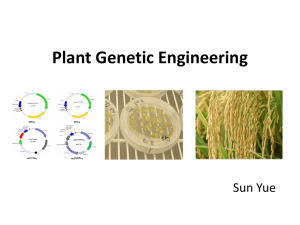
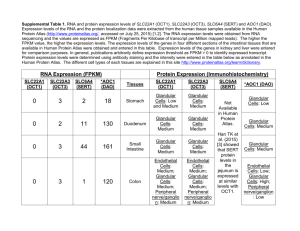
THAISZIA Systematic significance of trichomes and foliar epidermal
advertisement

Thaiszia - J. Bot., Košice, 22 (1): 1-31, 2012 THAISZIA http://www.bz.upjs.sk/thaiszia JOURNAL OF BOTANY Systematic significance of trichomes and foliar epidermal morphology in the species of Stachytarpheta Vahl. (Verbenaceae) from Nigeria OLUBUKOLA ADEDEJI Department of Botany, Obafemi Awolowo University, Ile-Ife, Osun State, Nigeria; oadedeji@oauife.edu.ng Adedeji O. (2012): Systematic significance of trichomes and foliar epidermal morphology in the species of Stachytarpheta Vahl. (Verbenaceae) from Nigeria. – Thaiszia – J. Bot. 22 (1): 1-31. – ISSN 1210-0420. Abstract: The foliar epidermal surfaces, leaf venation and trichome studies of different plant parts of the three species of Stachytarpheta Vahl. family Verbenaceae found in Nigeria were studied and compared. Observations were carried out by light microscope. Across the genus, leaves were amphistomatic and amphitrichomic. Qualitative foliar anatomical characters found to be most useful in species delimitation were stomata shape, trichome types, anticlinal cell wall pattern on adaxial surface and veinlet termination number. Quantitative characters of taxonomic importance, as revealed by analysis of variance (ANOVA) and duncan multiple range test (DMRT) were length and width of epidermal cells, stomatal index and stomatal size. Two trichome types, glandular and non-glandular were observed in the genus. The non-glandular trichomes were of higher numerical density on adaxial surface than on abaxial surface. The implication of this in terms of protection against excessive radiation, chemical control strategies and higher temperatures was mentioned. On the abaxial surface however, the glandular secreting trichomes were more numerically than the non-glandular. The importance of the preponderance of these trichomes on the abaxial surface in relation to the role they play in the protection of essential oils earlier reported in the family and genus, was discussed. Stomatal abnormalities observed in the genus were, contiguous stomata arrangement in S. cayennensis and aborted guard cells in S. indica. Leaf venation type in the genus is pinnate craspedodromous simple 1 with recurved secondary veins. Trichome distribution survey in the different plant parts revealed specific combination types that would be useful in species delimitation, especially in fragmentary parts identification. Keywords: Stachytarpheta, Verbenaceae, adaxial, abaxial, areole, veinlet, stomata, amphitrichomic, Nigeria. Introduction Verbenaceae is predominantly a tropical family exhibiting a wide range of growth habit and inhabiting diverse habitats (LILLYAMMA & SHAH 1987). The family has about 98 genera and 3,000 species (IDU et al. 2009). They are low shrubs, herbs or trees. Flowers are in spikes. The genus Stachytarpheta Vahl belongs to the family Verbenaceae and is represented in West Africa and Nigeria by three species namely Stachytarpheta cayennensis (Rich.) Vahl., S. angustifolia (Mill.) Vahl and S. indica (L.) Vahl (HUTCHINSON & DALZIEL 1963). They are economic plants and may be grown as ornamentals (GILL 1988). Verbenaceae family members are popular in traditional medicine (IYANG 2003). All the Stachytarpheta species have been used ethnomedically as anti-diabetic, arbotifacient, emmenagogue, sedative, anti-hypertensive, anti-asthmatic and anti-fever (SCHWONTKOWSKI 1993; BALIK 1985; DUKE & VAZQUEZ 1994). They are also used for dysentery, as vermifuge, anti-gonorrhoea, and to cure cataract, sore in children’s ear and also heart trouble. Trichomes were among the first anatomical features of plants to be recognized by early microscopists and they have played a key role in plant taxonomy (BEHNKE 1984). Simple or non-glandular trichomes serve the plant and humans in many ways. The morphological and mechanical features (density, size, shape, surface texture, hair orientation) of trichomes can influence many aspects of plant physiology and ecology (W AGNER et al. 2003). Trichomes may serve to protect buds of some plants until defence phytochemicals are produced (JOHNSON 1975). Thus although simple or non-glandular trichomes are not known to accumulate post-deterrent phytochemicals, their importance to plants and humans should not be under estimated (W AGNER et al. 2003). The importance of non-glandular trichomes to humans is well illustrated by cotton. The non-glandular trichomes of cotton ovules (cotton fibres, which aid seed dispersal) are exploited more by humans than any single medicinal phytochemical or spice (W AGNER et al. 2003). Glandular secretory trichomes, like simple trichomes, can influence plant functions by virtue of their physical properties (size, density). However, in addition, glandular secretory trichomes can affect host disease and pest resistance based on phytochemicals they secrete. The accumulation of toxic compounds at the surface allows their direct contact with insects, pathogens and herbivores. Thus, trichome exudates are ideally positioned to provide a first line of defence against attacking organisms, perhaps providing time for activation of induced defences, (W AGNER et al. 2003). The trichome types are not only useful in the identification of species, but also 2 their corresponding parts, thus being important in pharmacognosy, archaeobotany, paleobotany and agronomy (RAO & RAMAYYA 1977). The foliar epidermis is one of the most significant taxonomic characters from the biosystematic point of view and the taxonomic studies of a number of families are made on the basis of leaf epidermis (BHATIA 1984; JONES 1986; ADEDEJI 2004; ADEDEJI & JEWOOLA 2008). Although the epidermal anatomy has been described in the leaves of a number of Verbenaceae species (PASSOS et al. 2009; METCALFE & CHALK 1950; INAMDAR 1969), surprisingly, Stachytarpheta is a genus with paucity of anatomical research information. IDU et al. (2009), in their work on morphological and anatomical studies of the leaf and stem of S. jamaicensis (L. C. Rich) Schau, mentioned the presence of trichomes and stomata on the leaf surfaces of the two species, but did not identify the types nor give any description. One of the aims of this study is to fill this knowledge gap. Other aims are to undertake an in-depth study of the foliar epidermis and distribution of trichomes on plant parts in the three species of Stachytarpheta in West Africa and Nigeria as reported by HUTCHINSON & DALZIEL (1963). Material and methods Leaf parts, stem, flowers were collected fresh from the three species of Stachytarpheta, S. cayennensis (Rich.) Vahl., S.indica (Linn.) Vahl. and S. angustifolia (Mill.) Vahl. from various locations on the campus of Obafemi Awolowo University, Ile-Ife, Osun State, Nigeria. For the purpose of comparative foliar anatomical work, small sizeable portions of the leaves were cut from the median parts of mature and well expanded leaves, that is, midway between the base and the apex, for each species studied. The portions of the leaf blade were boiled for