AMP
is lowered whencells are subjected to catabolite repression (73), that
exogenous addition of cyclic AMPreverses catabolite repression (74, 75), and
that mutants unable to express any catabolite sensitive operon are defective either
in adenyl cyclase or CAP(76-78). The hypothesis that CAPand cyclic AMPare
necessary for the initiation of mRNA
synthesis at lacp has been supported by the
observation that CAPand cyclic AMPare absolutely required for correct in
vitro synthesis of lac mRNA
in the purified RNApolymerase, lac DNAsystem
(55, 60, 79). One would suspect that CAPwould show a cyclic AMPdependent
affinity for lac p DNA,since it does not show an affinity for RNApolymerase
(79) and since a mutation in CAPcan phenotypically revert la c pr omoter
mutation (Plenge & Beckwith, personal communication). Unfortunately CAP
shows a cyclic AMPstimulated affinity for any DNA(67).
Possible promoter mutations decreasing gene activity have been isolated in
the his operon (80), the leucine (leu) operon (81), and the trp operon (61) while
promoter-like mutations increasing gene activity have been discovered associated
with the lac i gene (82) and the gene coding for glucose 6-phosphate dehydrogenase (83).
The terminator, mRNAtermination site.--While not mentioned in the original
operon hypothesis (3, 4), the synthesis of specific polygenic operon mRNA
clearly implies a specific mRNA
termination site at the end of each operon.
The lac i gene has the same orientation as the lac operon (84, 85) (Fig. 1).
Annual Reviews
www.annualreviews.org/aronline
Annu. Rev. Genet. 1972.6:133-156. Downloaded from arjournals.annualreviews.org
by University of Wisconsin - Madison on 03/29/07. For personal use only.
13 8
REZNIKOFF
therefore offers an opportunity to ask whether mRNA
terminators exist, or, in
other words, do RNApolymerase
molecules that transcribe lac i stop before
reaching the lac operon ? The answer is YES--evenwhere the transcription of lac
i has been dramatically elevated. However,transcription of the lac i gene does
read-through into lac when the lac #lac p deletion L1 is introduced, suggesting
that L1 removesthe lac i gene terminator (84, 86). Alternatively one could argue
that normal termination of lae i---qae read-through is a secondary (polar) result
of translation termination. Evidence against this possibility comes from the
observation that the polarity suppressor SuAdoes not effect lac i-)lac readthrough (87) and that, in order to fuse lae to another operon (i.e. trp), it seemsto
be necessary that the fusion deletion extend beyond the end of lac i (88).
Transcription of the trp operon does not cease with the translation termination signal of the last gene (trpA). Starting with the 480 dlac lysogen mentioned
previously in which the transposed lac operon is located in the same orientation
as the nearby trp operon
lac
(POED
CBA
.....
~onB .....
i
po~y~),
it has been possible to isolate trp-tonB--lac deletions that fuse the two operons.
In two cases, the deletions do not cut into trp,4, indicating that the trp mRNA
terminator is located some distance from the end of trp,4 (88). Recently Berger
Hardman(personal communication) have obtained results that suggest the same
conclusion. They have isolated the tryptophan synthetase A protein made in a
strain carrying a frameshift mutation near the end of the trl~,4 gene. The molecular
weight of this frameshift protein is approximately 25%larger than that for normal
. tryptophan synthetase A. The sequence of this added segment is currently being
determined. Wesuspect that the region in which our trp,4 + trp-lac fusion deletions end and the region attached to the trpA frameshift protein is not an unidentified gene. Rose &Yanofsky have shownthat there is not enough detectable
mRNA
produced beyond trp,4 to code for another "trp,4-size" protein (89).
In vitro evidence has suggested that a protein termed rho isolable from uninfected E. coli may play a role in the correct tcrmlnation of ~ mRNAs
(90).
direct in vivo evidence exists to suggest that rho (or a rho-like protein) acts
terminate mRNA
synthesis at the end of bacterial operons. However, Nissley et
al (91) have found that the addition ofrho to an in vitro transcription system using
~-gal DNAas a template blocks gal promoter stimulated transcription of k genes.
The site of rho action may be the gal terminator or a k terminator closely linked
to the gal substitution.
REGULATION OF (~ENE
EXPRESSION
Summary of model.--For each regulated operon there exists a specific
molecule termed the repressor, whose function is to control expression Of the
operon. The gene coding for the repressor is the regulator gene (an exampleis the
lac i gene in Figure 1). The repressor acts by binding to the operator region (o)
Annual Reviews
www.annualreviews.org/aronline
THE OPERONREVISITED
139
Annu. Rev. Genet. 1972.6:133-156. Downloaded from arjournals.annualreviews.org
by University of Wisconsin - Madison on 03/29/07. For personal use only.
thereby preventing transcription of the operonspecific polygenic mRNA.
For
each repressor-operatorsystemthere exists an effector molecule(the inducer or
the corepressor) that interacts with the repressor affecting its affmity for the
operator region. The corepressor (an end product or related compoundof
armbolicsystems)increases the repressor’s operator binding activity, while the
inducer acts in a reverse manner.This modelis pictorially representedfor lac
in Figure1. Somesystemsare not regulatedin this fashion.
Regulationoccursat the level of transcription.--Thecornerstoneof the operon
theory is that the decision for or against geneexpressionis madeat the levd of
transcription (3, 4). This hypothesis was primarily an economicalone, since
regulation at the translation level wouldresult in valuable energybeing squandered in makingunstable, unused mRNA.
A result that seemsto support this hypothesis is the observation that lac
specific mRNA
(pulse labeled RNAhybrizable to lae DNA)is found only in
inducedceils (24, 25). However,
since lac mRNA
is also missingfrominducedcells
that carry a translation defect in the lac z gene(a nonsensemutation)(25) we
not be sure that the absence of messengerunder repressed conditions is not an
indirect result of inhibited translation.
Anotherwayto discover the level at whichregulation occurs is to determine
the target site for the repressor. Geneticstudies havefailed to distinguishwhether
the operator is on the DNA
or on the mRNA.
Gilbert &Miiller-Hill (92) and
Riggset al (93) foundthat the lac repressor binds specifically and tightly to lac
operator double-stranded DNA.Other repressors (gal, A and 434) also bind to
DNAs
carrying their specific operators (94-96). Thesedata strongly support the
notion that operator DNA
is the in vivo repressor target site, although a DNArnRNA
hybrid or a hairpin mRNA
double-stranded region hasn’t been ruled out.
Finally, recent in vitro experimentsshowthat purified repressor can partially
block lac mRNA
synthesis in a purified DNA-RNA
polymerasesystem (55, 60).
Thissuggestsstronglythat lac regulationoccursat the level of transcription.
Althoughwe knowthat transcriptional control exists, several physiological
experimentssuggesttranslational control on top of transcriptional control (97).
Oneprobable mechanism
of translational control has been elucidated. T4 infection inhibits the translation of RNA
phagegenomes(98). In vitro protein synthesis studies have shownthat this defect in R17or MS2translation can be
localizedin the initiation factors (99) andmorespecificallyin the initiation factor
F3 (100). Whilethe experimentsdo not define howT4 infection alters F3 specificity for mRNA
ribosome binding sites, they identify a potential meansof
reversible regulation of translation. Onemerelyhas to postulate that F3-mRNA
recognition can be influenced by various effectors. It wouldbe an even more
attractive hypothesisif the cell had mutiple species of F3 protein each with a
preferred specificity for different mRNAs.
Multiple species of F3 havebeenfound
(101-105).
The repressor, negatiDecontroller of operonfunctioning.--The operonmodel
predicted the existence of two genetic regulator~ elements. Oneelement, the
Annual Reviews
www.annualreviews.org/aronline
Annu. Rev. Genet. 1972.6:133-156. Downloaded from arjournals.annualreviews.org
by University of Wisconsin - Madison on 03/29/07. For personal use only.
140
REZNIKOFF
regulator gene, whichcodes for the repressor, is independentfrom the operon
being regulated.
