Science of the Total Environment 463-464 (2013) 42–50
Contents lists available at SciVerse ScienceDirect
Science of the Total Environment
journal homepage: www.elsevier.com/locate/scitotenv
Riparian swallows as integrators of landscape change in a multiuse
river system: Implications for aquatic-to-terrestrial
transfers of contaminants
Jeremy M. Alberts a,⁎, S. Mažeika P. Sullivan b, A. Kautza b
a
b
Department of Biological Sciences, University of Cincinnati, 2600 Clifton Ave., Cincinnati, OH 45221, United States
School of Environment & Natural Resources, The Ohio State University, 2021 Coffey Rd., Columbus, OH 43210, United States
H I G H L I G H T S
•
•
•
•
We characterized the land use and land cover (LULC) of 11 riverine study sites.
We examined selenium (Se) and mercury (Hg) concentrations in riparian swallows.
Relationships were evident between LULC and contaminant concentrations.
Contaminant concentrations were higher in adult than in juvenile birds.
a r t i c l e
i n f o
Article history:
Received 21 January 2013
Received in revised form 12 May 2013
Accepted 13 May 2013
Available online xxxx
Editor: Christian E.W. Steinberg
Keywords:
Riparian swallows
Biomagnification
Aquatic–terrestrial linkages
a b s t r a c t
Recent research has highlighted the transfer of contaminants from aquatic to terrestrial ecosystems via predation of
aquatic emergent insects by riparian consumers. The influence of adjacent land use and land cover (LULC) on
aquatic-to-terrestrial contaminant transfer, however, has received limited attention. From 2010 to 2012, at 11
river reaches in the Scioto River basin (OH, USA), we investigated the relationships between LULC and selenium
(Se) and mercury (Hg) concentrations in four species of riparian swallows. Hg concentrations in swallows were significantly higher at rural reaches than at urban reaches (t = −3.58, P b 0.001, df = 30), whereas Se concentrations
were positively associated with adjacent land cover characterized by mature tree cover (R2 = 0.49, P = 0.006). To
an extent, these relationships appear to be mediated by swallow reliance on aquatic emergent insects. For example,
tree swallows (Tachycineta bicolor) at urban reaches exhibited a higher proportion of aquatic prey in their diet, fed at
a higher trophic level, and exhibited elevated Se levels. We also found that both Se and Hg concentrations in adult
swallows were significantly higher than those observed in nestlings at both urban and rural reaches (Se: t = −2.83,
P = 0.033, df = 3; Hg: t = −3.22, P = 0.024, df = 3). Collectively, our results indicate that riparian swallows integrate contaminant exposure in linked aquatic–terrestrial systems and that LULC may strongly regulate aquatic
contaminant flux to terrestrial consumers.
© 2013 Elsevier B.V. All rights reserved.
1. Introduction
Movement of materials across the landscape has been central to the
study of food webs (Polis et al., 1997, 2004). At the aquatic–terrestrial
interface, ecological subsidies, or the flux of material from one habitat
or ecosystem to another (Nakano and Murakami, 2001) has emerged
as a central theme of cross-boundary linkages (Akamatsu et al., 2004;
Vander Zanden and Sanzone, 2004; Ballinger and Lake, 2006). Whereas
the contribution of terrestrial systems to aquatic food webs via inputs of
nutrients and organic matter is well recognized, recent investigations
have focused on the importance of aquatic prey subsidies to the energy
budgets of riparian consumers (reviewed in Baxter et al., 2005).
⁎ Corresponding author. Fax: +1 513 556 5299.
E-mail address: albertjy@mail.uc.edu (J.M. Alberts).
0048-9697/$ – see front matter © 2013 Elsevier B.V. All rights reserved.
http://dx.doi.org/10.1016/j.scitotenv.2013.05.065
A classic example is the reliance of grizzly bears (Ursus arctos horribilis)
on fish as an important nutrient source (Mattson and Reinhart, 1995;
Varley and Schullery, 1996). Others have shown that aquatic prey
represent important food sources to riparian bats (Fukui et al., 2006;
Hagen and Sabo, 2012), raccoons (Procyon lotor) (Bigler et al., 1975;
Lord et al., 2002), piscivorous birds (Vessel, 1978; Hamas, 1994), lizards
(Sabo and Power, 2002), and terrestrial arthropods (Henschel et al.,
2001; Paetzold et al., 2005; Dreyer et al., 2012).
In particular, aquatic insects that emerge from the stream as adults
(hereafter, ‘aquatic emergent insects’) can be a critical food source for
insectivorous riparian bird species (Keast, 1990; Iwata et al., 2003).
For example, Sweeney and Vannote (1982) found that swarming
activity by mayflies attracted nighthawks (Chordeiles minor) to areas
of emergence, and Iwata et al. (2003) reported that flycatchers and
gleaners foraged intensively on aquatic emergent insects in a pattern
J.M. Alberts et al. / Science of the Total Environment 463-464 (2013) 42–50
that mirrored aquatic insect distribution, suggesting that the dynamics
of insect emergence can influence avian foraging behavior.
Within the context of the energetic pathways that link aquatic and
terrestrial ecosystems, there is growing interest in food-web studies
at the land–water ecotone. Many investigators have found that the
high reliance of riparian birds on aquatic prey resources can expose
them to contaminants found in aquatic systems (see Sullivan and
Rodewald, 2012). Cristol et al. (2008) demonstrated that mercury
(Hg) can bioconcentrate in terrestrial bird species living near contaminated water sources, while Wada et al. (2009) found that adrenocortical responses and thyroid hormones were suppressed in tree
swallows (Tachycineta bicolor) living near a river contaminated
with mercury. In the Chilliwack River of southern British Columbia,
resident American Dippers (Cinclus mexicanus) (i.e., those birds who
remain in the mainstem Chilliwack River year-round and have high
proportions of contaminated salmon fry in their diet) were found to
have higher polychlorinated biphenyl (PCB) and Hg concentrations
than their migratory counterparts (i.e., those birds who are found
on the Chilliwack mainstem only during the winter and that have
a significantly smaller proportion of fish fry in their diet; Morrissey
et al., 2004). Several studies have described the bioconcentration of
PCBs in tree swallows from aquatic insect prey (Echols et al., 2004;
Maul et al., 2006). Echols et al. (2004), for instance, found that PCB
concentrations in tree swallow nestlings closely tracked concentrations in aquatic insects. Maul et al. (2006) implicated aquatic insects
of the family Chironomidae as the key vector of PCBs to nestling
tree swallows from a lake in Illinois, USA.
Differences in age-related contaminant exposure patterns among
birds have also been documented. For example, Malinga et al. (2010)
noted a positive correlation between age and biomagnification of
contaminants in Glaucus gulls (Larus hyperboreus) in the Arctic. Likewise, Evers et al. (2005) found that concentrations of methylmercury
(MeHg) in adult tree swallows were higher than in nestlings. The authors suggest that differences in prey intake between adult and juvenile
birds, as well as the ability of young to transfer MeHg from blood into
feathers during rapid growth, may account for such age-based variation.