about 25 minutes in 90% alcohol in the oven at about 60ºC in order to remove the chlorophyll. The leaf portions were then washed in four to five changes of water, after which they were then boiled in 5 % sodium hydroxide solution for about 40 minutes until the materials were decolourized. As soon as the materials became decolourized, they were washed thoroughly in order to remove the alkaline solution. The partly cleared portions of the leaf were finally cleared in 5 % solution of domestic bleach (parozone) for about 30 minutes. The portions of the leaf were finally washed properly in three to four changes of water and stored in 50 % ethanol for anatomical studies. The cleared portions of the leaf were stained in 1 % aqueous solution of safranin O for 3-5 minutes, washed in 3-4 changes of water to remove excess stain and mounted in 10% glycerol solution for venation studies. For the preparation of epidermal peels, the scrape technique of METCALFE (1960) was used whereby the required epidermis was obtained by scraping it from the unrequired mesophyll. Epidermal peels of both adaxial and abaxial surfaces were made by placing the desired epidermal surfaces face down and scraping off with a sharp blade all tissues above the desired epidermis until the required epidermis was reached. The leaf material being scraped was intermittently irrigated with water. The desired epidermal surface was placed on a clean glass slide with the surface desired placed up. The epidermal peels were 3 stained in 1% aqueous safranin O for 5-10 minutes and rinsed carefully in water to remove excess stain. The peels were washed in 3-4 changes of water and mounted in dilute glycerol solution. Stomatal size was calculated by multiplying the length and width of the stomata. Stomatal index is the percentage proportion of the number of stomata to the other epidermal cells present on a leaf portion. The Stomatal Index is expressed by the formula S x 100% S. I. = E+S Where, S. I. = stomatal index, S = number of stomata per unit area, E = number of ordinary epidermal cells plus the subsidiary cells in the same unit area. For the trichome studies of the different plant organs, epidermal peels and transverse sections of different plant parts except the petals and petal tube (which were observed directly) were made, stained with safranin O and mounted in 70% glycerol. Photomicrographs were taken using a light microscope equipped with an AmScope camera. The mean, standard error and standard deviation values for the quantitative characters were obtained using PAST Statistical Packages. The analysis of variance (ANOVA) and duncan multiple range test (DMRT) were obtained using SASS Package. Cluster diagrams (dendrogram) of the foliar epidermal characters and trichomes on the adaxial and abaxial epidermal surfaces were also prepared Results/Observations Epidermal surfaces S. cayennensis (Tab. 1, Fig. 1A-D) Epidermal cells shape is largely polygonal, regularly arranged on adaxial surface, 50-87.5 µm long and 10-45 µm wide; polygonal irregularly arranged on abaxial surface, 32.5-77.5 µm long and 15-37.5 µm wide. Anticlinal cell wall straight to wavy on adaxial surface, sinuous only on abaxial surface. On both surfaces, epidermal cells overlaying the veins are elongated and polygonal, occasionally rectangular with straight anticlinal walls, 70-162.5 µm long and 17.537.5 µm wide on adaxial surface, 75-140 µm long and 12.5-20 µm wide on abaxial surface. Stomata are restricted to the non-vein regions on both surfaces, largely diacytic, often anisocytic, occasionally anomocytic on both surfaces, circular to elliptic in shape on both surfaces, with more circular shape on adaxial than abaxial and more elliptic on abaxial than on adaxial. Stomata size 309.38687.5 µm² on adaxial surface, 337.5-687.5 µm² on abaxial surface. Stomatal index 14.72%-21.77% on adaxial and 26.34-36.93% on abaxial. Big rectangular, triangular, spherical shaped crystals and small crystals in aggregates present. 4 S. angustifolia (Tab. 2, Fig. 5A-F, 8D, G) Epidermal cells shape is polygonal irregularly arranged on both adaxial and abaxial epidermal surfaces, 55-112.5 µm long and 30-72.5 µm wide on adaxial surface, 57.5-112.5 µm long and 25-50 µm wide on abaxial surface. Anticlinal cell wall undulating to sinuous on adaxial surface, sinuous only on abaxial surface. Epidermal cells overlaying the veins are elongated, polygonal to rectangular in shape with straight anticlinal walls on both surfaces, 75-192.5 µm long and 17.5-32.5 µm wide on adaxial surface, 90-182.5 µm long and 17.5-30 µm wide on abaxial surface. Stomata present on non-vein regions only on both surfaces, largely diacytic, occasionally anisocytic and anomocytic on adaxial and abaxial surfaces, elliptic in shape on both surfaces. Stomata size 525-650 µm² on adaxial surface, 375-568.75 µm²on abaxial surface. Stomatal index is 24.7231.34% on adaxial, 28.87-33.64% on abaxial surface. Spherical to rectangular shaped crystals and groups of small crystals present. S. indica (Tab. 3, Figs. 9A-E, I, 11F) Epidermal cells shape polygonal, irregularly arranged on both surfaces, 65137.5 µm long and 25-60 µm wide on adaxial surface, 55-127.5 µm long and 27.5-67.5 µm wide on abaxial surface. Anticlinal cell wall undulating to sinuous on adaxial surface, sinuous only on abaxial surface. On both surfaces, epidermal cells overlaying the veins are elongated, polygonal on adaxial surface, polygonal to rectangular on abaxial surface, anticlinal wall straight on both surfaces 77.5175 µm long and 22.5-35 µm wide on adaxial surface, 75-140 µm long and 12.520 µm wide on abaxial surface. Stomata found on non-vein regions only, largely elliptic in shape on both surfaces. Stomata size 568.75-987.5 µm² on adaxial surface, 407.5-937.5 µm² on abaxial surface. Stomatal index is 10.71-19.40% on adaxial surface, 22.73-30.43% on abaxial surface. Trichome studies S. cayennensis (Tabs. 1, 7-8, Figs. 2A-G, 3A-D, 4A-G) Epidermal surfaces: Adaxial: Non-glandular: spine-like, unicellular, bicellular, multicellular Glandular: 1, 2, 4, 5-9 cells secrotory head, short stalk 2 celled secretory head. Abaxial: Non-glandular: spine-like, unicellular. bicellular, multicellular Glandular: 1, 2, 4, 5-9 cells secretory head. Midrib: Non-glandular: bicellular, multicellular Glandular: absent Petiole: Non-glandular: bicellular, multicellular Glandular: 2 cells secretory head, short stalk, 4 cells secretory head. Stem: Non-glandular: bicellular, multicellular 5 Glandular: 2 cells secretory head. Petals: Non-glandular: unicellular Glandular: short stalked 1-5 cells secretory head Petal tube: Non-glandular: absent Glandular: long stalked 1-4 cells secretory head. Calyx: Non-glandular: spine-like, multicellular Glandular: 2 cells secretory head Veins: Non-glandular: spine-like, unicellular, bicellular Glandular: 2 cells secretory heads S. angustifolia (Tabs. 2, 7-8, Figs. 5E, H-I, 6A-H, 7A-I, 8A-B, E). Epidermal surfaces: Adaxial: Non-glandular: bicellular Glandular: 1, 3, 4, 5-9, 10-16 - celled secretory head. Abaxial: Non-glandular: bicellular, multicellular Glandular: 1, 3, 4, 5-9, 10-16 