The importantexperimentalcharacteristics of the lac repressor and its gene
knownin 1961include the following: (a) The repressor is distinct from the enzymeswhosesynthesis it regulates. It has a distinctly different "substrate"
specificity than do these enzymes(106), and mutationsthat alter its functioning
(those resulting in unregulatedhigh level expressionof lac) reside at a site close
to but different fromthe structural genesof the lac operon(3,107, 108). (b) These
so-called lac i- constitutive mutationsresult froma loss of function since they
are recessive and since somelac i- mutationsare deletions (3, 109). (c) lac i
gene codes for a diffusible product since lac i + is dominantto (most) lac imutations even whenthe only functional lac structural genes are linked to the
lac i- allele (3, 109). This informationalso confirmsthe conclusiondrawnfrom
a and b above. (d) The lac i genedoes not code for an essential function since
strains carryinglac i- deletionsare viable (3). (e) Therepressoris the onlyregulator (negative or positive) specific for lac since all knownlac constitutive mutations reside either in lac i or in the repressor target site (o) (3) and all known
lac- mutationsare located either in lac z or lacy or in lac i (so-called lac i"
mutationsthat generatea repressor insensitive to the inducer)(3, 4, 110).
Theinitial belief that the lac repressor wasRNA
(3, 4) provedto be incorrect.
The subsequent isolation of temperature sensitive mutations and suppressible
nonsensemutationsin lac i suggestedthat the repressor was a protein (111,112)
as did studies with protein synthesis inhibitors (113, 114). The protein nature
the repressor wasconfirmedwhenit was isolated and purified (93, 115, 116).
The current biochemicalknowledgeabout the lac repressor indicates that it
undergoesthree different recognition reactions, all of whichare essential to its
functioning. Individual lac repressor protein moleculescan recognizeeach other
and aggregate into tetramers (117, 118). Both the monomeric
and the tetrameric
formsof the repessor bind isopropyl-fl-thiogalactoside (IPTG),an inducer of the
lac operon(115, 117). Therepressor tetramerbinds specifically and tightly to the
lac operatorregion (92, 93).
Theability of the repressor to bind to the operator (and thereby inhibit lac
transcription) is determinedby the other two reactions described above. Only
the tetrameric formof the repressor can bind to the operator and there exists
only one binding site/tetramer (119). This suggests that repressors that fail
aggregatewill not repress. Presumably
somei- mutationsmight lead to defective
aggregation properties. Furthermore, mixed tetramers of operator binding
active and inactive monomers
should be inactive in operator binding. This is
thought to be the explanation for the rare transdominantlac i-(t "-~) mutations
(120, 121). IPTGcauses a drastic reduction in the repressor’s affinity for the
operator (92, 93) presumablyby altering the repressor’s conformation. Mutations that decrease the repressor’s affinity for the inducer shoiald havean uninduciblephenotypeand, in fact, the lac i ~ mutationsthat manifestthis phenotype
do have a loweraffinity for IPTG(119).
Annual Reviews
www.annualreviews.org/aronline
Annu. Rev. Genet. 1972.6:133-156. Downloaded from arjournals.annualreviews.org
by University of Wisconsin - Madison on 03/29/07. For personal use only.
THE OPERON REVISITED
141
Recently experiments by Eron & Block (55) and deCrombrugghe et al (79)
have shownthat the repressor can partially block in vitro synthesis of lac mRNA
in a purified RNApolymerase-/ac DNAsystem and that this activity is reversed
by IPTG.
To a first approximation, these results seem to apply to other systems. In
several cases apparent regulator gene mutants have been isolated. These usually
carry recessive mutations that result in constitutive expression of the system.
These regulator genes also supply a nonessential function to the cell. In many
cases one suspects that the active repressor is a protein since suppressible nonsense mutations have been isolated and in at least four cases the repressor is
thought to be an oligomer of identical subunits, since either intracistronic complementation or negative complementation has been detected [lon (122), metJ
(123), the R-factor repressor (124), trpR(125)]. However, in on ly a ver y few
cases has the repressor been isolated and shownto act in vitro as predicted [the
phage k repressor (95), the phage 434 repessor (96), and gal repr essor (94)
In some other systems, the evidence is not yet consistent with the classic
regulator gene concept. For instance, many man years of work have been invested in studying the regulation of the his operon and yet the regulator gene is
still to be identified.
Thefirst striking fact about the his systemis that there is a multiplicity of loci
where mutations can produce constitutive his phenotypes. These include five
locations (hisS, hisR, hisT, hisU, and hisW) in addition to the his operator region
(126, 127). Mutations in three of these loci are knownto be recessive and in all
cases but one (hisR) the products of these genes are thought to be diffusible
proteins (11, 17). These data would suggest an embarrassmentof riches (too many
repressors) but in fact, closer examination suggests that all the loci can be
relegated to roles in his regulation other than the production of the his repressor
protein. The real physiological effector (the corepressor in this case) is often
"processed product" of the "effector" added to the cell suspension and in this
case His-tRNAis thought to be the effector (11, 17, 128). All five of these his
regulatory loci appear to play roles in the production of or modification of HistRNA; hiss codes for His-tRNA synthetase (129), hisR ais
may code for tRNA
itself (130) or a positive regulator of its production, hisT codes for a tRNA
modifying function that introduces two pseudouracils into the anticodon loop
ms (131), and hisU and hisF¢ appear to play a role in the maturation of
of tRNA
rri~ and other tRNAs(11, 17, 128).
tRNA
One can therefore conclude that the mere presence of constitutive mutations
in a particular locus, even recessive, nonsense, constitutive mutations that do not
impair cell growth, is insufficient evidence to identify the locus as the regulator
gene. (The reader should be warned that I shall make this mistake subsequently.)
The product of the regulator gene must be demonstrated to have an affinity for
the real physiological effector and an affinity for the operator. Furthermore, it
must be shownto "do its thing" in vitro.
Before presenting the current favored hypotheses about the identity of the his
Annual Reviews
www.annualreviews.org/aronline
Annu. Rev. Genet. 1972.6:133-156. Downloaded from arjournals.annualreviews.org
by University of Wisconsin - Madison on 03/29/07. For personal use only.
142
REZNIKOFF
regulator gene, it mightbe wise to definethe conceptof the regulatorgeneso that
the reader can decide whythe regulator genemaynot have beenfoundin this and
certain other cases.
(a) The regulator gene exists. A priori, one wouldpredict that the cell must
provide an operon-specificmacromolecule
(the regulator) throughwhichan effector can exert a regulatory influence on the mRNA
or protein synthesizing machinery. To require operonspecific effectors to interact directly with the RNA
polymerasecomplex(or protein synthesizing complex)thereby inttuencing its
recognitionof a series of operons,is to demand
incredibleflexibility and specificity of a generalizedsynthetic system.Theregulator genecodesfor this regulator.
(b) Theregulator is a repressor--i.e., it acts in a negative fashion. Work
the ara system(9, 132), the maltose(real) system(133), and the rhamnose(rha)
system(134) has shownthat this generalization is not valid. Thesesystemsare at
least partially controlled by specific regulatory proteins (so-called activators)
whichact in a positive fashion. Constitutive mutations in an activator should
be rare.
(c) Theregulator acts throughthe cytoplasm.A cis-active protein is apparently codedfor by the phageP2 Agene. This protein is necessaryfor P2 geneexpression or DNA
synthesis, or both (135). Thepresenceof a cis-active regulatory
protein could be detected by the presence of cis-domiaant nonsense mutations
effecting derepressionof the operon(or, for positive control, pleiotropic negative
shutdownof the operon).
(d) The regulator has no other function. In certain cases the regulator may
havesomeother function necessaryfor cell survival. This wouldinclude the possibility of the regulator being a necessaryenzymeor being a regulator of another
essential system.
(e) Theregulator geneis an independentunit of transcription. Thegene coding for the regulator mightbe an operator proximalgene in an operonwhosedistal genes perform necessary functions. Anypolar mutation wouldprevent expression of these necessary genes. Oneexampleof an apparent regulator gene
being part of a larger operonis foundin the histidine utilization (hut) system
(136).
(f) There is one unique regulator gene for each system. There maybe more
than one regulator gene for a given system. Only very rare double mutations
wouldgive rise to a constitutive phenotype.Recentexperimentshave indicated
that the ~,-N geneoperator, as definedby the ~2 mutation,is the target site for
both the cI and tofgene products (137).
Toreturn to his, current hypothesessuggest that either the hiss gene(which
codesfor the His-tRNA
synthetase) or hisG (whichcodes for the first enzymein
the his pathway,phosphoribosyl-ATP
synthetase), or both is the his regulator
gene(11, 17). Theevidencefor the role of hisS comesnot fromthe constitutive
hiss mutations (these can all be explained by their effect on the His-tRNA
pool
size) but on someobservations by Wyche,whoisolated hiss mutationsthat prev~n~;full .derepressionof the ttis operon,H.e ~tl~o fo~n~dth.at in h~terologou~
Annu. Rev. Genet. 1972.6:133-156. Downloaded from arjournals.annualreviews.org
by University of Wisconsin - Madison on 03/29/07. For personal use only.