Given the widespread contamination of aquatic systems
(Fitzgerald et al., 1998; Hammerschmidt and Fitzgerald, 2005;
Blocksom et al., 2010), the amount of currently available information on aquatic-to-terrestrial contaminant fluxes does not reflect
the potential scale of the problem. In particular, although the impacts
of land use and land cover (LULC) on riparian birds have been widely investigated (Sullivan et al., 2007; Luther et al., 2008; Pennington et al.,
2008; Oneal and Rotenberry, 2009), the relationships between LULC
and contaminant exposure to riparian birds from aquatic systems is
poorly understood. For example, riparian swallow species [e.g., bank
(Riparia riparia), northern rough-winged (Stelgidopteryx serripennis),
tree, and cliff (Petrochelidon pyrrhonota) swallows] are susceptible
to biomagnification of contaminants because of their dependence on a
mixture of stream, riparian, and terrestrial food sources and habitats
(Harris and Elliot, 2000; Dods et al., 2005; Custer, 2011). During the
reproductive season, riparian swallows tend to forage within several
hundred meters (m) of their nest sites (Quinney and Ankney, 1985;
Dunn and Hannon, 1992), making them apt bioindicators of local
stream and riparian conditions. Land-management practices that alter
aquatic resource utilization by swallows might be expected to strongly
influence their exposure to aquatic contaminants. Roux and Marra
(2007), for instance, found that lead (Pb) concentrations in urban
passerine birds were significantly higher than in their rural counterparts,
which corresponded with relative Pb concentrations found in soils.
Our objective was to explore the relationships between adjacent
(to the river) patterns in LULC and the magnitude of Hg and Se
concentrations in four species of riparian swallows as mediated by
aquatic-to-terrestrial contaminant fluxes. Because of historic and
contemporary differences in land-management and chemical use, we
hypothesized that concentrations of contaminants in river sediments,
43
aquatic emergent insects, and riparian swallows would be greater at
urban than at rural reaches. Consequently, we anticipated that contaminant exposure to swallows would also differ by LULC class, but would
be mediated by the degree of aquatic resource utilization such that
swallows feeding at rural reaches (i.e., comprised of a mixture of forest,
grassland, and agriculture) would exhibit greater reliance on aquatic
emergent insects than swallows feeding at urban reaches, where aquatic
emergent insect communities have been shown to exhibit reduced
diversity (Stepenuck et al., 2002) and biomass (Lenat and Crawford,
1994; Meyer and Sullivan, in press). Finally, we expected that adult
swallows would exhibit greater biomagnification of contaminants than
nestlings at both urban and rural reaches.
2. Materials and methods
2.1. Study system and experimental design
The Scioto River is a multiuse river that drains ~ 16,882 km2 in
central and southern Ohio, flowing for 372 km through agricultural
(69%), forested (21%), and urban (9%) landscapes (White et al.,
2005; Blocksom and Johnson, 2009). The Olentangy River meets the
Scioto in Columbus, the 15th largest city in the US by population
(1,178,899 people; USCB, 2010). Once downstream of the city, the
Scioto is free-flowing and exhibits riparian zones dominated by deciduous
forest, row crops (corn and soybean), and smaller urban centers.
From 2010 to 2012, we conducted coordinated sampling of riparian swallows, riverine sediment and aquatic insects, and aquatic and
riparian vegetation at eleven river reaches [defined as a river segment based on similar valley features and channel geomorphology
(~ 1 kilometer [km] for this study)] of the Scioto River watershed,
representing a gradient of urban to rural (i.e., mixture of forest,
grassland, and agriculture) landscapes (Fig. 1). Study reaches were
distributed along the length of the river and were separated by an
average distance of 18.4 river kilometers (km), although there was
significant variability (SD = 16.8 km). Because of the spatial distribution of LULC patterns in the basin, a degree of clustering among
LULC types was unavoidable. We used ArcGIS 10.1 (ESRI, Redlands,
CA, USA) to calculate percent cover by vegetation class within
500 meters (m) of the channel following Zelt and Johnson (2005),
and performed ground-based surveys to document bridge crossings,
vegetation structure, and vegetation composition.
2.2. Field and laboratory methods
2.2.1. Sediment
In either 2010 or 2012, we collected five sediment cores (~ 10 cm
in depth) from each reach at approximately equidistant locations
along the length of the reach (Walters et al., 2010). We collected
sediment cores by push-coring approximately 2 cm with a polycarbonate
coring device. We sealed sediment composite samples in plastic sleeves,
placed them on ice, and froze them at −20 °C until analysis (Lutz, 2008).
2.2.2. Periphyton and vegetation
Following methods used in Bartel et al. (2010) and Pennington
and Blair (2011), we surveyed riparian vegetation along three 100-m
transects (upstream, middle, downstream) along both banks (perpendicular to the stream, n = 6/reach) at each reach. At each transect, we
established three 5 m × 5 m quadrats, located at 0, 50, and 100-m
from the river's edge. We then estimated vegetation cover by height
(b3 m, 3–5 m, and >5 m) and identified all trees, shrubs, grasses,
and other herbaceous plants to species. We visually estimated the
percent coverage of vegetation overhanging the water's edge along
both banks and counted standing dead trees of diameter at breast
height (dbh) > 10 cm.
For stable isotope analysis, we collected five terrestrial vegetation
samples per transect on both sides of the river (along survey transects):
44
J.M. Alberts et al. / Science of the Total Environment 463-464 (2013) 42–50
Fig. 1. Eleven study reaches of the Scioto River basin, Ohio, USA. Open circles in popouts indicate study reaches. Circles in right-hand popout represent nest box locations.
Popouts are not to scale.
three samples from the numerically dominant tree species and two
samples from the dominant species among the shrubs, grasses, herbs
and sedges. We stored leaf samples in paper bags until further processing in the laboratory. We collected periphyton by scrubbing a 25-cm2
section of the dominant hard substrate (cobble) at each of the five
transects within each reach following Reavie et al. (2010). The samples
were composited by reach into a single polyethylene jar and preserved
in 70% ethanol solution until analysis.
2.2.3. Riparian swallows
We selected a suite of riparian swallow species (tree, cliff, bank,
and northern rough-winged swallows) that were common to the
study system. These swallow species are obligate aerial insectivores
that naturally breed in riparian habitats (riparian tree cavities,
riverbanks, cliffs and bridges, etc.) (Robertson et al., 1992; Brown and
Brown, 1995; DeJong, 1996; Garrison, 1999). Given potential differences in species-specific exposure patterns to contaminants, we ideally
would have focused our efforts on one riparian swallow species.
However, the distribution of swallow species in the study system
precluded this option, as there was no one species of swallow found
at all the study reaches. Others have also used a foraging guild-based
approach to investigate contaminant exposure in birds (e.g., Jackson
et al., 2011). We conducted all sampling during the swallow reproductive season (May–early August in 2011 and 2012).
At seven of the eleven research reaches, we captured swallows using
mist nets set over the river or in the riparian zone. We erected six nest
boxes at each of four additional reaches, following design protocols set
by the Golondrinas de las Américas (2011) project at Cornell University.
All nest boxes were placed within or along the edge of the riparian zone
and were within 100 m from the river on the average. We equipped
each box with an external trapping mechanism (i.e., wig-wag), which
allowed the capture of adult tree swallows while they were inside the
nest box. At first capture, we banded all birds with aluminum USFWS
size 0 bands for tree, bank, and northern rough-winged swallows, and
size 1 bands for cliff swallows. We drew blood from the jugular vein
to be used for stable isotope and contaminant analyses (Ardia, 2005;
Sullivan and Vierling, 2012). We collected blood from a minimum of
three adults and four nestlings during the second week after hatching
at each of the experimental nest box reaches, where at least two
nests were sampled per reach. At all other reaches, variability in
the accessibility and abundance (i.e., colony-nesting cliff swallows
at some reaches, couple/few pairs of northern rough-winged swallow at others) of swallows constrained sample size. However, we
collected blood from a minimum of four swallows at all non-nest
box reaches, except at the study reach “Mosquito”, where we were
only able to capture three birds. Once drawn, we immediately transferred blood samples to centrifuge tubes where it was stored in 70%
ethanol for later stable isotope analysis. We placed additional blood
on Dried Blood Spot cards (PerkinElmer, Waltham, MA, USA) for
contaminant analysis. DBS cards are an appropriate technique
to sample blood from small-bodied animals (b20 g; mean weight
of birds in our study at collection was 16.6 g, SD = 3.8 g), from
which adequate sample volume can preclude traditional methods
(Shlosberg et al., 2011).