cells secretory head Midrib: Non-glandular: bicellular, multicellular Glandular: 3, 4, 5-9 cells secretory head. Petiole: Non-glandular: unicellular, bicellular, multicellular Glandular: 4 cells secretory head Stem: Non-glandular: spine-like, multicellular Glandular: 2, 5-9 cells secretory head Petals: Non-glandular: unicellular Glandular: 1 cell secretory head, short stalk 1-6 cells secretory head. Petal tube: Non-glandular: absent Glandular: long stalk 1-2 cells secretory head Calyx: Non-glandular: spine-like, unicellular, bicellular Glandular: 3-4 cells secretory head. Veins: Non-glandular: spine-like, unicellular, bicellular, multicellular Glandular: 2, 4, 5-9 cells secretory head. S. indica (Tabs. 3, 7-8, Figs. 9F-G, 10A-G, 11A-E, G-I) Epidermal surfaces: 6 Adaxial: Non-glandular: spine-like, unicellular Glandular: 1, 2, 3 cells secretory head. Abaxial: Non-glandular: unicellular Glandular: 1, 2, 3 cells secretory head Midrib: Non-glandular: bicellular, multicellular Glandular: 2 cells secretory head. Petiole: Non-glandular: bicellular, multicellular Glandular: 2 cells secretory head, short stalk 1-2 cells secretory head Stem: Non-glandular: unicellular, multicellular Glandular: 1-2 cells secretory head. Petals: Non-glandular: unicellular Glandular: short stalk 1-7 cells secretory head Petal tube: Non-glandular: absent Glandular: short stalk, 1-2 cells secretory head, long stalk 1 cell secretory head. Calyx: Non-glandular: spine-like Glandular: 1-2 cells secretory head. Veins: Non-glandular: spine-like, bicellular, multicellular Glandular: 2 cells secretory head. Leaf Venation Studies S. cayennensis (Tab. 6, Fig. 9H) Venation type is pinnate craspedodromous simple, secondary veins recurved. Areole shape is polygonal, occasionally rectangular 260-450 µm long and 200430 µm wide. Veinlet termination 0-2 per areole, simple to branched. S. angustifolia (Tab. 6, Fig. 5G) Venation type is pinnate craspedodromous simple with recurved veins. Areole shape largely polygonal, occasionally triangular or rectangular, 230-700 µm long and 180-410 µm wide. Veinlet termination 0-4 per areole, largely branched. S.indica (Tab. 6, Fig. 9H) Venation type is pinnate craspedodromous simple, secondary veins recurved. Areole shape largely polygonal, occasionally rectangular, 310-830 µm long and 200-310 µm wide. Many areoles without any veinlet termination observed, 1-2 per areole when they occur, simple to branched. 7 Trichome Density on the foliar epidermal surfaces S. cayennensis (Tab. 7): Adaxial surface: Non-glandular 5-10; Glandular 0-3 Abaxial surface: Non-glandular 0-3; Glandular 1-6 S. angustifolia (Tab. 7): Adaxial surface: Non-glandular 0-1; Glandular 4-17 Abaxial surface: Non-glandular 0-1; Glandular 6-18 S. indica (Tab. 7): Adaxial surface: Non-glandular 0-1; Glandular 0-1 Abaxial surface: Non-glandular 0-1; Glandular 1-2. Statistical Analysis Tables 1-3 present the Simple Descriptive Statistics of the quantitative characters of foliar epidermal surfaces of the three Stachytarpheta species. analysis of variance (ANOVA) for epidermal cell length, epidermal cell breadth, stomatal size, stomata index, vein cell length and vein cell breadth revealed significant differences at Pr. 0.0001 for all characters on both adaxial and abaxial epidermal surfaces except for vein cell length and breadth on adaxial surface which showed no significant difference. Duncan multiple range test (DMRT) showed interesting species groupings based on these characters (Tab. 4). The dendrogram from the Cluster Analysis based on quantitative data also show species relationships (Figs. 12-13). Discussion The present investigation represents the first detailed qualitative and quantitative study of leaf epidermis, leaf venation and trichome studies in the genus Stachytarpheta in Nigeria. The three species reported in Nigeria and West Africa, S. cayennensis, S. angustifolia, S. indica, are included in the study. The foliar epidermis is one of the most significant taxonomic characters from the biosystematic point of view and the taxonomic studies of a number of families and genera have been made on the basis of leaf epidermis (BHATIA 1984; JONES 1986; ADEDEJI 2004; ADEDEJI et al 2007; ADEDEJI & JEWOOLA 2008). SCOTLAND et al. (2003) is of the opinion that rigorous and critical anatomical studies of fewer morphological characters in the context of molecular phylogenies is fruitful to integrate the strength of morphological data with those of sequence data (HAYAT et al. 2009) Leaves are amphistomatic and amphitrichomic in the three species. Epidermal cells on adaxial surface are generally polygonal in shape, regularly arranged in S. cayennensis, sinuous, irregularly arranged in the other two species. Anticlinal cell walls on adaxial surface are undulating to sinuous in S. angustifolia and S. indica but straight to wavy in S. cayennensis. This delimits S. cayennensis from the other two species. On the abaxial epidermal surface however, the epidermal cell 8 shape is polygonal, irregularly arranged with sinuous anticlinal walls in the three species. The sinuosity of the epidermis cells appears to be fixed genetically. SHAHEEN et al. (2010) delimited Althaea and Alcea, the two closely related genera in the family Malvaceae using anatomical sizes of the epidermal cells. In this study, length and width of the epidermal cells is of taxonomic significance and can be helpful in delimiting the three species of Stachytarpheta as indicated by the species grouping from Duncan Multiple Range Test (Tab. 4). It is longest in S. indica on both adaxial and abaxial surfaces while it is widest in S. angustifolia on the adaxial surface and widest in S. indica on abaxial surface. Epidermal cells overlaying the veins on both adaxial and abaxial epidermal surfaces are largely polygonal with straight anticlinal walls. From the ANOVA Test for significance and Duncan Multiple range Test, the cells are not significantly different in length and breadth on the adaxial epidermal surfaces of the three species but are significantly different on the abaxial surface, cell breadth can therefore be used for delimiting the species. According to DILCHER (1974), stomata are ecologically less variable characters. Stomata in the genus are largely diacytic, occasionally anisocytic and anomocytic. No clear cut delimitation of any taxa can be made based on stomatal type, hence the character is not of taxonomic importance in the genus. METCALFE & CHALK (1950), in their summary on family Verbenaceae reported that stomata on the adaxial surface of a few species of the genus Stachytarpheta are arranged in groups. INAMDAR (1969) reported contiguous stomata, stomata with a single guard cell, and aborted guard cells as abnormalities in the family Verbenaceae. Two out of the three mentioned abnormalities occur in this genus. They are stomata arrangement in groups (contiguous) in the upper or adaxial surface of S. cayennensis and aborted guard cells occurring in S. indica. In the three species studied, the stomatal index was higher on the abaxial surface than on the adaxial surface. This seems to be a preventive