Annual Reviews
www.annualreviews.org/aronline
THE OPERONREVISITED
143
S. abony/S,typhimurium
hisS+ diploids the basal level of his expressionis raised
(138). Theseresults are compatiblewith the hisS protein havinga direct positive
role in his regulation or mightbe explainedby altered affinities of the His-tRNA
m~. This latter possibility is being examined(B. N. Ames,
synthetases for tRNA
personal communication).His-tRNAsynthetase binds to DNA.Whether it is
specific for his operator DNA
is also being studied (Ames).
The possibility of a regulatory role for the hisGenzymecomesfromthe followingobservations.The enzyme’sfeedbackresistant propertyaffects the regulation of his operonexpression(139, 140), the enzyme
displays an affinity for HistRNA(141), and the enzymebinds to DNA
in general and his-DNApreferentially
(R. Goldberger, personal communication).Evidencethat suggests that hisG is
not a regulatory gene(or at least not the uniquehis regulator gene) includes the
facts that no constitutive mutationshavebeenisolated in hisG, and in the cases
tested so far, hisG nonsense and frameshift mutations have no effect on his
regulation (17).
The operator, the binding site for the repressor.--Jacob,Monod
and coworkers
postulated that there must be a target site for the repressor. Theytermedthis
target site the operator. In 1960mutations were isolated whosephenotypeand
location fit the predicted properties of operator mutations(35). Thesemutations
had a partially constitutive phenotype, they mappedbetweenthe promoter and
the start of the lac z gene, and their constitutive phenotypewascis-dominant(3,
4, 35, 54). Subsequentin vitro experimentsdemonstratedthat DNA
containing
lac o~ mutations has a lower affinity than wild type DNA
for the lac repressor
protein (92, 93) and, therefore, that the operatorregion, as definedby these mutations, is the bindingsite for the repressor. Similarcis-dominant
operatorconstitutive mutationshave been isolated for manyother systems.
The most detailed genetic study of an operator region was accomplishedby
Smith &Sadler, whoisolated and characterized hundredsof independentpoint
mutationsin the lac operator region (66, 142). Their findings indicate at least
and probably16 sites in the operatorwherea transition or a transversionmutation
can generate a detectable oc phentoype.Operatormutationsfoundat a given site
all seemto generate the samephenotype.Althoughthe total phenotype(i.e. the
maximallevel of expressionand the inductionratio) is uniqueto the c mutations
of a given site, there appearto be pairs of sites in whichthe o° mutationshave
identical induction ratios. Moreoverthe tentative maporder suggests that these
pairedsites are arrangedin a bilaterally symmetrical
fashion.It is surprisingthat
the operator’s base sequence, as deducedfrom mutagenicdata, does not reflect
this phenotypic symmetry(somematchedsites do not have the same nucleotide
pairs).
Anotherimportantgenetic systemfor studying the lac o region results from
the isolation of tonB- deletions endingin or near lac o. Wehavedevelopedtechniquesthat allowus specificallyto isolate deletionsendingin this region(86, 88).
Theparental strain contains: a ~b80dlac prophagesuch that the trp and lac ope-
Annual Reviews
www.annualreviews.org/aronline
Annu. Rev. Genet. 1972.6:133-156. Downloaded from arjournals.annualreviews.org
by University of Wisconsin - Madison on 03/29/07. For personal use only.
144
REZNIKOFF
rons are in the sameorientation, a trpR- mutationresulting in constitutive exr
pression of the trp operon, and a lac promoterpoint mutationreducing lac expression. Fromthis strain we isolated tonB- mutations that simultaneously
becamelac+. All the deletions analyzedto date end betweenlac i and lac z. We
hopethat these deletions will facilitate fine structure deletion mappingof lac o.
Since someof these ddetionsremoveall or mostof lac o but still recombinewith
all lac z mutationsand haveretained the ability to synthesizewild type/~-galactosidase, webelieve that lac o andlac z are distinct geneticentities (58, 143). This
result confirmsthe previousfinding of Steers et al that a lac oc mutationdoesnot
affect the properties of/~-galactosidase(144).
In a positively controlled systemthere shouldbe an operonlinked target site
for the activator protein. Starting with a strain carrying a deletion of the ara
positive regulatory element(the araCgene), ethyl methanesulfonate and 2-aminopurine inducedara+mutationsin the putative target site for the araCactivator
havebeenisolated (145, 146). Thesemutationsare called ~ (initiator c onstitutive) mutations. Theyare located near the arabgene, at one end of the operon,
and result in expressionof the ara operonin the absenceof the activator. Similar
mutations have been isolated by Hofnung&Schwartzin the real system(147).
However,the araIc mutationsdo not by themselvesgenerate ara transcription
+ allele (145, 146, 148). This is thoughtto be bewhenin the presenceof an araC
cause the araCprotein has both a repressor function (in the absenceof the inducer, arabinose) and an activator function (in the presenceof the inducer).
araIo mutationsobviate the necessity for the secondfunction but still leave the
operonsensitive to the first, suggestingthat the C-repressorhas an additional
target site, the ara operator. Deletion analysis indicates that the ara operator
resides betweenaral and araC(araO-araCdeletions makethe aral~ stimulated
+ allele). Sucha double
transcription of ara insensitive to the presenceof an araC
control systemseemsquite complicated.It required the evolution of two genetic
signals (araI and araO) which are recognized by the same protein in two apparently different ways.The ara systemis currently beinganalyzedin vitro (149151) to test this repressor-activatormodel.
Theeffector. A compound
whichcontrolsthe affinity of the regulatorproteinfor
the operator(or initiator).--Essential to a description of generegulation in bacteria is the ducidationof howthe metabolitesthat either induceor repress enzyme
synthesis can effect their roles. The Jacob-Monod
operon modelpredicts that
these metabolites influence gene expression by interacting with the particular
regulatoryprotein and altering its affinity for the particular geneticsignal associated with the operon.In general the in vitro workwith the lac repressor has substantiated this model.
However,the effector is frequently not identical to the metabolite addedto
the cell suspension.Forinstance, in the case of the lac operon,lactoseitself is not
the inducer (152). Rather the physiologicalinducer is allolactose, an isomer
lactose generated from lactose by/~-galactosidase (153). Traditionally, people
Annual Reviews
www.annualreviews.org/aronline
Annu. Rev. Genet. 1972.6:133-156. Downloaded from arjournals.annualreviews.org
by University of Wisconsin - Madison on 03/29/07. For personal use only.
THE OPERON REVISITED
145
studying the regulation of the lac operon have sidestepped this problem by using
gratuitous inducers, which are chemically synthesized, nonmetabolizable compounds such as isopropyl-~-D-thiogalactoside (IPTG).
Groups studying the regulation of biosynthetic operons have been very interested in tracking downthe true physiological effectors, Eidlic &Neidhardt (154)
isolated a temperature sensitive mutant in E. coli that helped define one such
effector. This mutant carried a defect in the Val-tRNAsynthetase which, at nonpermissive temperatures, not only stopped cell growth but also derepressed the
synthesis of the isoleucine-valine (ilv) regulon enzymes.Subsequent studies, with
va~ under
valine analogs and with measurements of the level of charged tRNA
various conditions, indicated that a likely candidate for an ilv regulon effector is
Val-tRNA(155, 156). As indicated in mydiscussion of the his system, other (but
not all) amino acid system effectors are also charged tRNAs.
A particularly complexsituation seems to exist in biosynthetic systemswhich
lead to the synthesis of more than one end product. In several cases repression
appears to result from the presence of more than one effector. Experimentally
two different kinds of multi-effector systems have been discovered: cumulative
repression, in which total repression seems to result from an additive repression
of each of the individual effectors [as in the regulation of carbamyl phosphate
synthetase synthesis by arglnine and a pyrimidine (157)], and multivalent repression, in which all the effectors must be present for repression to occur (158).
Despite the apparent differences between these two systems, we don’t know
whether they are mechanistically different (H. E. Umbarger,personal communication) so we will tentatively consider them as a group using the more common
term multivalent repression.