2.2.4. Invertebrates
Adult tree swallows typically return to the nest with a bolus of
invertebrate prey carried in their beaks, which we removed from their
mouths before the parent fed the nestlings at nest box reaches
(McCarty and Winkler, 1999; Echols et al., 2004). At all other reaches,
we deployed six floating aquatic emergent insect traps (i.e., pyramid
traps) per reach as described by Aubin et al. (1973). We deployed
traps along both banks of the reach such that they represented
dominant flow habitats (i.e., riffle, pool/run). For terrestrial insects, we
suspended half-sized Malaise traps (1-m high, 1-m long, 0.6-m wide,
0.5-mm mesh, MegaView Science Co., Taichung, Taiwan; Townes,
1972) from trees at 10 m and 50 m from the active river channel approximately in the middle of the study reach, on both banks. We collected all
invertebrates five and ten days after trap deployment.
Upon collection, we placed all invertebrates in jars, put them
on ice for transport, and froze them at − 20 ° C until analysis (Lutz,
2008). In the laboratory, we identified all individuals to family
using Johnson and Triplehorn (2004). For bolus invertebrates, we
also classified all individuals by their origin (i.e., aquatic or terrestrial
according to their larval habitat). Following identification, we dried
and weighed invertebrates by family (total dry mass per reach
[mg m− 2 d− 1]; Iwata et al., 2003).
J.M. Alberts et al. / Science of the Total Environment 463-464 (2013) 42–50
2.2.5. Contaminant and stable isotope analysis
All sediment, bolus, and swallow blood samples were analyzed for
Hg and Se concentrations [parts per billion (ppb) dry weight] at
the Diagnostic Center for Population and Animal Health, Toxicology
Section, Michigan State University. ICP-AES (Varian Vista, now part
of Agilent, Santa Clara, CA) and ICP-MS (Agilent 7500ce) instruments
were calibrated with standards derived from NIST-traceable stock solutions for each element (GFS Chemicals, Inc., Cincinnati, OH). Quality
was assured in each sequence run by analyzing lab reagent blanks,
NIST (National Institute of Standards & Technology, Gaithersburg,
MD)-traceable Multi-mix (Alfa Aesar Specpure, Ward Hill, MA) and
digests of NIST Standard Reference Materials (SRM), including Mussel
Tissue 2976, Trace Elements in Water 1643e and/or Montana II Soil
2711a, as appropriate. Mercury was analyzed by cold vapor atomic
absorption spectrometry (CETAC CVAA, Omaha, NB) with similar
calibration and QC. Analysis of blood was performed by methods
outlined in Lehner et al. (in press) with reference to BioRad (Hercules,
CA) Lypochek 2 and 3 standards for verification of accuracy.
Terrestrial vegetation, terrestrial insects (Curculionidae, Dolichopodidae, and Cicadellidae), periphyton, aquatic emergent insects
(Chironomidae, Hydropsychidae, and Heptageniidae), and bird blood
samples were analyzed for 13C and 15N stable isotopes using continuous
flow isotope ratio mass spectrometry (EA-IRMS) at the University of
Washington Stable Isotope Core (Pullman, WA, USA). The isotopic
composition of samples was expressed using the δ13C notation
defined as follows:
13
δ C¼
h
i
Rsample =Rstandard –1 1000; where R is
13
12
C= C
ð1Þ
where parts per thousand (‰) are the atomic ratios of the number
of atoms in the sample or standard. Typical analytical precision
was +/- 0.2‰ for δ13C determination.
2.2.6. Numerical and statistical analyses
Because of the largely longitudinal distribution of our study reaches
along the river system, we considered testing for spatial autocorrelation
of all measured variables. However, the focus of our study was on the
role terrestrial environmental characteristics played in mediating fluxes
of contaminants to riparian consumers, and not on the distribution of
river contaminants themselves. Therefore, we used Moran's I (1950)
statistic (Cliff and Ord, 1973) to describe spatial patterns of swallow
contaminant concentrations only. Positive and significant (P b 0.05)
Moran's I values indicate a more spatially-clustered distribution than
would be expected if the underlying spatial processes were random,
whereas negative values indicate greater spatial dispersion.
Because we observed no differences in δ13C and δ15N values between adult and nestling swallows based on paired t-tests, we grouped
nestlings and adults by species for each study reach. We then analyzed
the contributions of aquatic and terrestrial food sources to riparian
swallows at each study reach using (1) dual isotope plots of δ13C and
δ15N, (2) linear regression between δ13C of swallows and periphyton
(i.e., gradient approach; Jardine et al., 2012), and (3) invertebrates
from bolus samples. For dual-isotope plots, spatial correspondence
between δ13C and δ15N values in swallows and aquatic prey indicates
greater use of aquatic prey (DeNiro and Epstein, 1978, 1981). We created
one plot for each land-use class (i.e., urban and rural) and plotted δ13C
and δ15N values for swallows by species, given potential variability in
isotopic signatures among species. Because of an unbalanced number
of swallows at each reach, we used weighted linear regression to determine the degree to which swallow δ13C tracked periphyton δ13C across
all study reaches. We used invertebrate composition from bolus samples
to further inform aquatic:terrestrial prey use by tree swallows.
We performed principal component analysis (PCA) on riparian
LULC variables that we had selected a priori as candidate predictors
of riparian swallow prey utilization. PCA axes with eigenvalues > 1
45
were retained for use in subsequent linear regression models
(Rencher, 1995; Sullivan and Watzin, 2008). We used weighted linear
regression to examine the best LULC predictors of contaminant
concentrations in swallows. We used two-sample t-tests to test for
potential differences in (1) the contribution of aquatic resources to
swallows and (2) contaminant concentrations in sediment, aquatic
insects, and riparian swallows between LULC types. We used paired
t-tests to test for differences between contaminant concentrations
in (1) adult and nestling swallows and (2) aquatic and terrestrial insects from bolus samples. To avoid potential pseuodreplication at nest
box reaches where multiple nestlings were sampled from the same
box, mean contaminant concentrations of nestlings per nest box
were used in the analysis. Data were tested at the α = 0.05 level.
However, given multiple regression tests, the Bonferroni adjustment
for α was α / k = 0.05 / 5 = 0.010, where k is the number of tests
(Wright, 1992). Prior to the analysis, concentrations of Se and Hg
were log10 were transformed to normalize data and eliminate
heteroscedasticity (Zar, 1984).
3. Results
Sediment concentrations of Se ranged from 367 to 2440 ppb,
while sediment Hg ranged from 31 to 137 ppb. Neither Se (t =1.14,
P = 0.327, df = 7) nor Hg (t = 0.157, P = 0.881, df = 7) in sediment
was significantly different between urban and rural reaches. Likewise,
neither Se (t = 1.77, P = 0.160, df = 7) nor Hg (t = 0.31, P = 0.766,
df = 7) in aquatic emergent insects was different between urban and
rural reaches. Concentrations of Se and Hg in riparian swallows varied
widely across study reaches (Supplemental Data Table 1). Hg concentrations in swallows were positively related to Hg concentrations
in sediment (Fig. 2), although no relationship was evident between Se in swallows and in sediment. Se concentrations were
greater in aquatic (mean = 3956 ppb, SD =2039) than in terrestrial (mean = 1132 ppb, SD = 1089) invertebrate swallow prey
(t = − 4.11, P = 0.027, df = 2). A similar, but weaker relationship
was found for Hg (aquatic: mean = 66.3 ppb, SD = 28.3, terrestrial:
mean = 32.7 ppb, SD = 10.8; t = −2.17, P = 0.081, df = 2). Moran's
I indicated that there was no spatial clustering of swallow contaminant
concentrations (P > 0.05 in all cases), suggesting that observations
from nearby study reaches were independent from one another.