mechanism against photoinhibition, since the adaxial surface is more exposed to solar radiation as most of the leaves are in horizontal position (SMITH et al. 1998). The size and shape of stomata are important characters taxonomically (THAIR & RAJPUT 2009). Stomatal index and size, though quantitative are of taxonomic importance in the genus Stachytarpheta as observed in this study. The analysis of variance (ANOVA) Test for significance, revealed a significant difference among species in the two characters, while the duncan multiple range test revealed delimitation at species level. S. indica has the biggest stomatal size on both adaxial and abaxial epidermal surfaces with a corresponding lower stomatal index on both surfaces. Stomatal shape is largely circular to elliptic in S. cayennensis but elliptic largely in S. angustifolia and S. indica. Venation type in the genus is pinnate craspedodromous simple with recurved secondary veins. The areole shape are generally polygonal in all 3 species, occasionally rectangular or triangular. Areole length is longest in S. indica and shortest in S. cayennensis (Tab. 6), while it is widest in S. cayennensis and narrowest in S. indica. Veinlet termination number per areole ranged from 0-2 in S. cayennensis, often 0, occasionally 1-2 in S. indica, 0-4 in S. angustifolia, 9 simple to branched in the 3 species with S. angustifolia having more branched veinlet termination in the areoles than S. indica and S. cayennensis. OGUNDIPE & W UJEK (2004) reported veinlet termination number to be one of the most useful anatomical characters in the family Bignoniaceae. A study of the trichome types in the different organs show that both glandular and non-glandular trichome types can be found in the genus and their organographic distribution within each species vary. ADEDEJI et al. 2007, reported the importance of trichome distribution types in the different organs of the plant body in the delimitation of genera and species within the family Solanaceae. RAMMAYA & RAO (1976), RAO & RAMMAYA (1977), YAN-MING & RUW EN (1993), CELKA et al. (2006), ADEDEJI (2007), have all emphasized the taxonomic implication of trichomes. The glandular and non-glandular trichomes observed in the Stachytarpheta species, are distributed widely in the Verbenaceae family (SOLEREDER 1908; METCALFE & CHALK 1950). It must however be noted that trichome distribution in the genus is not abundant, they are present but often sparsely distributed. The micro morphological characteristics of foliar trichomes have played an important role in plant systematics, especially of particular groups at generic and specific levels (HARDIN 1979). Such type of studies on the field has fascinated plant morphologists and systematists towards the diversity of trichome features (YANMING & RU-W EN 1993). On the foliar epidermal surfaces, non-glandular trichome is of higher frequency on adaxial surface than on abaxial surface, occurring largely in S. cayennensis than in S.angustifolia and S. indica. The largest frequency of these trichomes in the adaxial surface of S. cayennensis can be related to protection against excessive radiation and higher temperatures, as reported in the literature (FRANCINO 2006; VALKAMA 2003; PASSOS et al. 2009). However in most cases, the true ecological function of the trichomes has not been supported by experimental data (W ERKER 2000). The penetration of herbicides in the plant tissues is essential for the effective chemical control of weed (PROCOPIO 2003). The anatomical characteristics practically determine the ease with which these products can be absorbed (HESS & FALK 1990). Trichomes in the leaf surface can intercept pulverized drops, preventing these from reaching the epidermis. Thus the high frequency of non-glandular trichomes in the adaxial surface of S. cayennensis would be advantageous for the species when undergoing chemical control strategies. Chemical control would be prone to negatively affect S. angustifolia and S. indica with low frequency of non-glandular trichomes in the adaxial surface (Tab. 7), rendering the epidermal cells more exposed to the agrochemical action. S. angustifolia is unique in being the only species without the spine-like non-glandular trichomes on both adaxial and abaxial epidermal surfaces. The different types of glandular trichomes on the foliar epidermal surfaces described for the three species of Stachytarpheta are important distinctive characteristics. The Verbenaceae family stands out for presenting different types of glandular trichomes (PASSOS et al. 2009). PASSOS et al. (2009) reported that 10 the occurrence of glandular trichomes can be considered an important distinctive characteristic for L. camara and L. radula (family Verbenaceae) they studied. Trichomes, particularly the secretory ones are considered important tools for taxonomy (SOLEREDER 1908; METCALFE & CHALK 1950; FAHN 1979; THEOBALD et al. 1979). The three species of Stachytarpheta studied can be delimited on the basis of the glandular trichome type on the epidermal surfaces. The 4 cells secretory head glandular trichome type was observed to be the major glandular type in S. angustifolia while the 2 cells type was predominant on the foliar epidermal surfaces of S. indica. S. angustifolia can also be delimited from the other two species because of the presence of 10-16 cells secretory head glandular trichome which is absent in the other two species. S. cayennensis can also be delimited from S. indica by the presence of 4, 5-9 cells secretory head glandular trichome. The glandular trichomes in the three species are more abundant in the abaxial surface than in the adaxial (Tab. 7). This tallies with the observation of PASSOS et al. (2009), in their studies on the genus Lantana. Aromatic plants grow in sunny environments and the trichomes, being protected in the abaxial surface, allow the secretions to remain for an extended time in the plant.. SOLEREDER (1908), RAGHAVAN & ARORA (1960), INAMDAR (1969), THEOBALD et al. (1979), SINHA & SHARMA (1984), GOTTLIEB & SALATINO (1987), RUEDA (1993), JUDD et al. (1999) have reported the presence of glandular trichomes secreting essential oils in the Verbenaceae. These oils normally evaporates and are released under high temperature and low humidity, hence their preponderance on the abaxial surface is largely for protection. Moreover, the sites of accumulation have an important influence on the effectiveness of anti-herbivory substance (GERSHENZON & CROTEAU 1991). S. angustifolia is the species with the highest frequency of glandular trichomes, occurring more on the abaxial surface. This is quite diagnostic for this species, suggesting its ability for secretion of these essential oils more than in the other two species. The glandular trichomes also present ecological significance, being associated with the plant interaction with the environment, interfering efficiently against the attack of herbivores and pathogens (LEVIN 1973; JOHNSON 1975; RODRIGUEZ et al. 1984; JUNIPER & SOUTHWOOD 1986; MARQUIS 1992). Evidence from wild and cultivated species gives support to this ecological