Perhaps the best analyzed systems involving multivalent repression are the
threonine, isoleucine, valine, .and leucine metabolic pathways. Threonine is a
precursor for isoleucine biosynthesis; therefore, it is not surprising that the synthesis of the threonine biosynthetic enzymesis repressed by both isoleucine and
threonine (159). Most of the enzymesused in the biosynthesis of isoleucine from
threonine are also used for valine biosynthesis and for the synthesis of a-ketoisovalerate a precursor for leucine biosynthesis. Synthesis of these enzymes(or at
least those coded for by ilvA, ilvD, and ilvE) is subject to multivalent repression
by isoleucine, leucine, and valine. The steps unique to leucine biosynthesis are
subject only to leucine repression (158). In all these cases charged transfer RNAs
maybe the actual effectors (155, 160-162).
Several questions emerge from these studies. Does multivalent repression
result from all the effectors interacting with one repressor protein or are there
several different repressor proteins, each specific for a given effector ? In the latter
case all the repressors would have to act in concert in order to generate complete
repression. If there is a single repressor for, say, ilvA, ilvD, and ilvEwith affinities
for multiple effectors, does the same repressor protein also regulate other
operons that are sensitive to only one or two of the effectors ?
JEvid,enc¢ from another mu!tivalent repression system suggests that one r~pres-
Annual Reviews
www.annualreviews.org/aronline
Annu. Rev. Genet. 1972.6:133-156. Downloaded from arjournals.annualreviews.org
by University of Wisconsin - Madison on 03/29/07. For personal use only.
146
REZNIKOFF
sor can be influencedby multiple effectors. Thefirst step of the common
aromatic
aminoacid biosynthetic pathway(PEPq-E4P---~DAHP)
is catalyzed by three isoenzymes.The synthesis of one of these isoenzymes(aroG-DAHP
synthetase) is
repressed by phenylalanineand tryptophan(163, 164). The synthesis of a second
DAHP
synthetase (that codedfor by aroF)is repressed by tyrosine but high concentrations of phenylalanine(and possibly tryptophan)can substitute for tyrosine
(163, 165). This rather complexpicture has beenclarified considerablyby the fact
that mutations in one locus, tyrR, can result in derepression of aroG-DAHP
synthetase, and aroF-DAHP
synthetase (165, 166). Other mutations in tyrR derepress aroF-DAHP
synthetase but not aroG-DAHP
synthetase. If tyrR codes for
a repressor protein, these results suggest that this repressor has affinities for
phenylalanine,tyrosine, and tryptophan(or their derivatives), that these effeCtors
differentially affect the repressor’saffinity for different operatorregions,andthat
it is possible to alter by mutation one effector-repressor interaction without
drastically affecting the others.
There is one system thought to be controlled by more than one regulatory
protein althoughit is not clear whethermultiple effectors are involved. Thegal
operonis regulated by the gal repressor gene galR (167) and by Ion, the gone
regulating capsular polysaccharidesynthesis (122, 168). This regulation seems
be independent
andadditive(168). It is similar to that seenfor lac in trp-lae fusion
strains in whichboth trp and lac operators are present and whereboth repressors
can control lac expressionin an additive fashion (86).
A generalassumption
in the study of E. coil generegulationis that if an effector (or effectors) influencesthe productionof enzymescodedby genesin different
operons,then one repressor mustbe responsible for directly influencing the productionof all of them. Thisignores the possible existenceof "cascaderegulation"
whereone of the operonscodesfor the productionof an internal inducer (and an
activator ?) whichturns on one or moreof the other operons.It turns out that the
ilvC gene is probably controlled by an induction mechanism
in whichtheproduct
of the ilvB enzyme’sactivity is the inducer(169).
Trm MOLECtrLAIi BASIS OF RECULATION
lntroduction.--The Jacob-Monodoperonmodelis a broad conceptual frameworkcontaining few molecular details. Nowthat we can define the framework
with someaccuracy(at least in the case of lac) it is probablyworthconsidering
the molecular basis for that framework.For instance: Howdoes transcription
initiation occur ? Howdoes the repressor act to block transcription ? Whatis the
structure (size and sequence)of the operator ? Howdo proteins suchas the repressor and RNApolymerase recognize specific DNAsequences ? These questions
merelyscratch the molecularsurface of the problem.
Howdoes transcription initiation occur?--Recentin vitro work,such as that
by Zillig et al (170) has suggestedthat transcription initiation is composed
of
series of reactions. The first of these is the binding of RNA
polymeraseto DNA.
Annual Reviews
www.annualreviews.org/aronline
Annu. Rev. Genet. 1972.6:133-156. Downloaded from arjournals.annualreviews.org
by University of Wisconsin - Madison on 03/29/07. For personal use only.
THE OPERONREVISITED
147
Onecan ask whetherRNA
polymerasebinding is the only sequencespecific step
(the other steps followingautomatically) or whethereach step in transcription
initiation requires a specific DNA
signal. In other wordsdoes the promotercontain one or manysignals ?
Wenowhave evidence that the X-Ngene promoter is probably composedof
at least twospatially separate signals--a signal definedby the N promotermutations t27 and sex (possibly the RNA
polymerasebinding site) and the "start"
site whereN-mRNA
synthesis actually originates. Blattner et al (64, 65, Dahlberg
&Blattner, personal communication)have accomplisheda hybridization and
sequenceanalysis of the first 160 nucleotides of in vitro synthesized N mRNA.
This N mRNA
sequence, whichis identical to the in vivo sequence(171), starts
outside of the #nm434region found in X-imm434hybrids. Fromgenetic studies
and electron micrographicheteroduplex mappingdata we knowthat the t27 and
sex mutations are probably located 200 or more nucleotides within the imm434
region. This informationindicates that for N-genetranscription to occur, RNA
polymerasebinds at one site, and then migrates a distance greater than its own
diameter to a secondsite whereN-mRNA
synthesis starts.
Indirect evidencesuggeststhat the lac promotermutationsalso alter a genetic
signal necessaryfor an early step in transcription initiation. Perhapsthe signal
altered is the RNA
polymerasebinding site. In vitro studies by de Crombrugghe
et al (79) havesuggestedthat cyclic AMP
is required for the formationof a lac
specific, rifampicin resistant, RNA
polymerase-DNA
complex.The lac promoter
mutations reduce the cyclic AMP
requirementfor lac operonexpression (69-71).
Thusthese mutationsprobablyalter the signal necessaryfor the formationof the
RNApolymerase-DNA complex.
Howdoes the repressor block transcription ?--Doesthe repressor block binding of RNA
polymerase,initiation of RNA
polymeraseactivity, or elongation of
the messenger? We(86) tried to generatean in vivo systemfor analyzingthe mode
of lac repressoraction. Trp-lacfusiondeletions wereisolatedin whichlac expression
was the result of RNA
transcription initiating at the trp promoterone or more
genes removedfrom the lac operator region. Wethen asked whetherlac repressor
could block read-throughtranscription. If it did, one could concludethat the
repressor was capable of(and by analogy, acted by) blockingprogressof prebound
RNA
polymerase. Unfortunately the answer was ambiguous(partial repression
occurred).
Chert et al (60) examinedrepressor action in vitro by adding lac repressor
either before or after adding RNApolymerase. Repressor added prior to RNA
polymeraseaddition blocks lac mRNA
synthesis 2- 4-fold. Addition of IPTGto
this mixturein the presenceof rifampicin results in inductionof lac mRNA
synthesis suggesting that RNApolymerasebinding is not prevented by the repressor. The opposite order of addition was also tried. Repressor can block mRNA
synthesis by preboundRNA
polymeraseand this repression is reversible by the
addition of IPTG.
Annual Reviews
www.annualreviews.org/aronline
Annu. Rev. Genet. 1972.6:133-156. Downloaded from arjournals.annualreviews.org
by University of Wisconsin - Madison on 03/29/07. For personal use only.
14 8
REZNIKOFF
Theseresults (and others relying on the in vitro lac system)are promisingbut
they shouldonly be taken as tentative becauseof the nature of the in vitro system.
Theontogenyof the in vitro lac systemreflects a long search for conditions that
accuratelyreflect in vivophenomena
(i.e., sensitivity to repressor-inducercontrol,
to CAP-cyclicAMP
control, to promoter mutations). Thoughone suspects that
the initial experimentaldifficulties were due to real problems,such as phage
specific promoteractivities, it wouldhavebeen morepleasing if the systemhad
workedcorrectly in a straightforward manner.Furthermore,the response of the
in vitro systemis onlyqualitatively equivalentto the in vivoresults. For instance
the real repressionratio is 2,000fold, not 2- 4-fold. This discrepancyis probably
due to real experimentaldifficulties but nonethelessshould makeus realize that
the results mightnot reflect the true in vivoevents.
Similar in vitro experimentshave been done using a ~-DNA
system (171a).