Fig. 2. Relationship between Hg concentrations (log10) in riparian swallows and sediment
(log10) (ppb). The slope for the regression model was significant (y = 0.39x + 3.25,
R2 = 0.33, P = 0.008). Two data points (open circles) were considered outliers and omitted
from the analysis, although the regression was also significant with these two points included
(y = 0.46x + 2.95, R2 = 0.23, P = 0.030). Dashed lines represent confidence curves at
α = 0.05.
46
J.M. Alberts et al. / Science of the Total Environment 463-464 (2013) 42–50
3.1. Contribution of aquatic resources to swallow diet
Isotopic values (Table 1) for riparian swallows ranged from −26.77
to −20.29‰ for δ13C and from 6.63 to 12.74‰ for δ15N. Signatures for
aquatic emergent insect δ13C ranged from −28.27 to −27.04‰, while
δ15N ranged from 10.04 to 12.31‰. Terrestrial insect δ13C ranged from
−25.84 to −22.47‰ whereas δ15N of terrestrial insects ranged from
5.10 to 8.74‰. Terrestrial vegetation δ13C ranged from − 31.53 to
− 28.00‰, and δ15N from 2.72‰ to 4.68‰. Periphyton exhibited
the greatest variation, ranging from − 21.46 to − 11.26‰ for δ13C,
and from 6.14 to 12.39‰ for δ15N. We found no significant differences for either δ13C (t = − 0.12, P = 0.455, df = 3) or δ15N
(t =1.33, P = 0.137, df = 3) between adult (δ13C: mean = −24.2,
SD = 0.99; δ15N: mean = 10.1, SD = 1.35) and juvenile (δ13C:
mean = −24.4, SD = 0.72; δ15N: mean = 10.4, SD = 1.06) swallows,
although the sample size was small (n = 10).
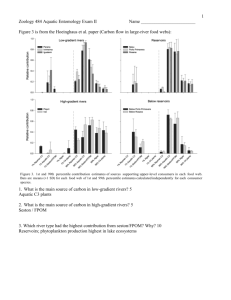
Swallows at both rural and urban reaches are plotted within the
stream–riparian food web (Fig. 3a & b). For rural (Fig. 3a) and urban
(Fig. 3b) reaches, isotope plots indicated that swallows were reliant
on a mixture of aquatic and terrestrial food resources, although swallows were presumably more δ15N enriched at urban reaches (based
on the lower δ15N enrichment of primary producers at urban reaches
along with the invariant swallow δ15N signatures for swallows at both
urban and rural reaches). Swallow δ13C negatively tracked periphyton
δ13C at urban, but not rural, reaches (Fig. 4), indicating that swallows
were not heavily reliant on instream primary productivity at urban
reaches. However, this relationship does not preclude swallow reliance
on aquatic emergent insects, which can be highly reliant on terrestrial
inputs of vegetation, as in this study (Fig. 3a and b). The variability
explained by this regression serves as our minimum contribution of
aquatic emergent insections to swallow diet (Fig. 4).
Bolus results from the four nest box reaches indicated that
swallows at urban locations relied more heavily on aquatic insects
than at rural locations (Fig. 5). By weight, aquatic emergent insects
comprised 84.0% of swallow diet (vs. 16.0% from terrestrial invertebrates) at urban locations, whereas aquatic emergent insects comprised
only 46.4% of swallow diet (vs. 53.6% from terrestrial invertebrates)
at rural nest box locations. Individuals of the family Coenegrionidae
(narrow-winged damselflies) made up nearly 30% of all aquatic individuals collected, whereas Aeshnidae (hawkers or darners) represented
the largest-bodied prey item.
Concentrations of Se in riparian swallows were linked with both
δ15N and δ13C isotopic values (Fig. 6a and b). Swallow Se concentrations
increased with δ15N enrichment and δ13C depletion, both of which are
isotopic signals indicative of a shift toward aquatic prey resources in
our system. Concentrations of Hg were not related to either swallow
δ15N (P = 0.971) or δ13C (P = 0.152) signatures.
Fig. 3. Dual isotope plots of δ13C and δ15N values (mean ± 1 SE) for the Scioto River
basin (Ohio, USA) aquatic and terrestrial food-web components of the Scioto River
basin (Ohio, USA) in (a) rural and (b) urban reaches.
3.2. Influence of LULC
Principal component analysis of eleven LULC measures identified
three axes with eigenvalues > 1 (Table 2). The first principal component (PC1) captured 54.4% of the variance, and was mainly driven
by factors related to urbanized landscapes (e.g., population density
[r2 = 0.91, + correlation], % impervious surface coverage [r2 = 0.86,
+], % agriculture [r2 = 0.86, −]). Accordingly, this PC was named
‘Urbanization Axis’. The second principal component explained about
19% of the variation that remained and was driven almost solely by %
tall tree cover (r2 = 0.86, +), and is hereafter referred to as “Mature
Trees Axis”. No other metric within this axis had a correlation > 0.32.
The third PC, “Shoreline Habitat Axis”, explained about 14% of the
Table 1
δ13C and δ15N values for riparian swallows, periphyton, riparian vegetation, aquatic emergent insects, and terrestrial invertebrates for all study reaches in the Scioto River basin,
Ohio, USA. Reaches are presented upstream to downstream. Note: CLSW = cliff swallow, BANS = bank swallow, TRSW = tree swallow, NRWS = northern rough-winged swallow.
Study reach
1 (Highbanks)
2 (Wetlands)
3 (Fawcett)
4 (Horseshoe)
5 (Berliner)
6 (Mosquito)
7 (Darby)
8 (Gravel Bar)
9 (Piketon)
10 (Treefall)
11 (Last)
Land use
Rural
Urban
Urban
Urban
Urban
Rural
Rural
Rural
Rural
Rural
Rural
Swallows
Periphyton
Terrestrial
Vegetation
Aquatic
Emergent Insects
Terrestrial Insect
Species
δ13C
δ15N
δ13C
δ15N
δ13C
δ15N
δ13C
δ15N
δ13C
δ15N
TRSW
TRSW
TRSW
CLSW
TRSW
NRWS
TRSW
BANS
CLSW
BANS
BANS
−24.69
−24.55
−24.42
−23.24
−26.14
−24.61
−24.07
−24.42
−23.35
−25.15
−23.98
10.016
11.00
10.80
9.04
12.39
11.77
10.00
10.37
7.91
10.87
11.13
N/A
−18.49
N/A
−17.69
−11.26
−18.79
N/A
−11.68
−21.46
−14.82
−13.41
N/A
6.88
N/A
6.14
7.35
8.42
N/A
12.39
8.66
8.78
9.64
N/A
−30.96
N/A
−30.37
−30.81
−28.06
N/A
−28.00
−31.52
−31.53
−30.56
N/A
2.72
N/A
3.08
4.22
4.68
N/A
3.74
4.53
3.27
2.89
N/A
−27.41
N/A
−27.51
−28.27
−27.04
N/A
27.78
−27.97
−27.99
−27.34
N/A
11.29
N/A
10.04
11.09
11.55
N/A
11.15
12.31
12.66
11.83
N/A
−27.78
N/A
−27.97
−27.99
−27.34
N/A
−25.09
−25.09
−23.83
−25.42
N/A
11.15
N/A
12.31
12.66
11.83
N/A
6.79
7.74
8.31
7.64
Note: N/A = not applicable because diet composition was analyzed directly using bolus samples.