role (DUFFEY 1986; JEFFREE 1986; DAVID & EASWARAMOORTHY 1988; W OODMAN & FERNANDES 1991; BERNAYS & CHAPMAN 1994; PETER et al. 1995; ROMEIS et al. 1999). A trichome distribution survey of the other plant parts, (Tab. 8) revealed specific combination types that can be used for species delimitation. The bicellular and multicellular non-glandular trichome types can be found on the midrib of the three species while S. indica can be delimited from the other two species by the presence of the 2 cells secretory head glandular trichome which is absent in the other two species. S. angustifolia is also unique in possessing the 3, 4, and 5-9 cells secretory head glandular tichome absent in the other two species. Trichome types in the petiole of S. indica and S. cayennensis are the same, bicellular and multicellular non-glandular, 2 cells secretory head and short stalked secretory head glandular trichomes, while S. angustifolia differs from the 11 others in having the unicellular non-glandular and the 4 cells secretory head glandular trichome. On the stem, trichome types multicellular non-glandular and 2 cells secretory head glandular tichomes are observed in the three species while each has specific trichome types delimiting it. Unicellular non-glandular trichome and short stalked secretory head glandular trichome are both present on the petals of the three species with S. angustifolia having an additional 1 cell secretory head glandular trichome type. The long stalked secretory head glandular trichome type is unique to the petal tube and observed in the three species. Spine-like non-glandular trichome was observed on the calyx of the three species while each has additional types diagnostic for it. The veins also revealed specific trichome combination types unique for each species. The specific organographic trichome combination types in each species would be useful in fragmentary parts identification. Crystals in the form of small needles or prisms are widely distributed in the parenchymatous tissues of both leaf axis, of the family Verbenaceae (METCALFE & CHALK 1950). In this study, crystals of different sizes and shapes were observed in the various plant parts in the three species of Stachytarpheta, occurring singly when big, and in groups when small. Cluster diagrams (Figs. 12-13) show the relationship or closeness of the three species to one another based on the trichomes and epidermal surfaces quantitative characters. Based on the adaxial surface characters, S. angustifolia and S. indica are closer while S. cayennensis separates out. Dendrogram of the abaxial characters reveals a closer similarity between S. cayennensis and S. angustifolia. This confirms the report of ADEDEJI (2010) who worked on the palynology of the genus, that S. cayennensis and S. angustifolia are the closest with S. indica closer to S. angustifolia and farther from S. cayennensis References ADEDEJI O. (2004): Leaf epidermal studies of the species of Emilia Cass. (Senecioneae, Asteraceae) in Nigeria. - Botanica Lithuanica, 10(2): 121-133. ADEDEJI O, AJUWON O.Y. & BABAWALE O. (2007): Foliar epidermal studies, organographic distribution and taxonomic importance of trichomes in the family Solanaceae. - Int. J. Bot., 3(3): 276-282. ADEDEJI O. & JEWOOLA O.A. (2008): Importance of Leaf Epidermal Characters in the Asteraceae Family. - Not. Bot. Hort. Agrobot. 36 (2): 7-16. ADEDEJI O. (2010): Palynology of the genus Stachytarpheta Vahl. (Verbenaceae). - Not. Bot. Hort. Agrobot.. 2(4): 27-33. BALIK M.J. (1985): Note on some medicinal and poisonous plants of Amazonia Peru. Advance in Economic Botany, (1): 1-8. BEHNKE H.D. (1984): Plant trichomes structure and ultrastructure: general terminology, taxonomic applications and aspects of trichome bacterial interaction in leaf tips of Dioscorea. In: Biology and Chemistry of plant trichomes, RODRIGUEZ E., HEALEY P.L., MEHTA I., (eds.), New York: Plenum Press; 1-21. BERNAYS E.A. & CHAPMAN R.F. (1994): Host-Plant Selection by Phytophagous Insects. Chapman and Hall: New York. BHATIA R.C. (1984): Foliar Epidermal Studies of Heliotropium supinum L. - Folia Geobot. Phytotaxon, (19): 381-385. 12 CELKA Z., SZKUDLARZ P. & BIEREZNOJ U. (2006): Morphological variation of hairs in Malva alcea L. (Malvaceae). - Biodiversity Res. Conserv., 3: 258-261. DAVID H. & EASWARAMOORTHY S. (1988): Physical resistance mechanisms in insect plant interaction. In: Dynamics of Insects-Plant Interactions: Recent Advances and Future Trends (T.N. ANANTHAKRISHNAN, RAMAN A., eds.), New Delhi: Oxford and IBH Publishers; 45-70. DILCHER D.L. (1974): Approaches to the identification of angiosperms leaf remains. - Bot. Rev.,40(1): 1-157. DUFFEY S.S. (1986): Plant glandular trichomes: their partial role in defence against insects. In: Insects and the plant surface, JUNIPER B & SOUTHWOOD T.R.E, (eds.). Edward Arnold, London: 151-172. DUKE J.A. & VAZQUEZ M.R., (1994): Amazonia Ethnobotanical Dictionary (Peru), CRC Press: Boca Roton Fl, USA, p. 215. FAHN A. (1979): Secretory Tissues in Plants. Academic Press: London. FRANCINO D.M, (2006): Anatomia foliar e caulinar de Chamaecrista trichopoda (Caesalpinioideae) e histoquimica do nectario extrafloral. - Planta Daninha, (24): 695-705. GERSHENZON J. & CROTEAU R. (1991): Terpenoids. In: Herbivores: Their interactions with Secondary Plant Metabolites, Rosenthal G.A and Janzen D.H (eds.), Academic Press: New York; 151-172. GILL L.S. (1988). Taxonomy of Flowering Plants. Africana-FEP Publishers Limited: Bamenda, Cameroon; 388. GOTTLIEB O.R. & SALATINO A. (1987): Funcao e evolucao de oleos essenciais e suas estruturas secretoras. - Ciencia e Cultura, 39: 707-716. HARDIN J.W. (1979): Patterns of variation in foliar trichomes of eastern north American Quercus. American J. Bot. (6): 576-585. HAYAT M.Q.; ASHRAF M.; KHAN M.A.; YASMIN G.; SHAHEEN N. & JABEEN S. (2009): Phylogenetic Relationships in Artemisia spp. (Asteraceae) Based on Distribution of Foliar Trichomes. - Int. J. Agric. Biol., (11): 553-558. HESS F.D & FALK R.H. (1990): Herbicide deposition on leaf surfaces. - Weed Sci. 38(3): 280-288. nd HUTCHINSON J. & DALZIEL J.M., (1963): Flora of West Tropical Africa. (2 edn), vol 11, Crown Agents: London. IDU M.; ERHABOR J.O. & ODIA E.A. (2009): Morphological and Anatomical Studies of the Leaf and Stem of some Medicinal Plants: Stachytarpheta jamaicensis (L.) Vahl and S. cayennensis (L.C. Rich) Schau. - Ethnobotanical Leaflets, (13): 1417-1425. INAMDAR J.A. (1969): Epidermal structure and ontogeny of stomata in some Verbenaceae. - Ann. Bot.. 71(4):323-370. IYANG E. (2003): Ethnobotany. Conventional and Traditional Uses of Plants. The Verdict Press: Uyo Akwa-Ibom State Nigeria, p. 191. JEFFREE C.E. (1986): The cuticle, epicuticular waxes and trichomes of plants, with reference to their structure, function and evolution. In: Insects and the plant surface, JUNIPER B. & SOUTHWOOD T.R.E. (eds). Edward Arnold: London; 23-64. JOHNSON B. (1975): Plant pubescence: an ecological perspective. - Bot. Rev. (41): 233258. JONES J.H. (1986): Evolution of the Fagaceae: the implications of foliar features. Annl. Missouri Bot. Gard. (73): 228-275. JUDD W.S.; CAMPBELL C.S.; KELLOGG E.A. & STEVENS P.F. (1999): Plant Systematics – A Phylogenetic Approach. Sinauer Associates, Inc.: Sunderland, Massachusetts. JUNIPER B. & SOUTHWOOD T.R.E. (1986): Insects and the plant surface. Edward Arnold: London. 