These experiments show that N mRNA
synthesis by prebound RNApolymerase
can be blockedby the ~,cI repressor. However,the reverse reaction sequenceindicated that ~,cI repressor not only blocks mRNA
synthesis, but also blocks RNA
polymerasebinding.
The previouslymentionedstudies on X-Ntranscription (64, 65, 171) also tell
us something about the modeof action of the kcI and tof repressors. The N
operator, as defined by the o2 mutation, resides inside the imm434substitution
close to the sites occupiedby the sex and t27 mutations.Since this is a considerable distance from wherethe N mRiNA
starts, the cI and tofgene products must
blockan early step in transcription initiation. In the lac and trp systems,however,
genetic evidencesuggests that the operators are transcribed [lac o-lac z and trp
O-trp A deletions do not impair operon expression (59, 62).] Thus the lac and
trp repressors maypreventa late step in transcription initiation.
Structure of the lac operator.--Onecan argue a priori that the lac operator
region must be equal to or larger than the minimum
nonrepeatingsequencelength
for the E. eoli chromosome
(or >12 nucleotide pairs). Qualifying statements
wouldinclude the fact that single nucleotide pair modificationsof the operator
mustalso not be foundelsewhere(142) (the necessarylength increases) and
the existenceof operator-like repeats wouldnot be just the result of randomprobability but wouldbe subject to negativeselective pressure (the necessarylength
shrinks). The minimum
operator length has been determinedexperimentally by
Smith&Sadler, whodiscoveredthat there are at least 12 sites capableof generating o° mutations(66, 142). Recently Sadler and coworkers(personal communication) and Gilbert and coworkers(personal communication)
have isolated operator DNA
using the fact that repressor boundto the operator will protect it from
digestion by DNase.Sadler’s group estimates, from the fraction of input q~80
dlac DNAthat is protected by repressor from DNaseattack, that the operator
is < 75 pairs long.
Mutagenicdata indicate that both A-Tand G-Cpairs exist in the operator
with A-T pairs predominating(66). This fits with someexperiments by Riggs
Annual Reviews
www.annualreviews.org/aronline
THE OPERONREVISITED
149
Annu. Rev. Genet. 1972.6:133-156. Downloaded from arjournals.annualreviews.org
by University of Wisconsin - Madison on 03/29/07. For personal use only.
et al (172) whichshowthat the repressor interacts best with A-Trich DNA
polymers, althoughthis interaction is still several orders of magnitude
less than that
manifestedby the repressor towardsthe real operator.
Structure and function of the repressor.--A strong effort is being madeto
elucidate the structure of the lac repressor using both genetic and chemicaltechniques. Thanksto a large collection of tonB--laci- deletions isolated by Miller,
it has beenpossible to mapmanylac i mutations(16, 173,174,174a).
The lac i -d mutationsare clustered close to the aminoterminusof the lac i
gene. Thesemutationsare knownto result in the formationof repressor molecules
that cannot interact with the operator (173, 174, Jackson, personal communication). Theeffect of these lac i-d mutationsmaybe due to a direct changein the
operator bindingsite, or they mayalter the repressor’s conformationalproperties
so that operator binding cannot occur, or they maydestroy the operator binding
capability by changingthe repressor’s aggregationproperties. The observation
that these mutationsare transdominantsuggests that their subunits mayaggregate moreslowly than normal repressor monomers
(16). Sadler (personal communication)has recently isolated pseudorevertantsof lac o~ mutationsthat map
in lac i. Thesemutationsmayhelp to define the operatorbindingsite and it would
be interesting to see if they werelocatedclose to the lac i TM mutations.
Mutationsknownto decreasethe affinity of the repressor for IPTG(the slac i
mutations)(119) mapin five different locations (16, 173). Againwe do not
whetherthe effect is a direct or indirect result of the mutationalchange.In one
case (that located closest to the lac i-a region)the lac i s mutationalso alters the
affinity for the operator(16, 119, 173, 174). Thelac i t mutation[whichgeneratesa
repressor showinga higher affinity for the inducer (115)] maybe located in the
IPTGbinding site.
To achievethe long range goal of determiningthe structures of wild type and
mutantforms of the lac repressor and perhapsof various repressor-inducer and
repressor-operator complexes,sequence, crystallographic, and other physiochemicalstudies are being initiated. Asequenceanalysis of the first 50 amino
acids of the wild type repressor has beenaccomplishedand sequencesof repressors carrying various i -d mutations are being examined(174, 174a, Fanning,
personal communication).Manyof these mutations should alter this sequence
(174). Crystals of the repressor have been obtained and are currently being
analyzed(Steitz, personal communication).
Mechanism
of protein-DNArecognition reactions.--See the article by Peter
yonHippelfor a conciseanalysis of this topic (175). Manyof the ideas presented
beloware similar to his.
If one assumesthat DNA
is uniformly in the B structure, then one can immediately concludethat there exists no sequencespecificity in the phosphate
groups or the deoxyribose groups and that the normal hydrogenbondingsites
of the purine-pyrimidinepairs wouldalso be unavailable to an outside reactant.
Annual Reviews
www.annualreviews.org/aronline
150
REZNIKOFF
Annu. Rev. Genet. 1972.6:133-156. Downloaded from arjournals.annualreviews.org
by University of Wisconsin - Madison on 03/29/07. For personal use only.
BROAD GROOVE
I
c--c
.....
c--c
\
\_,
/
...,/...,
ill..."
NARROW GROOVE
Flo. 2. Possible reactive groupsin the broadand narrowgroovesof DNA.
Groups
absolutelyuniqueto a singlebaseare circled. Thosethat perhapsare onlyuniqueto the
purinesor pyrimidinesare in squares.
Is there anything else ? Observationof a space-filled modelof DNA
suggests a
wealth of specific informationavailable for protein-nucleic acid interactions in
the broad and narrow grooves. Most of these are possible hydrogenbonding
sites but somehydrophobicgroupsare present too. Manyof these sites havebeen
describedby Wilkins(176). In Figure 2 1 havetried to indicate somepotentially
important groups. Those circled maybe absolutely unique centers (defining
single base) while those enclosedin squares mayeither provide absolute uniqueness or twofold uniqueness (distinguishing purines from pyrimidines). Kelly
Smithfound evidencefor both types of specificity in the Haemophilisinfluenza
restriction enzyme
recognitionsite (177). Thedata for lac o give evidenceonly for
completelyunique base pair recognitions (66, 142). Recently, Adleret al (174)
Annual Reviews
www.annualreviews.org/aronline
Annu. Rev. Genet. 1972.6:133-156. Downloaded from arjournals.annualreviews.org
by University of Wisconsin - Madison on 03/29/07. For personal use only.
THE OPERONREVISITED
151
haveusedthe sequenceof the first 50 aminoacids in the lac repressor to construct
a molecularmodelof the repressor-operatorinteraction. In this model,a portion
of the repressor’s N-terminuslies in the broad groove whereit specifically
recognizessomeof the uniquecenters pictured in Figure 2.
Our assumption that DNAresides in a uniform B structure is probably not
valid. DNAseemsto "breathe," that is, it undergoeslocalized denaturations
(178), suggestingthat the normalinternal hydrogenbondingsites could be opened
up for interactions. Since hydrogenbondswouldbe lost in the process, this
configurationprobablywouldnot be favoredunless stabilized by a specific interaction with a protein. Suchan event might be considereda protein inducedconformational change in the DNA.
Furthermore, as postulated by Gierer (179), somestretches of DNAmay
composedof two identical antiparallel sequences capable of forming hairpin
loops similar to those found in tRNA.Suchloops might give uniqueness to the
phosphate and deoxyribose groups. The possible twofold symmetryof the
operator (142) suggestssuch a structure. Sincehairpin loops wouldbe less stable
than the normal conformation because somehydrogen bondingsites wouldbe
lost, they probablywouldhave to be inducedby protein too.
Finally, the B structure of DNA
is probably an average structure. A short
stretch of DNAmayhave a slightly different conformationgenerated by the
base compositionand sequenceof the particular region. This possibility has been
deducedlargely from physicochemicalstudies of synthetic DNA
polymers composed of defined repeating sequences[e.g., see Wells et al (180)]. Sequence
specified changesin the helical structure couldgeneratestructure specific (i.e.,
sequencespecific) deoxyriboseand PO~groupsfor recognition purposes.
It is also possible that aromaticaminoacid residues of a protein mightintercalate into a DNA
molecule (181). A possible modelfor such an intercalation
comesfrom the crystallographic analysis of puromycin,whichshowsan intermolecularstacking of adenine bases and the tyrosine aromaticrings (182). However, it is not clear wherethe specificity for such a reaction wouldcomefrom.