J.M. Alberts et al. / Science of the Total Environment 463-464 (2013) 42–50
47
Fig. 4. Relationship between riparian swallow and periphyton δ13C at urban (solid line,
closed circles) and rural (dotted line, open circles) reaches. The slope for the regression
model was significant for urban reaches (y = −0.25x − 28.8, R2 = 0.54, P = 0.001),
but not significant for rural reaches (y = −0.07x − 25.4, R2 = 0.10, P = 0.132).
remaining variance and was strongly influenced by standing dead trees
(r2 = 0.69, +) and % overhanging vegetation (r2 = 0.48, +).
At a coarse landscape resolution, we found that swallow Hg
concentrations were significantly higher in rural than in urban
reaches (t = − 2.96, P = 0.003, df = 24), and marginally so for
Se (t = − 1.54, P = 0.068, df = 24). A combination of urban characteristics and minimal agricultural activity in the riparian zone
appeared to be, in part, driving this association, as we found that
the Urbanization Axis was negatively related to both Hg (Fig. 7a)
and Se concentrations in swallows (Fig. 7b). Urbanization was
also negatively related to δ 15N enrichment (Fig. 8), which in our
system indicates a decreasing reliance on aquatic prey along this
axis.
We found that Hg concentrations were significantly higher
in adult swallows (mean = 183.1 ppb) than nestlings (mean =
57.0 ppb) (t = − 4.01, P = 0.016, df = 4). Se concentrations in
adults (mean = 9635 ppb) were also significantly higher than in
nestlings (mean = 1997 ppb) (t = −3.10, P = 0.036, df = 4). Agedependent trends in contaminant concentration were consistent across
both urban LULC and rural LULC.
Fig. 6. Relationship between swallow Se concentration (log10) and (a) swallow δ15N
and (b) swallow δ13C. The slopes for the regression models were significant for
both δ15N (y = 0.55x + 3.26, R2 = 0.76, P b 0.001) and δ13C (y = −0.30x − 1.90,
R2 = 0.27, P = 0.032). Dashed lines represent confidence curves at α = 0.05.
4. Discussion
Aquatic emergent insects serve as important prey items for myriad
riparian consumers (Akamatsu et al., 2004; Ballinger and Lake, 2006).
As a result of these strong, cross-system linkages, terrestrial consumers
are exposed to aquatic contaminants (Akamatsu and Toda, 2011).
Walters et al. (2008), for example, showed that aquatic emergent
insects transported PCBs from contaminated lake sediments into
adjacent riparian food webs. Our understanding of the mechanisms
Table 2
Adjacent (500 m on each side of river) land-use and land-cover (LULC) principal
component analysis: eigenvalues and the percent variance captured by the principal
components (eigenvalues > 1), along with each principal component's loadings and
the proportion of the variance (r2) each variable shared with the PCA axes. Bold type
indicates the axis used as successful predictor variable in regression models.
Urbanization
Fig. 5. Contribution by weight of aquatic emergent and terrestrial insects to swallow
boluses from nest box reaches (n = 40). Error bars represent ± 1 SE from the mean.
r2
Mature trees
Loading
r2
Shoreline
habitat
Loading
r2
Riparian landscape
composition metrics
Loading
% Agriculture
% Shrub cover(1–3 m)
% Small tree cover (3–6 m)
% Tall tree cover (>6 m)
# Standing dead trees
Riparian forest width (m)
# Bridge crossings
% Impervious surface
# Canopy layers
Human population density
(residents/km2)
% Overhanging vegetation
Eigenvalue
% variance
−0.38
0.34
0.30
−0.01
0.00
−0.30
0.30
0.38
0.32
0.39
0.86
0.69
0.54
0.00
0.00
0.54
0.54
0.86
0.61
0.91
0.08
0.02
0.31
0.64
−0.34
−0.10
−0.39
−0.23
0.36
−0.10
0.01
0.00
0.20
0.86
0.24
0.02
0.32
0.11
0.27
0.02
0.02
−0.38
0.05
0.17
0.67
−0.08
−0.17
−0.04
−0.09
0.13
0.00
0.22
0.00
0.04
0.69
0.01
0.04
0.00
0.01
0.03
0.26
0.40
5.98
54.4
0.13
0.04
2.10
19.1
0.56
0.48
1.53
14.0
48
J.M. Alberts et al. / Science of the Total Environment 463-464 (2013) 42–50
Fig. 7. Relationship between Urbanization Axis (i.e., PC1) and (a) Hg concentration
(log10) and (b) Se concentration (log10) in riparian swallows. Degree of urbanization
increases from −3 to 4. The slope for the regression model was significant for
both Hg (y = −0.11x − 4.91, R2 = 0.38, P = 0.004) and Se (y = −0.17x + 9.12,
R2 = 0.63, P b 0.001). Dashed lines represent confidence curves at α = 0.05.
that influence and mediate these exposure pathways remains limited,
but given the geographic range and abundance of aquatic emergent
insects and riparian consumers, and the global proliferation of aquatic
contamination (Menzie, 1980; Walters et al., 2008), an improved
grasp of these cross-boundary flows will be crucial for managing terrestrial exposure risk from aquatic contaminants.
Many investigators have illustrated the variability in swallow
dietary composition. For example, Blancher and McNicol (1991) determined that the diet of non-egg-laying adult tree swallows was
Fig. 8. Relationship between Urbanization Axis (i.e., PC1) and δ15N in riparian swallows.
Urbanization increases from −3.0 to 4.0. The slope for the regression model was significant
(y = −0.23x + 10.9, R2 = 0.38, P = 0.031). Dashed lines represent confidence curves at
α = 0.05.
comprised of 54.9% aquatic insects by biomass. Similarly, Wayland et
al. (1998) found that insects of aquatic origin made up 50–60% of tree
swallow diet in western Canada. Collectively, our results indicate that
swallows rely on a mixture of terrestrial and aquatic insects, but that reliance on aquatic insects is likely greater at urban vs. rural reaches
(Figs. 3a & b, 5) despite no significant differences in either aquatic
(t = −0.121, P = 0.477) or terrestrial (t = 0.756, P = 0.908) aerial
insect productivity between rural and urban reaches in the study system (Kautza and Sullivan, unpublished data). Many riparian swallow
species tend to forage in open areas (Brown et al., 2002; Ghilain and
Belisle, 2008), indicating a likely preference for feeding over open
water vs. heavily-canopied riparian areas. Our urban reaches were generally characterized by densely-forested strips in the immediate riparian zone, which may encourage foraging directly over water and explain
why urban swallows fed more heavily on aquatic emergent insections
than their rural counterparts.
Contrary to our hypothesis, sediment contaminant concentrations
did not differ between land-use classes. However, Hg and Se concentrations in riparian swallows were lower at urban than at rural reaches,
despite the fact that urban nest box swallows depredated aquatic
insects more commonly than terrestrial insects, and that bolus samples
indicated that aquatic insects exhibited higher concentrations of Se
and Hg than terrestrial insects. Concomitantly, concentrations of Hg
in swallow blood and river sediment were positively related (Fig. 2),
which suggests that riparian swallows are exposed to aquatic contaminants via aquatic prey. Likewise, Se concentrations in swallows were
positively linked to both δ15N enrichment and δ13C depletion (Fig. 6),
indicators of reliance on aquatic emergent insects in our study system.