13 LEVIN D.A. (1973). The role of trichomes in plant defence. - Quart. Rev. Biol. 48: 3-15. LILLYAMMA M. & SHAH G.L. (1987): Anatomical contributions to the taxonomy of some Verbenaceae: Petiole. - Proc. Indian Acad. Sc. (Plant Sci.), 97(3): 235-246. MARQUIS R.J. (1992): The selective impact of herbivory. In: Plant Resistance to Herbivory and Pathogens. Ecology. Evolution and Genetics, FRITZ R.S & SIMMS E.L, (eds.). The University of Chicago Press: Chicago; 301-325. METCALFE C.R & CHALK L. (1950): Anatomy of the Dicotyledons, Vol. 1, Oxford. METCALFE C.R. (1960): Anatomy of monocotyledons. Vol. 1. Gramineae, Oxford University Press: Oxford; 61-66, 167-170. OGUNDIPE O.T & W UJEK D.E. (2004): Foliar anatomy on twelve genera of Bignoniaceae (Lamiales). - Acta Botanica Hungarica, 46(3-4): 337-361. PASSOS J.L; MEIRA R.M.S.A. & BARBOSA L.C.A. (2009): Foliar anatomy of the species Lantana camara and L. radula (Verbenaceae). - Planta Daninha, 27(4): 1-8. PETER A.J, SHANOWER T.G. & ROMEIS J. (1995): The role of plant trichomes in insects resistance: a selective review. - Phytophaga (Madras), 7: 41-64. PROCOPIO S.O. (2003): Leaf Anatomical Studies in Weed Species Widely Common in Brazil. 111 – Galinsoga parviflora, Crotalaria incana, Conyza bonariensis and Ipomoea cairica. - Planta Daninha, 21(1): 1-9. RAGHAVAN R.S. & ARORA C.M. (1960): Morphological and Cytological Studies in the Genus Lantana L. - Bull. Bot. Survey, 2: 299-303. RAMMAYA N. & RAO R.S. (1976): Morphology phylesis and biology of the peltate scale, stellate and tufted hairs in some Malvaceae. - J. Indian Bot. Soc., 55: 75-79. RAO R.S. & RAMAYYA N. (1977): Structure distribution and taxonomic importance of trichomes in the Indian species of Malvastrum. - Phytomorphology, 27: 40-44. RODRIGUEZ E.; HEALY P.L & MEHTA I. (1984): Biology and Chemistry of Plant Trichomes. Plenum: New York. ROMEIS J.; SHANOWER T.G & PETER A.J. (1999): Trichomes on Pigeon pea [Cajanus cajan (L) Millsp.] and two wild Cajanus spp. - Crop Sci. 39: 564-569. RUEDA R.M. (1993). The genus Clerodendron (Verbenaceae) in Mesomerica. - Ann. Missouri Bot. Garden, 80: 870-890. SCHWONTKOWSKI D. (1993): Herbs of the amazon. Traditional and common uses. Science Student Brain Trust Publishing: New York. SCOTLAND R.W.; OLMSTEAD R.G. & BENNETT J.R. (2003): Phylogeny Reconstruction, Role of Morphology. - Syst. Biol., 52: 539-548. SHAHEEN N.; KHAN M.A.; YASMIN G.; HAYAT M.Q, MUNSIF S. & AHMAD K. (2010): Foliar Epidermal Anatomy and Pollen Morphology of the Genera Alcea and Althaea (Malvaceae) from Pakistan. - Int.l. J. Agric .Biol., 12: 329-334. SINHA S. & SHARMA A. (1984): Lantana camara L. – A review. - Feddes Reppertorium, 5: 633-681. SMITH W.K; BELL D.T & SHEPHERD K.A. (1998): Associations between leaf structure, orientation and sunlight exposure in five western Australian communities. - Am. J. Bot., 85(1): 56-63. SOLEREDER H. (1908): Systematic anatomy of the Dicotyledons. Clarendon Press: Oxford; 643. THAIR S.S. & RAJPUT M.T. M. (2009): S.E.M. structure, distribution and taxonomic significance of foliar stomata in Sibbaldia L., species (Rosaceae). - Pak. J. Bot., 41(5): 2137-2143. THEOBALD W.L.; KRAHULIK J.L. & ROLLINS R. (1979): Trichome description and classification. In: Anatomy of the Dicotyledons, systematic anatomy of the leaf and stem. METCALFE C. R., CHALK L (eds.). Oxford Clarendon Press: Oxford, (1): 40-53. 14 VALKAMA E. (2003): Comparative analysis of leaf trichome structure and composition of epicuticular flavonoids in Finnish Birch species. - Ann. Bot., 91(6): 643-655. W AGNER, G.J.; W ANG E. & SHEPHERD, R.W. (2003): New Approaches for studying and exploiting an old protuberance, the plant trichome. – Annals of Botany 93(1): 3-11. W ERKER E. (2000): Trichome diversity and development. In: Advances in Botanical research, incorporating advances in plant pathology - Plant trichomes, HALLAHAN D.L., GRAY J.C., CALLOW J.A. (eds.). Academic Press: San Diego, 31: 1-35. W OODMAN R.L. & FERNANDES G.W. (1991): Differential mechanical defence: herbivory, evapotranspiration and leaf hairs. - Oikos, 60: 11-19. YAN-MING F. & RU-W EN F. (1993): Variation and evolution of leaf trichomes in Chinese Hamamelidaceae. - Acta Phytotaxon. Sin., 31: 147-152. Legend to Tables 1-3 X1 X2 X3 X4 X5 X6 X7 X8 X9 X10 X11 X12 X13 X14 X15 X16 X17 X18 X19 X20 X21 X22 X23 X24 X25 X26 Epidermal cell length (µm) Epidermal cell breadth (µm) Stomata size (µm²) Stomatal index (%) Vein cell length (µm) Vein cell breadth (µm) Spine-like non-glandular trichome length (µm) Spine-like non-glandular trichome breadth (µm) Unicellular non-glandulartrichome length (µm) Unicellular non-glandular trichome breadth (µm) Bicellular non-glandular trichome length (µm) Bicellular non-glandular trichome breadth (µm) Multicellular non-glandular trichome length (µm) Multicellular non-glandular trichome breadth (µm) 1 cell (unicellular) secretory head glandular trichome length (µm) 1 cell (unicellular) secretory head glandular trichome breadth (µm) 2 cells (bicellular) secretory head glandular trichome length (µm) 2 cells (bicellular) secretory head glandular trichome breadth (µm) 3 cells (tricellular) secretory head glandular trichome length (µm) 3 cells (tricellular) secretory head glandular trichome breadth (µm) 4 cells (multicellular) secretory head glandular trichome length (µm) 4 cells (multicellular) secretory head glandular trichome breadth (µm) 5-9 cells (multicellular) secretory head glandular trichome length (µm) 5-9 cells (multicellular) secretory head glandular trichome breadth (µm) 10-16 cells (multicellular) secretory head glandular trichome length (µm) 10-16 cells (multicellular) secretory head glandular trichome breadth (µm). 15 Tab. 1. Simple descriptive statistics of the quantitative characters of foliar epidermal surfaces of S. cayennensis. Variation X1 X2 X3 X4 X5 X6 X7 X8 X9 X10 X11 X12 X13 X14 X15 X16 X17 X18 X19 X20 X21 X22 X23 X24 X25 X26 Minimum Ad Ab 50.00 32.50 10.00 15.00 309.38 306.25 14.72 26.34 70.00 75.00 17.50 12.50 20.00 19.00 10.00 9.00 35.00 33.00 15.00 13.00 90.00 200.00 17.50 19.00 187.50 187.50 30.00 22.50 20.00 20.00 15.00 19.00 25.00 27.50 17.50 22.50 0 0 0 0 27.50 27.50 27.50 24.00 28.00 30.00 26.00 27.50 0 0 0 0 Maximum Ad Ab 87.50 77.50 45.00 37.50 687.50 687.50 21.77 36.93 162.50 140.00 37.50 20.00 30.00 28.00 17.50 18.00 65.00 60.00 25.00 23.00 135.00 240.00 37.50 25.00 452.50 252.50 52.50 25.00 32.50 31.00 25.00 25.00 27.50 32.00 22.50 26.00 0 0 0 0 36.00 35.00 30.00 28.00 36.00 35.00 28.00 30.00 0 0 0 0 Mean Standard deviation Standard error Ad Ab Ad Ab Ad Ab 66.40 55.20 11.34 10.82 2.27 2.16 27.70 25.90 7.84 5.44 1.57 1.08 483.24 489.75 101.34 112.59 20.27 22.52 17.98 31.72 1.91 2.80 0.38 0.56 116.60 112.30 29.96 19.87 5.99 3.97 26.00 17.60 5.99 2.55 1.99 0.51 27.00 26.00 4.47 3.90 2.00 1.90 14.00 13.80 2.85 2.05 1.27 1.15 51.50 49.50 13.87 12.00 