Theseare rather simple ideas about a very complexsubject. Dowe have any
evidence for any of these recognition mechanisms? Right nowwe are at the
intuitive guess stage. I personally believe that several mechanisms
are active.
For instance, at somespecific stage in its interaction with DNA,RNA
polymerasemustinduce (or stabilize) localized denaturation. Therepressor probably
doesn’t denature DNA
(the protected piece is doublestranded) (Sadler, personal
communication).Furthermore, more than one type of reaction maybe involved
in a particular instance. For instance, the lac repressor interacts with
poly(dA--dT), poly(dA-dT)equally well in the presence or absence of IPTG.
The binding constant for this interaction is approximatelytwo orders of magnitude lower than that of the repressor-IPTGcomplexfor the operator (Riggs,
personal communication).Perhaps the repressor mayinteract with any DNA
that has an overall structure similar to poly(dA--dT),poly(dA--dT)but
the correct sequence,as determinedby the reactive groupsin one or both grooves,
Annual Reviews
www.annualreviews.org/aronline
15 2
REZNIKOFF
can result in verytight repressor-operatorinteraction. Theinducermayinhibit a
conformationalchangein the repressor necessaryfor this very tight binding.
Myprejudice is that only base specific interactions in the broadand narrow
groovecan give the necessary specificity for the tight binding. There are too
manysequencespecific proteins (repressors, restricting enzymes,modifyingenzymes,etc.) for eonformafionalvariations to do it alone.
Annu. Rev. Genet. 1972.6:133-156. Downloaded from arjournals.annualreviews.org
by University of Wisconsin - Madison on 03/29/07. For personal use only.
ACra~OW~.~DCES~,~TS
The author’s research is supported by a National Science FoundationGrant
(GB-20462),a Wisconsin Alumni Research Foundation Grant, and a National
Institutes of Health BiomedicalSciences Grant. I amparticularly indebted to
J. Beckwith,T. Davis, P. E. Hartmanand C. A. Thomas,Jr. for stimulating my
interest in gene regulation. I am also very thankful to M. Sundaralingam,
R. Wells, and F. Blattner for helping medevelop myideas about protein-nucleic
interactions. Others whosecommentsor preprints were very helpful include:
B. N. Ames,H. Berger, S. Bourgeois, K. D. Brown,J. Cairo, G. Craven, J. Dahlberg, J. E. Davies, W.Dove, G. H. Echols, E. Fanning, T. Fanning, R. Goldberger, J. Hardman,
R. Hazelkorn,D. Jackson,A. Jobe, B. Mtiller-Hill, I. Pastan,
A. Riggs, J. Sadler, R. Sommerville, T. Steitz, W. Summers,W. Szybalski,
R. Wixom,H. E. Umbarger,3. Miller, and T. Platt.
Annual Reviews
www.annualreviews.org/aronline
THE OPERON REVISITED
Annu. Rev. Genet. 1972.6:133-156. Downloaded from arjournals.annualreviews.org
by University of Wisconsin - Madison on 03/29/07. For personal use only.
LITEIIATUEE
153
CITED
1. Monod,J., Cohn, M. 1952. Advan.
S2~ring
HarborSyrup. Quant.Biol.
Enzymol. 13:67-119
:363-72
2. Vogel, H. J. 1957. The Chemical
26. Nathans, D., Notani, G., Schwartz,
Basis of Heredity, ed. W. D. McJ. H., Zinder, N. D. 1962. Proc.
Elroy, B. Glass, ~76-89.Baltimore,
Nat. Acad. Sci. USA48:1424-31
Md.: Johns Hopkins. 848 pp.
27. Summers,W. C. 1970. J. Mol. BioL
3. Jacob, F., Monod,J. 1961. J. Mol.
51:671-78
Biol. 3:318-56
28. Marrs, B., Yanofsky,C. 1971. Nature
4. Jacob, F., Monod, J. 1961. Cold
NewBiol. 234:168-70
Spring HarborSyrup. Quant. Biol.
29. Demerec, M. 1956. Cold Spring
26:193-209
Harbor Symp. Quant. Biol. 21:
5. Beckwith, J. R. 1970. The Lactose
113-20
Operon, ed. J. R. B eckwith, D.
30. Ames,B. N., Garry, B. 1960. Proc.
Zipser, 5-26. Cold Spring Harbor,
Nat. Acad. Sci. USA.45:1453-61
N.Y.: Cold Spring Harbor Labora31. Kalckar, H. M., Kurahashi, K.,
tory. 437 pp.
Jordan, E. 1959. Proc. Nat. Acad.
Sci. USA45:1776-86
6. Gilbert, W., Mfiller-Hill, B. SeeRef.
32. Ames,B. N., Garry, B., Herzenberg,
5, 93-110
L. A. 1960. J. Gen. Microbiol. 22:
7. Miller, J. H. See Ref. 5, 173-88
8. Magasanik,B. See Ref. 5, 189-220
369-78
9. Englesberg,E. 1971. MetabolicRegu33. Franklin, N. C., Luria, S. E. 1961.
Virology 15:299-311
lation, Metabolic Pathways, ed.
H. J. Vogel, 5:257-96. NewYork:
34. Englesberg,E. ~961.J. Baeteriol. 81:
Academic.3rd ed. 5 vols. 576 pp.
996-1006
10. Zubay, G., Chambers,D. A. See Ref.
35. Jacob, F., Perrin, D., Sanchez, C.,
9, 297-349
Monod,J. 1960. C. R. Acad. Sci.
11. Brenner, M., Ames,B. N. See Ref. 9,
1727-29
350-88
36. Scaife, J., Beckwith,J. R. 1966. Cold
12. Margolin, P. See Ref. 9, 389-446
pring HarborSymp, Quant. Biol.
1:403~8
13. Umbarger,H. E. See Ref. 9, 447-63
14. Calvo, J. M.; Fink, G. R. 1971. Ann.
37. Guttman, B. S., Novick, A. 1963.
Rev. Bioehem.40:943-68
Cold Spring HarborSyrup. Quant.
15. Umbarger, H. E. 1969. Attn. Rev.
Biol. 28:373-74
Bioehem. 38:323-70
38. Kiho, Y., Rich, A. 1965. Proc. Nat.
16. Mtiller-Hill, B. 1971. Angew.Chem.
Acad. Sci. USA54:1751-58
lnt. Edit. 10:160-72
39. Martin, R. C. 1963. Cold Spring
17. Goldberger, R. F., Kovaeh, J. S.
Harbor Symp. Quant. Biol. 28:
Current Topics in Cellular Regula357-61
tion, ed. B. L. Horecker, E. R.
40. Imamoto,F., Morikawa,N.,Sato, K.
Stadtman. NewYork: Academic.
1965. J. Mol. Biol. 13:169-82
Inpress
41. Alpers, D. H., Tomkins,G. M. 1965.
18. Brenner, S., Jacob,.F., Meselson,M.
Proc. Nat. Acad. Sci. USA 53:
1961. Nature 190:576-80
797-803
19. Volkin, E., Astrachan,L. See Ref. 2,
42. Kepes, A. 1967. Biochim. Biophys.
686--94
Acm 138:107-23
20. Gros, F. et al. 1961. Nature 190:
43. Imamoto,F. 1968. Proc. Nat. Acad.
581-85
Sci. USA60:305-12
21. Leive, L.,Kolin, V. 1967. J. Mol.BioL 44. Baker, R. F., Yanofsky, C. 1968.
24:247-59
Proc. Nat. Acad, Sci. USA 60:
22. Kaempfer, R. O. R., Magasanik, B.
313-20
1967. J. Mol. Biol. 27:475-94.
45. Vogel, H. J. 1961. Cold Spring
23. Jacquet, M., Kepes, A. 1969.
Harbor Symp. Quant. Biol. 26:
Biochem. Biophys. Res. Commun.
163-72
36:84-92
46. Gorini, L., Gundersen, W., Burger,
24. Hayasbi, M., Spiegelman,S., FrankM. 1961. Cold Spring Harbor
ll’n, N. C., Luria, S. E. 1963.Proc.
Syrup. Quant. Biol. 26:173-82
47. Maas, W. K. 1961. Cold Spring
Nat. Acad. Sci, USA.49:729-36
25. Attardi, G., Naono,S., Rouvi~re,J.
Harbor Syrup. Quant. Biol. 26;
,lacob~ F, Gros~ F. 1963, CoM
!83-9!