Swallows also fed at a higher trophic level at our urban reaches
(Fig. 3), indicating a potential increase in aquatic prey utilization. One
possible explanation for this apparent discrepancy between contribution of aquatic prey and contamination patterns is that swallows were
actively selecting prey that were not dominant within the system,
and therefore not included in our isotopic or contaminant analyses.
Blancher and McNicol (1991) for example, found that mayflies were
an important component of tree swallow diet despite composing a
small fraction of the insects caught in emergent traps. It is also possible
that varying bioavailability among metal speciations found within the
sediments leads to unequal transfer to swallows through predation of
aquatic emergent insects. For instance, Langner et al. (2012) observed
that blood Hg concentrations in osprey (Pandion haliaetus) chicks did
not reflect Hg concentrations in river sediments, which they suggested
was due to decreased methylation, and thus bioavailability, of Hg.
Influences of land cover on avian foraging behavior likely explain
patterns we observed in swallow contaminant concentrations.
Ormerod and Tyler (1991) found that gray wagtails (Motacilla
cinerea) changed their diet to incorporate more lepidopteran larvae
in broadleaved forests than in moorlands or coniferous forests in
Wales. Such dietary shifts in response to land-cover variation could
have a profound effect on contaminant uptake. Biomagnification of
aquatic contaminants in tree swallows, for instance, is known to be
linked with their reliance on aquatic emergent insects (Brasso and
Cristol, 2008; Hallinger et al., 2011; Jackson et al., 2011), which is
consistent with our results for Se (Fig. 6).
Our observation that contaminant biomagnification was higher in
adult swallows relative to nestlings aligns with other studies and was
consistent with our hypothesis. Evers et al. (2005), for instance, found
that methylmercury (MeHg) concentrations were five to ten times
higher in adult tree swallows than in nestlings. Although we did not
observe a difference in the dietary composition (aquatic vs. terrestrial)
between adult and immature swallows, nestlings might efficiently
remove contaminants from their blood as they are deposited into
growing feathers (Tsipoura et al., 2008). Alternatively, adult swallows
may feed on larger-bodied aquatic insects, thereby increasing exposure
to aquatic contaminants. In Canada, Quinney and Ankney (1985) found
that Insects 1–6 mm in length made up between 80% and 86% of
J.M. Alberts et al. / Science of the Total Environment 463-464 (2013) 42–50
nestling tree swallow diet, whereas McCarty and Winkler (1999)
found that adult tree swallows actively selected for larger-bodied
prey (e.g., Odonata) as compared to smaller-bodied insections (0–3 mm
in length). Temporal disparities between aquatic insect emergence and
terrestrial insect production might also influence prey availability and
feeding behavior (Nakano and Murakami, 2001; Jonsson et al., 2012),
which could result in nestlings ingesting a higher proportion of terrestrial
insects, thereby decreasing their exposure to aquatic contaminants.
5. Conclusions
Overall, our results (1) support recent research examining the
cross-system trophic linkages between aquatic emergent insects and
riparian consumers (Ballinger and Lake, 2006; Fukui et al., 2006;
Daley et al., 2011), and (2) implicate landscape alterations as a key
factor in regulating aquatic-to-terrestrial contaminant transfers. Our results also provide insight into the role of riparian swallows as integrators
of linked aquatic–terrestrial systems within the context of contaminant
exposure risk. Specifically, the reliance of swallows upon a mixture of
terrestrial and aquatic invertebrate prey makes them highly vulnerable
to contaminants from both terrestrial and aquatic systems. Their ability
to range freely and utilize multiple prey sources makes them excellent
organisms for examination of large, cross-boundary trophic and material
transfer processes.
Building upon previous work (Blancher and McNicol, 1991;
Bishop et al., 1999; Harris and Elliott, 2000; McCarty, 2002), we
suggest that riparian swallow species may capture system-level
mechanisms of contamination. We argue that the trophic dynamics
of riparian swallows and other insectivorous riparian birds represent
a critical pathway for cross-boundary contaminant transfer and
movement across linked aquatic–terrestrial landscapes. Studies across
broader geographical ranges and with increased sample sizes will be
helpful in further testing this hypothesis as will additional research relative to species-specific swallow feeding behavior, movement patterns,
and resource utilization. The potential role of swallows themselves as
vectors of contaminants (e.g., via feces, feathers, and nesting colonies)
across both local (i.e., nesting) and broader (i.e., migratory) spatial scales
will also be important in further understanding aquatic–terrestrial
contaminant fluxes. Lastly, continued research related to the mediating
role of terrestrial landscapes on transfers of contaminants from aquatic
to terrestrial systems will be critical for improved remediation and
long-term conservation of river ecosystems.
Supplementary data to this article can be found online at http://
dx.doi.org/10.1016/j.scitotenv.2013.05.065.
Acknowledgments
Research support was provided by state and federal funds appropriated to The Ohio State University, Ohio Agricultural Research and
Development Center. We would like to thank Paradzayi Tagwireyi
for his help in the field.
References
Akamatsu F, Toda H. Aquatic subsidies transport anthropogenic nitrogen to riparian
spiders. Environ Pollut 2011;159:1390–7.
Akamatsu F, Toda H, Okino T. Food source of riparian spiders analyzed by using stable
isotope ratios. Ecol Res 2004;19:655–62.
Ardia DR. Tree swallows trade off immune function and reproductive effort differently
across their range. Ecology 2005;86:2040–6.
Aubin A, Bourassa JP, Pellissi M. Effective emergence trap for capture of mosquitoes.
Mosq News 1973;33:251–2.
Ballinger A, Lake PS. Energy and nutrient fluxes from rivers and streams into terrestrial
food webs. Mar Freshw Res 2006;57:15–28.
Bartel RA, Haddad NM, Wright JP. Ecosystem engineers maintain a rare species of
butterfly and increase plant diversity. Oikos 2010;119:883–90.
Baxter CV, Fausch KD, Saunders WC. Tangled webs: reciprocal flows of invertebrate
prey link streams and riparian zones. Freshw Biol 2005;50:201–20.
49
Bigler WJ, Jenkins JH, Cumbie PM, Hoff GL, Prather EC. Wildlife and environmental health
— raccoons as indicators of zoonoses and pollutants in southeastern United States.
J Am Vet Med Assoc 1975;167:592–7.
Bishop CA, Mahony NA, Trudeau S, Pettit KE. Reproductive success and biochemical
effects in tree swallows (Tachycineta bicolor) exposed to chlorinated hydrocarbon
contaminants in wetlands of the Great Lakes and St. Lawrence River basin, USA and
Canada. Environ Toxicol Chem 1999;18:263–71.
Blancher PJ, McNicol DK. Tree swallow diet in relation to wetland acidity. Can J Zool
(Rev Can Zool) 1991;69:2629–37.
Blocksom KA, Johnson BR. Development of a regional macroinvertebrate index for large
river bioassessment. Ecol Indic 2009;9:313–28.
Blocksom KA, Walters DM, Jicha TM, Lazorchak JM, Angradi TR, Bolgrien DW. Persistent
organic pollutants in fish tissue in the mid-continental great rivers of the United
States. Sci Total Environ 2010;408:1180–9.
Brasso RL, Cristol DA. Effects of mercury exposure on the reproductive success of tree
swallows (Tachycineta bicolor). Ecotoxicology 2008;17:133–41.
Brown CR, Brown MB. Cliff swallow (Hirundo pyrrhonota). In: Poole A, Gill F, editors. The
birds of North America, 149. Washington, D.C.: The Academy of Natural Sciences,
Philadelphia, PA, and The American Ornithologists' Union; 1995
Brown CR, Sas CM, Brown MB. Colony choice in cliff swallows: effects of heterogeneity
in foraging habitat. Auk 2002;119:446–60.