6.20 5.80 20.00 19.00 3.95 3.00 1.77 1.46 122.00 219.00 18.57 14.32 8.31 6.40 31.40 21.80 8.32 2.59 3.72 1.16 309.00 215.50 129.98 33.88 58.13 15.15 38.50 23.60 9.78 1.29 4.37 0.58 24.50 26.80 5.42 4.44 2.42 1.98 19.50 21.20 3.70 2.39 1.66 1.07 27.00 29.30 1.12 1.96 0.50 0.87 20.00 24.10 1.77 1.43 0.79 0.64 0 0 0 0 0 0 0 0 0 0 0 0 32.00 30.50 4.17 3.32 1.86 1.48 28.40 26.50 1.08 1.87 0.48 0.84 31.80 32.00 3.49 2.12 1.56 0.95 27.00 28.70 0.71 0.97 0.32 0.44 0 0 0 0 0 0 0 0 0 0 0 0 Key: Ad - Adaxial, Ab - Abaxial Tab. 2. Simple descriptive statistics of the quantitative characters of foliar epidermal surfaces of S. angustifolia. Variation X1 X2 X3 X4 X5 X6 X7 X8 X9 X10 X11 X12 X13 X14 X15 X16 X17 X18 X19 X20 X21 X22 X23 X24 X25 X26 Minimum Ad Ab 55.00 57.50 30.00 25.00 525.00 375.00 24.72 28.87 75.00 90.00 17.50 17.50 0 0 0 0 0 0 0 0 220.00 235.0 28.00 27.00 0 237.50 0 32.50 35.00 35.00 35.00 35.00 0 0 0 0 27.50 34.00 27.00 27.50 32.50 32.50 32.50 35.00 30.00 35.00 28.00 37.50 50.00 50.00 47.50 50.00 Maximum Mean Standard Deviation Standard Error Ad Ab Ad Ab Ad Ab Ad Ab 112.50 112.50 88.44 88.10 16.67 14.90 3.33 2.98 72.50 50.00 50.70 36.50 13.43 5.91 2.69 1.18 650.00 568.75 614.23 505.25 49.12 56.12 9.82 11.23 31.34 33.64 28.88 31.14 1.80 1.31 0.36 0.26 192.50 182.50 130.30 126.80 34.22 28.17 6.84 5.63 32.50 30.00 26.10 23.50 3.54 2.80 0.71 0.56 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 232.00 245.00 227.90 241.40 4.69 4.16 2.10 1.86 32.00 29.00 29.60 27.90 1.52 0.74 0.68 0.33 0 495.00 0 325.50 0 101.62 0 45.44 0 55.00 0 48.50 0 9.29 0 4.15 38.00 37.50 36.10 36.00 1.52 1.37 0.68 0.61 40.00 40.00 37.20 38.00 1.89 2.09 0.85 0.93 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 33.00 40.00 30.40 36.90 2.33 2.41 1.04 1.08 29.00 40.00 27.80 33.90 0.76 5.92 0.34 2.65 40.00 43.00 36.50 36.60 2.85 3.99 1.27 1.78 42.50 41.00 35.00 38.30 4.33 2.33 1.94 1.04 33.00 41.00 32.00 38.40 1.17 2.30 0.52 1.03 30.00 40.00 29.00 38.70 1.00 0.97 0.45 0.44 59.00 59.00 55.90 54.80 3.61 3.27 1.62 1.46 55.00 56.00 52.00 53.00 3.26 2.24 1.46 1.00 Key: Ad - Adaxial, Ab - Abaxial 16 Tab. 3. Simple descriptive statistics of the quantitative characters of foliar epidermal surfaces of S. indica. Variation X1 X2 X3 X4 X5 X6 X7 X8 X9 X10 X11 X12 X13 X14 X15 X16 X17 X18 X19 X20 X21 X22 X23 X24 X25 X26 Minimum Maximum Mean Standard deviation Standard error Ad Ab Ad Ab Ad Ab Ad Ab Ad Ab 65.00 55.00 137.50 155.00 104.70 103.90 18.85 20.99 3.77 4.20 25.00 27.50 60.00 67.50 38.70 39.80 7.22 8.84 1.44 1.77 568.75 407.50 987.50 937.50 771.44 710.75 122.35 127.79 24.47 25.56 10.71 22.73 19.40 30.43 15.93 26.93 2.80 2.01 0.56 0.40 77.50 100.00 175.00 210.00 134.00 165.50 29.32 34.47 5.86 6.89 22.50 20.00 35.00 37.50 26.80 25.40 3.50 3.66 0.70 0.73 20.00 0 30.00 0 23.00 0 4.47 0 2.00 0 12.00 0 15.00 0 13.40 0 1.47 0 0.66 0 27.50 27.50 62.00 32.00 47.30 29.10 15.11 1.88 6.76 0.84 15.00 9.00 25.00 11.00 21.00 10.00 3.81 0.71 1.70 0.32 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 35.00 33.50 42.00 42.50 39.60 37.40 2.70 4.46 1.21 2.00 37.00 22.00 40.00 28.00 39.20 24.00 1.30 2.55 0.58 1.14 35.00 33.00 42.50 42.50 38.00 37.60 4.11 4.55 1.84 2.03 25.00 22.00 30.00 30.00 26.50 25.80 2.24 3.19 1.00 1.43 35.00 33.00 43.00 45.00 39.60 37.80 4.20 5.36 1.88 2.40 25.00 28.00 30.00 30.00 27.50 29.60 1.80 0.89 0.81 0.40 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 Key: Ad - Adaxial, Ab - Abaxial Tab. 4. Species grouping from Duncan’s Multiple Range Test (Means with the same letter are not significantly different) Character Epidermal cell length (µm) Epidermal cell breadth (µm) Stomatal size (µm²) Stomatal Index (%) Vein cell length (µm) Vein cell breadth Species name S. indica S. angustifolia S. cayennensis S. indica S. angustifolia S. cayennensis S. indica S. angustifolia S. cayennensis S. indica S. angustifolia S. cayennensis S. indica S. angustifolia S. cayennensis S. indica S. angustifolia S. cayennensis 17 Grouping Adaxial 104.70 A 88.44 B 66.40 C 50.70 A 38.70 B 27.70 C 771.44 A 614.23 B 483.24 C 28.88 A 17.98 B 15.93 C 134.00 A 130.30 A 116.60 A 26.80 A 26.10 A 26.00 A Abaxial 103.90 A 88.10 B 55.20 C 36.50 A 39.80 A 25.90 B 710.75 A 505.25 B 489.75 B 31.14 A 31.72 A 26.73 B 165.50 A 126.80 B 112.30 B 25.40 A 23.50 B 17.60 C Tab. 5: Summary of the qualitative foliar epidermal characters Species Epidermal cells shape Adaxial Abaxial S. cayennensis Polygonal Polygonal Regularly Irregularly arranged arranged S. angustifolia Polygonal Polygonal Irregularly Irregularly arranged arranged Polygonal Polygonal S. indica Irregularly Irregularly arranged arranged Anticlinal cell wall Epidermal cells overlaying veins Adaxial Abaxial Adaxial Abaxial Straight to Sinuous Polygonal Polygonal wavy Stomatal type Adaxial Diacytic Anisocytic Anomocytic Undulating Sinuous Polygonal to Polygonal to Diacytic to sinuous rectangular rectangular Anisocytic Anomocytic Undulating Sinuous Polygonal Polygonal to Diacytic to sinuous rectangular Anisocytic Anomocytic Stomatal shape Abaxial Diacytic Anisocytic Anomocytic Diacytic Anisocytic Anomocytic Diacytic Anisocytic Anomocytic Crystals Adaxial Abaxial Adaxial Abaxial Circular Elliptic to + + to elliptic circular Elliptic Elliptic + + Elliptic Elliptic + + Key: + present 18 Tab. 6: Leaf venation studies; minimum (mean ± standard error) maximum. Characters Venation type: Primary veins Secondary veins Areoles: Areole shape Areole Length (µm) Areole Width (µm) Veinlet termination: Number per areole Description S. cayennensis S. angustifolia S. indica Pinnate craspedodromous simple Recurved Pinnate craspedodromous simple Recurved Pinnate craspedodromous simple Recurved Polygonal, occasionally rectangular 260(372±33.38)450 200(314±40.69)430 Polygonal, occasionally triangular or rectangular 230(502±76.90)700 180(310±48.89)410 Polygonal, occasionally rectangular. 310(572±91.24)830 200(244±20.15)310 0-2 Simple to branched 0–4 Often branched 0ften 0, occasionally 1 – 2. Simple to branched Tab. 7: Trichome density on the foliar epidermal surfaces of the Stachytarpheta species. Trichome type Non-glandular Glandular Species S. cayennensis S. angustifolia S. indica S. cayennensis S. angustifolia S. indica Ad 5 0 0 0 4 0 Minimum Ab 0 0 0 1 6 1 Ad 10 1 1 3 17 1 Maximum Ab 3 1 1 6 18 2 Ad 7.2 0.2 0.6 1.6 8.6 0.4 Mean Ab 0.6 0.2 0.2 4.2 10.2 1.4 Standard deviation Ad Ab 1.92 1.34 0.45 0.45 0.55 0.40 1.34 3.11 5.18 2.97 0.55 0.55 Standard Error Ad Ab 0.86 0.60 0.20 0.20 0.24 0.20 0.60 1.39 2.32 1.33 0.24 0.24 Key: Ad - Adaxial, Ab - Abaxial Tab. 8: Distribution of trichomes on the different organs of the Stachytarpheta species Species 19 Adaxial epidermal surface S. cayennensis A, B, C, D, E, F, H, K* S. angustifolia C, E, G, H, I, J A, E, F, G S. indica Key: A B C D E F G H I J K L Abaxial epidermal surface A, B, C, D, E, F, H, I D, H, I E, F, G Midrib Petals Petal tube C, D C, D, F, K C, D, F. B, K L C, D, G, H, I B, C, D, H A, D, F, I B, E, K L C, D, F C, D, F, K B, D, E, F B, K K, L Spine-like non-glandular trichome Unicellular non-glandular trichome Bicellular non-glandular trichome Multicellular non-glandular trichome 1 cells (unicellular) secretory head glandular trichome 2 cells (bicellular) secretory head glandular trichome 3 cells (tricellular) secretory head glandular trichome 4 cells (multicellular) secretory head glandular trichome 5-9 cells (multicellular) secretory head glandular trichome 10-16 cells (multicellular) secretory head glandular trichome Short stalk secretory head glandular trichome Long stalk secretory head glandular trichome. Petiole Stem Calyx Veins A, D, F. A, B, C, D, F. A, B, C, G, H A, B, C, D, F, H, I A, E, F A, C, D, F A B C D Fig. 1. S. cayennensis (A-D, X400): A. Adaxial epidermal surface (arrow indicates anisocytic stomatal type); B. Abaxial epidermal surface (arrow indicates diacytic stomata, double arrow indicates anomocytic stomatal type); C. Adaxial epidermal surface (arrow indicates anomocytic stomatal type); D. Adaxial epidermal surface showing contiguous stomata. 