~
Annual Reviews
www.annualreviews.org/aronline
Annu. Rev. Genet. 1972.6:133-156. Downloaded from arjournals.annualreviews.org
by University of Wisconsin - Madison on 03/29/07. For personal use only.
154
REZNIKOFF
48. Maas, W. K., Clark, A. J. 1964. J.
Mol. Biol. 8:365-70
49. Berberieh, M. A., Venetianer, P.,
Goldberger, R. F. 1966. J. Biol.
Chem. 241:4426-33
50. Berberich, M. A., Kovach, J. S.,
Goldberger,R. F. 1967. Proc. Nat.
Acad. ScL USA 57:1857-64
51, Jacob, F., Ullman, A., Monod,J.
1964. C. R. Acad. ScL 258:
3125-28
52. Epstein, W., Beckwith, J. R. 1968.
Ann. Reo. Biochem. 37:411-36
53. lppen, K., Miller, J. H., Scaife, J.,
Beckwith, J. 1968. Nature 217:
825-27
54. Miller, J. H., Ippen,K., Seaife, J. G.,
Beckwith,J. R. 1968. J. Mol. Biol.
38:413-20
55. Eron, L., Block, R. 1971. Proc. Nat.
Acad. ScL USA68:1828-32
56. Signer, E. R., Beckwith,J. R. 1966.
J. Mol. Biol. 22:33-51
57. Miller, J. H. et al. 1970.J. Bacteriol.
104:1273-79
58. Reznikoff, W.S., Beckwith, J. R.
1969. J. Mol. Biol. 43:215-18
59. Eron, L., Beckwith,J. R., Jacob, F.
See Ref. 5, 353-58
60. Chert, B. et al. 1971.NatureNewBiol.
233:67-70
61. Callahan, R., Blume, A. J., Balbinder, E. 1969. J. Mol. Biol.
51:709--15
62. Balbinder, E. et al. 1970. Genetics
66:31-53
63. Fankhauser, D. B. 1971. The promoter-operator region of the his
operon in Salmonella typhimuRIOM.Ph.D. thesis. The Johns
Hopkins Univ., Baltimore, 95 PP64. Blattner, F. R., Boettinger, J. K.,
Dahlberg, J. E. 1971. Fed. Proc.
30:1315 Abstr.
65. Blattner, F. R. et al. 1972.Fed. Proe.
31:472Abstr.
66. Smith, T. F., Sadler, J. R. 1971. J.
Mol. Biol. 59:273-305
67. Riggs, A. D., Reiness, C. G., Zubay,
G. 1971. Proe. Nat. Acad. Sci.
USA 68:1222-25
68. Emmer, M., deCrombrugghe, B.,
Pastan, I., Perlman,R. 1970.Proc.
Nat. Aead. Sci. USA66:480-87
69. Silverstone, A. E., Magasanik,B.,
Reznikoff, W.S., Miller, J. H.,
Beckwith,J. R. 1969. Nature 221:
1012-14
70. Perlman, R. L., deCrombrugghe,B.,
Pastan, I. 1969. Nature 223:
810-12
71. Beckwith, J. R., Grodzicker, T.,
Arditti, R. R. unpublishedexperiments
72. Silverstone, A. E., Arditti, R. R.,
Magasallik, B. 1970. Proc. Nat.
,4cad, Sci. USA66:773-79
73. Makmam,
R. S., Sutherland, E. W.
1965. J. Biol. Chem.240:1309-14
74. Perlman, R. L., Pastan, I. 1968.
Biochem. Biophys. Res. Commun.
30:656-64
75. Ullmann, A. Monod,J. 1968. Fed.
Eur. Biol. Soc. Lett. 2:57-60
76. Schwartz, D., Beckwith, J. R. See
Ref. 5, 417-22
77. Perlman, R. L., Pastan, I. 1969.
Biochem. Biophys. Res. Commun.
37:151-57
78. Zubay, G., Schwartz, D., Beckwith,
J. 1970. Proc. Nat..4cad. Sci. USA
66:104-10
79. de Crombrugghe,B. et al. 1971. Nature NewBiol. 231:139-42
80. Voll, M. J. 1967. J. Mol. Biol. 30:
109-24
81. Margolin, P. 1963. Genetics 48:44156
82. Miiller-Hill, B, Crapo, L., Gilbert,
W. 1968. Proc. Nat. Acad. Sci.
USA 59:1259-64
83. Fraenkel,
D. G.,56:183-94
Banergee, S. 1971.
J. MoLBiol.
84. Miller, J. H., Beckwith,J., Mi~llerHill, B. 1968. Nature 220:1287-90
85. Kumar, S., Szybalski, W. 1969. J.
MoLBiol. 40:145-51
86. Reznikoff,W.S., Miller, S. H., Scaife,
J. G., Beckwith,J. R. 1969. J. MoL
Biol. 43:201-13
87. Reznikoff, W. S. Unpublished experiments
88. Mitchell, D., Reznikoff, W.S., Beckwith, J. R., Michels, C. A., Unpublished experiments
89. Rose, J. K., Yanofsky, C. 1971. J.
Bacteriol. 108:615-18
90. Roberts, J. W. 1969. Nature 224:
1168-74
91. Nissley, S. P., Anderson, W. B.,
Gottesman,M. E., Perlman, R. L.,
Pastan, I. 1971.J. Biol. Chem.246:
4671-78
92. Gilbert, W., Miiller-Hill, B. 1967.
Proc. Nat. Acad. Sci. US.4. 58:
2415-21
93. Riggs, A. D., Newby,R. R., Bourgeois, S., Cohn, M. 1968. J. MoL
Biol. 34:365-68
94. Parks, J. S. et al. 1971. Proc. Nat.
Acad. Sci. USA68:1891-95
95. Ptashne, M. 1967. Nature 214:232-34
96. Pirrotta, V., Ptashne, M. 1969. Nature 222:541-44
Annual Reviews
www.annualreviews.org/aronline
Annu. Rev. Genet. 1972.6:133-156. Downloaded from arjournals.annualreviews.org
by University of Wisconsin - Madison on 03/29/07. For personal use only.
THE OPERON REVISITED
97. Lavall~, R., DeHauwer,G. 1970. J.
MoLBiol. 51:435-47
98. Hsu, W., Weiss, S. B. 1969. Proc.
Nat. Acad. Sci. USA64:345-51
99. Steitz, J. A., Duke,S. K., Rudlaad,
P. S. 1970. Nature226:824-27
I00. Pollack, Y., Groner, Y., Aviv, H.,
Revel, M. 1970. Fed. Eur. Biol.
Soc. Left. 9:218-21
101. Revel, M., Aviv, H., Groner, Y.,
Pollack, Y. 1970. Fed. Eur. Biol.
Scl. Lett. 9:231-17
102. Berissi, H., Groner, Y., Revel, M.
1971, Nature NewBiol. 234:44-47
103. Lee-Huang,S., Ochoa, S. 1971. Nature NewBiol. 234:236-39
104. Grunberg-Manago,M., Rabinowitz,
J. C., Dondon,J., Lelong, J. C.,
Gros,
1971. Fed. Eur. Biol. Soc.
Lett. F.
19:193-200
105. Vermeer, C., Talens, I, BloemsmaJonkman,F., Bosch, L., Davies,
J.
W.19:201-06
1971. Fed. Eur. BioL Soc.
Lett.
106. Monod,J., Cohen-Bazire, G., Cohn,
M. 1951. Biochim. Biophys. Acts
7:585-99
107. Lederberg, J. 1951. Genetics in the
20th Century,ed. L. C. Durra, 26389 NewYork: MacMillan. 634 pp.
108. Cohen-Bazire, G., Jolit, M. 1953.
Ann. Inst. Pasteur. 84:940-45
109. Pardee, A. B., Jacob, F., Monod,J.
1958. C.R. Acad. ScL 246:3125-27
110. Willson, C., Perrin, D., Cohn, M.,
Jacob, F., Monod,J. 1964. J. Mol.