Cliff A, Ord K. Spatial autocorrelation. London: Pion; 1973.
Cristol DA, Brasso RL, Condon AM, Fovargue RE, Friedman SL, Hallinger KK, et al.
The movement of aquatic mercury through terrestrial food webs. Science 2008;320:
335-335.
Custer CM. Swallows as a sentinel species for contaminant exposure and effect studies.
Wildlife Ecotoxicology: Forensic Approaches 2011;3:45–91.
Daley JM, Corkum LD, Drouillard KG. Aquatic to terrestrial transfer of sediment associated
persistent organic pollutants is enhanced by bioamplification processes. Environ
Toxicol Chem 2011;30:2167–74.
DeJong MJ. Northern rough-winged swallow (Stelgidopteryx serripennis). In: Poole A,
Gill F, editors. The birds of North America, 234. Washington, D.C.: The Academy
of Natural Sciences, Philadelphia, and The American Ornithologists' Union; 1996
Deniro MJ, Epstein S. Influence of diet on distribution of carbon isotopes in animals.
Geochim Cosmochim Acta 1978;42:495–506.
Deniro MJ, Epstein S. Influence of diet on the distribution of nitrogen isotopes in
animals. Geochim Cosmochim Acta 1981;45:341–51.
Dods PL, Birmingham EM, Williams TD, Ikonomou MG, Bennie DT, Elliott JE. Reproductive
success and contaminants in tree swallows (Tachycineta bicolor) breeding at a wastewater treatment plant. Environ Toxicol Chem 2005;24:3106–12.
Dreyer J, Hoekman D, Gratton C. Lake-derived midges increase abundance of shoreline
terrestrial arthropods via multiple trophic pathways. Oikos 2012;121:252–8.
Dunn PO, Hannon SJ. Effects of food abundance and male parental care on reproductive
success and monogamy in tree swallows. Auk 1992;109:488–99.
Echols KR, Tillitt DE, Nichols JW, Secord AL, McCarty JP. Accumulation of PCB congeners
in nestling tree swallows (Tachycineta bicolor) on the Hudson River, New York.
Environ Sci Technol 2004;38:6240–6.
Evers DC, Burgess NM, Champoux L, Hoskins B, Major A, Goodale WM, et al. Patterns
and interpretation of mercury exposure in freshwater avian communities in northeastern North America. Ecotoxicology 2005;14:193–221.
Fitzgerald WF, Engstrom DR, Mason RP, Nater EA. The case for atmospheric mercury
contamination in remote areas. Environ Sci Technol 1998;32:1–7.
Fukui D, Murakami M, Nakano S, Aoi T. Effect of emergent aquatic insects on bat foraging
in a riparian forest. J Anim Ecol 2006;75:1252–8.
Garrison BA. Bank swallow (Riparia riparia). In: Poole A, Gill F, editors. The birds of
North America, 414. Philadelphia, PA: The Birds of North America, Inc.; 1999.
Ghilain A, Belisle M. Breeding success of tree swallows along a gradient of agricultural
intensification. Ecol Appl 2008;18:1140–54.
Golondrinas de las Americas. Nest box design. http://golondrinas.cornell.edu/
Data_and_Protocol/NestBoxDesignCentimeters.html, 2011 Accessed March 24,
2010.
Hagen EM, Sabo JL. Influence of river drying and insect availability on bat activity along
the San Pedro River, Arizona (USA). J Arid Environ 2012;84:1–8.
Hallinger KK, Cornell KL, Brasso RL, Cristol DA. Mercury exposure and survival in
free-living tree swallows (Tachycineta bicolor). Ecotoxicology 2011;20:39–46.
Hamas MJ. Belted kingfisher (Cerlye alcyon). In: Poole A, Gill F, editors. The birds of
North America, 84. Washington, DC, USA: The American Ornithologists' Union;
1994.
Hammerschmidt CR, Fitzgerald WF. Methylmercury in mosquitoes related to atmospheric mercury deposition and contamination. Environ Sci Technol 2005;39:
3034–9.
Harris ML, Elliott JE. Reproductive success and chlorinated hydrocarbon contamination
in tree swallows (Tachycineta bicolor) nesting along rivers receiving pulp and
paper mill effluent discharges. Environ Pollut 2000;110:307–20.
Henschel JR, Mahsberg D, Stumpf H. Allochthonous aquatic insects increase predation
and decrease herbivory in river shore food webs. Oikos 2001;93:429–38.
Iwata T, Nakano S, Murakami M. Stream meanders increase insectivorous bird abundance in riparian deciduous forests. Ecography 2003;26:325–37.
Jackson AK, Evers DC, Folsom SB, Condon AM, Diener J, Goodrick LF, et al. Mercury
exposure in terrestrial birds far downstream of an historical point source. Environ Pollut
2011;159:3302–8.
Jardine TD, Kidd KA, Rasmussen JB. Aquatic and terrestrial organic matter in the diet of
stream consumers: implications for mercury bioaccumulation. Ecol Appl 2012;22:
843–55.
Johnson NF, Triplehorn CA. Borror and Delong's introduction to the study of insects.
Thomson Brooks/Cole; 2004.
50
J.M. Alberts et al. / Science of the Total Environment 463-464 (2013) 42–50
Jonsson M, Strasevicius D, Malmqvist B. Influences of river regulation and environmental
variables on upland bird assemblages in northern Sweden. Ecol Res 2012;27:945–54.
Keast A. The annual cycle and activity on the breeding grounds in a Canadian
broad-leafed deciduous forest bird community relationship to the prey resource
base. V+410pIn: Keast a, editor. Biogeography and ecology of forest bird communities. The Hague, Netherlands: Spb Academic Publishing Bv; 1990. p. 197–214.
Illus Maps.
Langner HW, Greene E, Domenech R, Staats MF. Mercury and other mining-related contaminants in ospreys along the Upper Clark Fork River, Montana, USA. Arch Environ
Contam Toxicol 2012;62:681–95.
Lehner A, Rumbeiha W, Shlosberg A, Stuart K, Johnson M, Domenech R, et al. Diagnostic
analysis of veterinary dried blood spots for toxic heavy metals exposure. J Anal
Toxicol 2013 In press.
Lenat DR, Crawford JK. Effects of land-use on water quality and aquatic biota of 3 North
Carolina piedmont streams. Hydrobiologia 1994;294:185–99.
Lord CG, Gaines KF, Boring CS, Brisbin LL, Gochfeld M, Burger J. Raccoon (Procyon lotor)
as a bioindicator of mercury contamination at the US Department of Energy's
Savannah River Site. Arch Environ Contam Toxicol 2002;43:356–63.
Luther D, Hilty J, Weiss J, Cornwall C, Wipf M, Ballard G. Assessing the impact of local
habitat variables and landscape context on riparian birds in agricultural, urbanized,
and native landscapes. Biodivers Conserv 2008;17:1923–35.
Lutz MA. Procedures for collecting and processing streambed sediment and pore water
for analysis of mercury as part of the National Water-Quality Assessment Program.
Open File Report 2008;2008-1279.
Malinga M, Szefer P, Gabrielsen GW. Age, sex and spatial dependent variations in heavy
metals levels in the Glaucous Gulls (Larus hyperboreus) from the Bjornoya and Jan
Mayen, Arctic. Environ Monit Assess 2010;169:407–16.
Mattson DJ, Reinhart DP. Influences of cutthroat trout (Oncorhynchus clarki) on behaviour
and reproduction of Yellowstone grizzly bears (Ursus arctos), 1975–1989. Can J Zool
(Rev Can Zool) 1995;73:2072–9.