20 A B C F D E G Fig. 2. S. cayennensis (A-C, E-G X400; D, X100): A. Spine-like non-glandular trichome; B. Bicellular non-glandular trichome on petiole; C. Bicellular nonglandular trichome on adaxial epidermal surface; D. Tricellular (Multicellular) nonglandular trichomes on the midrib; E. Multicellular non-glandular trichome with the basal cell shrivelled on petiole and stem; F. Multicellular non-glandular trichome on stem and veins; G. Multicellular non-glandular trichome on calyx. 21 A B C E D F G Fig. 3. S. cayennensis (A-G, X400): A. cells secretory head glandular trichome on adaxial epidermal surface; B. 2 cells secretory head glandular trichome on abaxial epidermal surface; C. short stalk secretory head glandular trichome on adaxial epidermal surface (arrow indicates short stalk); D. short stalk, 4 cells secretory head glandular trichome on petiole and petal; E. Oblong crystals on petals and epidermal surfaces;F. Spherical crystals on petals and epidermal surfaces; G. Rectangular crystals on petals and epidermal surfaces. 22 B A E C D F G Fig. 4. S. cayennensis (A-G, X400): A. Unicellular non-glandular trichome on petal; B., D., G. Short stalk, 2 cells secretory head glandular trichome on petal; C. Short stalk (with arrow), 5 cells secretory head glandular trichome on petal; E. Bicellular non-glandular trichome on petal; F. Short stalk, 4 cells secretory head glandular trichome on petal. 23 A C B D E F G H I Fig. 5. S. angustifolia (A-F, H-I, X400; G, X100): A. Adaxial epidermal surface (arrow indicates diacytic stomata, double arrow, anisocytic stomata); B. Abaxial epidermal surface (arrow indicates diacytic stomatal type, double arrow anisocytic stomata type); C.-D. Anomocytic stomatal type on abaxial epidermal surface; E. Trichome base on abaxial epidermal surface (with the base of a trichome); F. Vein cells on abaxial surface; G. Venation pattern (arrow indicates branched veinlet termination); H. Unicellular non-glandular trichome on adaxial epidermal surface; I. Bicellular non-glandular trichome on adaxial epidermal surface. 24 A B C D F E G H Fig. 6. S. angustifolia (A-H, X400): A. Tricellular (multicellular) non-glandular trichome on midrib; B. Multicellular non-glandular trichomes on midrib and stem; C. 1 cell secretory head glandular trichome on epidermal surfaces; D. 3 cells secretory head glandular trichome on epidermal surfaces, midrib and calyx; E. 4 cells secretory head glandular trichome on epidermal surfaces, midrib, petiole, calyx and vein; F. 6 cells secretory head glandular trichome on epidermal surfaces; G. 8 cells secretory glandular trichome on epidermal surfaces; H. 16 cells secretory head glandular trichome on epidermal surfaces. 25 A B D G C E F H I Fig. 7. S. angustifolia (A-I, X400): A. 1 cell secretory head glandular trichome on petals; B.-C. Short stalk (faint, with arrow) 8 cells secretory head glandular trichome on petals; D. Short stalk (faint, with arrow) 5 cells secretory head glandular trichome on petals; E. Short stalk, 1 cell secretory head glandular trichome on petals; F. Short stalk, 2 cells secretory head glandular trichome on petals; G. Short stalk, 3 cells secretory head glandular trichome on petals; H. Short stalk (hooked), 5 cells secretory head glandular trichome on petals; I. Unicellular, non-glandular trichome on petals. 26 B A C E D F G Fig. 8. S. angustifolia (A.-G., X400): A. Long stalk, 1-2 cells secretory head glandular trichome on petal tube; B. Short stalk 2-6 cells secretory head glandular trichome on petals; C. Groups of small crystals on petal tube (arrow indicates crystals); D. Groups of small crystals on epidermal surfaces (arrows indicate crystals); E. Crystals on petal with short stalk, 2 cells secretory head glandular trichome; F. Spherical crystal on stem and epidermal surfaces; G. Rectangular crystal on epidermal surfaces. 27 A B C D F E G H I Fig. 9. S. indica (A-G, I, X400; H, X100): A. Adaxial epidermal surface (arrow indicates diacytic stomatal type); B. Abaxial epidermal surface (arrow indicates aborted guard cell); C. Abaxial epidermal surface with anisocytic (with arrow) stomatal type on abaxial surface; D. Abaxial epidermal surface with diacytic (with arrow) and anisocytic (double arrow) stomatal types; E. Abaxial epidermal surface with normal snd aborted guard cells; F. Adaxial epidermal surface with spine-like non-glandular trichome; G. 1 cell secretory head glandular trichome on epidermal surfaces; H. Venation pattern with 0 (arrow) and 1 (double arrow) veinlet termination in areole; I. Vein cells on adaxial epidermal surface. 28 A B D C E F G Fig. 10. S. indica (A-H, X400): A. Spine-like non-glandular trichome on adaxial and abaxial surfaces; B. Unicellular non-glandular trichome on stem; C. Bicellular nonglandular trichome on midrib, petiole and veins; D. Multicellular non-glandular trichome on stem (terminal cell shrivelled); E. Multicellular non-glandular trichome on stem (1 cell shrivelled); F. Multicellular non-glandular trichome on petiole; G. Multicellular non-glandular trichome on midrib. 29 A D G B C E F H I Fig. 11. S. indica (A-I, X400): A. 2 cells secretory head glandular trichome on epidermal surfaces; B. Short stalk, 5-7 cells secretory head glandular trichome (stalk faint, with arrow), on petals; C.-D. Short stalk, 1-2 cells secretory head glandular trichome on petals and petioles; E. Short (hooked) stalk, 2 cells secretory head glandular trichome on petals; F. Spherical crystal on abaxial epidermal surfaces with a stomata with aborted guard cells; G. Small spherical crystals with short stalk 1 cell secretory head glandular trichome on petal tube; H. Short stalk, 3 cells secretory head glandular trichome on petals; I. Long stalk, 1 cell secretory head glandular trichome on petal tube. 30 3 2 1 1 3 2 0 1 2 3 0 -10 -10 -20 -20 -30 Similarity Similarity -30 -40 -40 -50 -50 -60 -60 -70 -70 -80 1 2 3 4 Fig. 12. Dendrogram of the adaxial epidermal surface and trichome characters. Key: 1 S. cayennensis 2 S. angustifolia 3 S. indica 4 Fig. 13: Dendrogram of the abaxial epidermal surface and trichome characters. Key: 1 S. cayennensis 2 S. angustifolia 3 S. indica Received: Revised: Accepted: 31 nd March 02 2011 th November 11 2011 th November 27 2011