Biol. 8:582-92
111. Sadler, J., Novick,A. 1965. J. Mol.
BioL 12:305-27
112. Bourgeois, S., Cohn, M., Orgel,
L. E. 1965. J. Mol. Biol. 14:300-02
113. Barbour, S. C., Pardee, A. B. 1966.
J. Mol.Biol. 20:505--15
114. Horiuchi, T., Ohshima,Y. 1966. J.
MoLBiol. 20:517-26
115. Gilbert, W., Mtiller-Hill, B. 19f6.
Proc. Nat. Acad. Sci. USA 56:
1891-98
116. Riggs, A. D., Bourgeois,S. 1968. J.
Mol. Biol. 34:361-64
117. Riggs, A. D., Suzuki, J., Bourgeois,
S. 1970. J. Mol.Biol. 48:67-83
118. Miiller-Hill, B., Beyreuther,K., Gilbert, W. 1971. Methods Enzymol.
21:483-87
119. Bourgeois, S., Jobe, A. See Ref. 5,
325-41
120. Davies, J., Jacob, F. 1968. J. MoL
Biol. 36:413-17
121. Miiller-Hill, B., Crapo, L., Gilbert,
W. 1968. Proc. Nat. Acad. Sci.
USA 59:1259-64
155
122. Markovitz,A. 1964. Proc. Nat. Acad.
Sci. USA51:239-46
123. Chater, K. F. 1970. J. Gen. Microbiol. 63:95-109
124. Hoar, D. I. 1971. J. BacterioL 108:
582-83
125. Corredor, J., Reznikoff, W.S. 1972.
Abstr. Ann. Meet. Am. Soc. MicrobioL 72:52
126. Roth, J. R., Ant6n, D. N., Hartman,
P. E. 1966. J. Mol. Biol. 22:305-23
127. Ant6n,D. N. 1968. J. Mol. Biol. 33:
533-46
128. Lewis, J. A., Ames,B. N. 1972, J.
Mol.Biol. In press
129. Roth, J. R., Ames, B. N. 1966. J.
Mol. Biol. 22:325-34
130. Silbert, D. F., Fink, G. R., Ames,
B. N. 1966. J. Mol.Biol. 22:335-47
131. Singer, C. E., Smith,G. R., Cortesse,
R. Ames, B. N. 1972. Nature In
press
132. Englesberg, E., Irr, J., Power, J.,
Lee, N. 1965. J. Bacteriol. 90:94657
133. Schwartz, M. 1967. Ann. Inst. Pasteur 112:673-700
134. Power, J. 1967. Genetics 55:557-68
135. Lindahl, G. 1970. Virology 42:522-33
136. Smith, G. R., Magasanik, B. 1971.
Proc. Nat. Acad. Sci. USA68:
1493-97
137. Sly, W.S., Rabideau,K., Kolber, A.
1971. The Bacteriophage X, ed.
A. D. Hershey, 575-88. Cold
Spring Harbor, N.Y.: Cold Spring
Harbor Laboratory. 792 pp,
138. Wyche,J. H. 1971. Histidyl-tRNA
synthetase mutants and regulation
in the histidine operon of Salmonella typhimurium. Ph.D. thesis.
Johns Hopkins Univ., Baltimore.
87 pp.
139, Kovaeh, J. S., Berberich, M. A.,
Venetianer, P., Goldberger, R. F.
1969. J. Bacteriol. 97:1283-90
140. Kovach,J. S., Phang,J. M., Ference,
M., Goldberger, R. F. 1969. Proc.
Nat. Acad. ScL USA63:481-88
141.Kovach,J. S. et al. 1970.J. Bacteriol.
104:787-92
142. Sadler, J. R., Smith, T. F. 1971. J.
Mol, Biol. 62:139-69
143. Bhorjee,J. S., Fowler,A. V., Zabin,
I. 1969. J. Mol. Biol. 43:219-22
144. Steers, E., Craven,G. R., Anfinsen,
C. B. 1965. Proc. Nat. ,4cad. Sci.
USA54:1174-81
145. Englesberg, E., Squires, C., Meronk,
F. 1969. Proc. Nat..4cad. Sci. USA
62:1100-07
Annual Reviews
www.annualreviews.org/aronline
Annu. Rev. Genet. 1972.6:133-156. Downloaded from arjournals.annualreviews.org
by University of Wisconsin - Madison on 03/29/07. For personal use only.
156
REZNiKOFF
146. Gielow, L., Largen, M., Englesberg,
E. 1971. Genetics 69:289-302
I47. Hofnung, M., Schwartz, M. 1971.
Mol. Gen. Genet. 112:117-32
148. Englesberg,E. Sheppard,D., Squires,
C., Meronk,F. 1969. J. Mol. Biol.
43:281-98
149. Wilcox,G., Clemetson,K. J., Santi,
D. V., Englesberg, E. 1971. Proc.
Nat. Acad. Sci. USA68:2145-48
150. Zubay, G., Gielow, L., Englesberg,
E. 1971. Nature New Biol. 233:
164-65
151. Greenblatt, J., Schleif, R. 1971. Nature NewBiol. 233:166-70
152. Burstein, C., Cohn, M., Kepes, A.,
Monod,J. 1965. Biochim. Biophys.
Acta 95:634-39
153. Jobe, A., Bourgeois, S. 1972. J. MoL
Biol. In press
154. Eidllc, L., Neldhardt, F. C. 1965.
Proc. Nat. Acad. Sci. USA53:53943
155. Freundlich, M. 1967. Science 157:
823-25
156. Williams, L. S., Freundlich, M. 1969.
Biochim. Biophys. Acta 186:30516
157. Pi6rard, A., Glandorff, N., Mergeay,
M., Wiame, J. M. 1965. J. Mol.
Biol. 14:23-36
158. Freundlich, M., Burns, R. O., Umbarger, H. E. 1962. Proc. Nat.
Acad. Sci. USA48:1804-08
159. Freundlich, M. 1963. Biochem.Biophys. Res. Commun.10:277-82
160. Szentirmai, A., Szentirmai, M,, Umbarger, H. E. 1968. J. Bacteriol.
95:1672-79
161. Dwyer,S. B., Umbarger,H. E. 1968.
J. Bacteriol. 95:1680-84
162. Calvo, J. M., Fretmdlich, M., Umbarger, H. E. 1969. J. Bacteriol.
97:1272-82
163. Brown,K. D., Doy, C. H. 1966. Biochim. Biophys. Acta 118:157-72
164. Brown,K. D. 1968. Genetics 60:3148
1.65. Brown, K. D., Somerville, R. L.
1971. J. Bacteriol. 108:386-99
I66. rm,s. w. K., Davidson,H., Pittard,
J. 1971. J. Bacteriol. 108:400-09
167. Buttin, G. 1963. J. Mol. Biol. 7:183205
168. Mackie,G., Wilson, D. B. 1971. Fed.
Proc. 30:1262 Abstr.
169. Arfin, S. M., Ratzkin, B., Umbarger,
H. E. 1969. Biochem.Biophys. Res.
Commun.37:902-08
170. Zillig, W.et al. 1970. Cold Spring
Harbor Symp. Quant. Biol. 35:
47-58
171. Lozeron, H. A., Funderburgh,M. L.,
Dahlberg, J. E., Stark, B. P.,
Szybalski, W. 1972. Abstr. Ann.
Meet. Am. Soc. Microbiol. 72:237
171a. Wu,A. M., Ghosh, S., Echols, H.
1972.J. Mol.Biol. In press
172. Riggs, A. D., L/n, S., Wells, R. D.
1972. Proc. Nat. Acad. Soc. USA
69:761-64
173. Pfahl, M. 1971. Kartierungand charakterisierung yon lac-i-genmutanten
in Escherichia coll. Ph.D. thesis.
Univ. Koln, Germany.67_pp.
174. Adler, K. et al. 1972.NatureIn press.
174a. Platt, T., Weber, K., Ganem,D.,
Miller, J. H. 1972Proc.Nat. Acad.
Sci. USAIn press
175. yon Hippel, P. H. 1969. J. Cell
Physiol. 74: Suppl, 1 235-38
176. Wilkins, M. H. F. 1957. Biochem.
Soc. Syrup. 14:13-25
177. Kelly, T, J., Smith, H. O. 1970. J.
Mol. Biol. 51:393-409
178. Printz, M. P, yon Hippel, P. H.
1965. Proc. Nat. Acad. Sci. USA
53:363-70
179. Gierer, A. 1966. Nature 212:1480-81
180. Wells, R. D., Larson, J. E., Grant,
R. C., Shortle, B. E., Cantor,
D. R. 1970. J. Mol. Biol. 54:465-97
181. H61~ne,C. 1971. Nature NewBiol.
234:120-21
182. Sundaraligam, M., Arora, S. K.
1969. Proc. Nat. Acad. Sci. USA
64:1021-26
Annu. Rev. Genet. 1972.6:133-156. Downloaded from arjournals.annualreviews.org
by University of Wisconsin - Madison on 03/29/07. For personal use only.