Maul JD, Belden JB, Schwab BA, Whiles MR, Spears B, Farris JL, et al. Bioaccumulation
and trophic transfer of polychlorinated biphenyls by aquatic and terrestrial insects
to tree swallows (Tachycineta bicolor). Environ Toxicol Chem 2006;25:1017–25.
McCarty. Use of tree swallows in studies of environmental stress. Rev Toxicol (Amsterdam)
2002;4:61–104.
McCarty JP, Winkler DW. Foraging ecology and diet selectivity of tree swallows feeding
nestlings. Condor 1999;101:246–54.
Menzie CA. Potential significance of insects in the removal of contaminants from
aquatic systems. Water Air Soil Pollut 1980;13:473–9.
Meyer LA, Sullivan SMP. Bright lights, big city: influences of ecological light pollution
on reciprocal stream–riparian invertebrate fluxes. Ecol Appl 2013 In press.
Morrissey CA, Bendell-Young LI, Elliott JE. Linking contaminant profiles to the diet
and breeding location of American dippers using stable isotopes. J Appl Ecol 2004;41:
502–12.
Nakano S, Murakami M. Reciprocal subsidies: dynamic interdependence between
terrestrial and aquatic food webs. Proc Natl Acad Sci U S A 2001;98:166–70.
Oneal AS, Rotenberry JT. Scale-dependent habitat relations of birds in riparian corridors
in an urbanizing landscape. Landsc Urban Plann 2009;92:264–75.
Ormerod SJ, Tyler SJ. The influence of stream acidification and riparian land-use on the
feeding ecology of gray wagtails (Motacilla cinerea) in Wales. Ibis 1991;133:53–61.
Paetzold A, Schubert CJ, Tockner K. Aquatic terrestrial linkages along a braided-river:
riparian arthropods feeding on aquatic insects. Ecosystems 2005;8:748–59.
Pennington DN, Blair RB. Habitat selection of breeding riparian birds in an urban
environment: untangling the relative importance of biophysical elements and spatial
scale. Divers Distrib 2011;17:506–18.
Pennington DN, Hansel J, Blair RB. The conservation value of urban riparian areas
for landbirds during spring migration: land cover, scale, and vegetation effects.
Biol Conserv 2008;141:1235–48.
Polis GA, Anderson WB, Holt RD. Toward an integration of landscape and food web
ecology: the dynamics of spatially subsidized food webs. Annu Rev Ecol Syst
1997;28:289–316.
Polis GA, Sanchez-Pinero F, Stapp PT, Anderson WB, Rose MD. Trophic flows from
water to land: marine input affects food webs of islands and coastal ecosystems
worldwide. In: Polis GA, Power ME, Huxel GR, editors. Food webs at the landscape
level. Chicago, Illinois: University of Chicago Press; 2004. p. 200–16.
Quinney TE, Ankney CD. Prey size selection by tree swallows. Auk 1985;102:
245–50.
Reavie ED, Jicha TM, Angradi TR, Bolgrien DW, Hill BH. Algal assemblages for large river
monitoring: comparison among biovolume, absolute and relative abundance
metrics. Ecol Indic 2010;10:167–77.
Rencher AC. Methods of multivariate analysis. New York, NY, U.S.A.: Johny Wiley and
Sons, Inc.; 1995
Robertson RJ, Stutchbury BJ, Cohen RR. Tree swallow (Tachycineta bicolor). In: Poole A,
Stettenheim P, Gill F, Poole A, Stettenheim P, Gill F, editors. The birds of North America,
11. Philadelphia, PA: The Academy of Natural Sciences; 1992. (The American Ornithologists' Union, Washington, D.C.).
Roux KE, Marra PP. The presence and impact of environmental lead in passerine birds
along an urban to rural land use gradient. Arch Environ Toxicol Chem 2007;53:
261–8.
Sabo JL, Power ME. Numerical response of lizards to aquatic insects and short-term
consequences for terrestrial prey. Ecology 2002;83:3023–36.
Shlosberg A, Rumbeiha WK, Lublin A, Kannan K. A database of avian blood spot
examinations for exposure of wild birds to environmental toxicants: the DABSE
biomonitoring project. J Environ Monit 2011;13:1547–58.
Stepenuck KF, Crunkilton RL, Wang LZ. Impact of urban land use on macroinvertebrate
communities in southeastern Wisconsin streams. J Am Water Resour Assoc
2002;38:1041–51.
Sullivan SMP, Rodewald AD. In a state of flux: the energetic pathways that move
contaminants from aquatic to terrestrial environments. Environ Toxicol Chem
2012;31:1175–83.
Sullivan SMP, Vierling KT. Exploring the influences of multiscale environmental factors
on the American dipper Cinclus mexicanus. Ecography 2012;35:624–36.
Sullivan SMP, Watzin MC. Relating stream physical habitat condition and concordance of biotic productivity across multiple taxa. Can J Fish Aquat Sci
2008;65:2667–77.
Sullivan SMP, Watzin MC, Keeton WS. A riverscape perspective on habitat associations
among riverine bird assemblages in the Lake Champlain Basin, USA. Landsc Ecol
2007;22:1169–86.
Sweeney BW, Vannote RL. Population synchrony in mayflies — a predator satiation
hypothesis. Evolution 1982;36:810–21.
Townes H. A light weight malaise trap. Entomol News 1972;83:239–47.
Tsipoura N, Burger J, Feltes R, Yacabucci J, Mizrahi D, Jeitner C, et al. Metal concentrations
in three species of passerine birds breeding in the Hackensack Meadowlands of
New Jersey. Environ Res 2008;107:218–28.
United States Census Bureau. State and county quick facts. http://quickfacts.census.
gov/qfd/states/39/39049.html, 2010 (Accessed July 29, 2012).
Vander Zanden MJ, Sanzone DM. Food web subsidies at the land–water ecotone. In:
Polis GA, Power ME, Huxel GR, editors. Food webs at the landscape level. Chicago,
USA: University of Chicago Press; 2004.
Varley JD, Schullery P. Yellowstone Lake and its cutthroat trout. Science and ecosystem
management in the national parks 1996;49–73.
Vessel RD. Energetics of the belted kingfisher (Megaceryle alcyon). Corvalis, OR, USA:
Oregon State University; 1978.
Wada H, Cristol DA, McNabb FMA, Hopkins WA. Suppressed adrenocortical responses
and thyroid hormone levels in birds near a mercury-contaminated river. Environ
Sci Technol 2009;43:6031–8.
Walters DM, Fritz KM, Otter RR. The dark side of subsidies: adult stream insects export
organic contaminants to riparian predators. Ecol Appl 2008;18:1835–41.
Walters DM, Mills MA, Fritz KM, Raikow DF. Spider-mediated flux of PCBs from contaminated sediments to terrestrial ecosystems and potential risks to arachnivorous
birds. Environ Sci Technol 2010;44:2849–56.
Wayland M, Trudeau S, Marchant T, Parker D, Hobson KA. The effect of pulp and paper
mill effluent on an insectivorous bird, the tree swallow. Ecotoxicology 1998;7:
237–51.
White D, Johnston K, Miller M. Ohio river basin. Rivers of North America. Burlington,
Massachusetts: Elsevier Academic Press; 2005. p. 375–424.
Wright SP. Adjusted p-values for simultaneous inference. Biometrics 1992;45:1005–13.
Zar JH. Biostatistical analysis. Englewood Cliffs, New Jersey: Prentice-Hall; 1984.
Zelt RB, Johnson MR. Protocols for mapping and characterizing land use/land cover
in riparian zones. U.S. geological survey open-file report; 2005. [2005-1302].