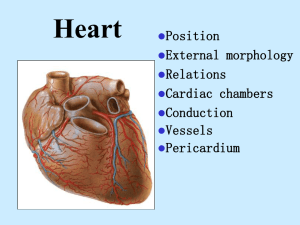
Cardiovascular Embryology
advertisement

Pediatr Cardiol 25:191–200, 2004 DOI: 10.1007/s00246-003-0585-1 Cardiovascular Embryology R. Abdulla,1 G. A. Blew,2 M.J. Holterman3 1 2 3 Pediatric Cardiology, The University of Chicago. MC4051, 5841 S. Maryland Ave., Chicago, IL 60637-1470, USA School of Biomedical Visualization, University of Illinois at Chicago, 840 S. Wood Street, Chicago, IL 60637, USA Department of Surgery, University of Illinois at Chicago, 840 S. Wood Street, Chicago, IL 60637, USA Abstract. During the first 20 days of development, the human embryo has no cardiovascular structure. Over the next month, the heart and great vessels complete their development and look very much like they will at full gestation. This amazing process transforms isolated angiogenic cell islets into a complex, four-chambered structure. During this transformation, the single heart tube begins to beat at 23 days of development and by 30 days blood circulates through the embryo. Keywords: Heart — Cardiovascular — Embryology — Primitive heart — Heart looping — Outflow tract septation This review of human embryology attempts to document the many different, and sometimes disputing, theories of the development of the heart and its great vessels. The goal is to provide a broad spectrum and detailed information for those interested in the field of pediatric cardiology. Many details were intentionally left out, such as molecular biology issues, because it is impossible to include this ever-expanding topic together with morphogenesis in one article. Many publications are available for understanding molecular biology and neural crest involvement in the development of the cardiovascular system [11, 12, 14, 15, 17, 23, 24, 32–35, 38]. It is difficult to describe or use two-dimensional (2-D) imagery when describing a three-dimensional (3-D) object. Despite this fact, we continue to describe in our literature, lectures, and conferences the heart using 2-D terminology and illustrations, expecting the audience to recreate a mental 3-D figure. Unfortunately, the inability to conceive what is being described is frequent, leading to confusion, the need Correspondence to: R. Abdulla, email: rabdulla@peds.bsd. uchicago.edu for repetition and elaboration, or, worse, misunderstanding and error. Pediatric cardiologists, particularly those in training, frequently realize when examining a heart from an autopsy that their understanding of spatial relationship of cardiac structures of that particular lesion was wrong. This difficulty becomes even more immense when dealing with a 3-D object in a state of continual and complex change, such as that of the cardiovascular system during its embryological development. Therefore, it becomes increasingly useful to depict these changes with four-dimensional imagery (i.e., computer animations depicting 3-D structures changing over time). The task of preparing these animations is enormous, requiring expertise in computer medical illustration and mastery over userhostile software. This is possible for only a few of us, and even then it is time-consuming and costly. The use of computer-generated 3-D images and animations in the field of cardiac embryology is becoming more frequent. This technique is implemented in research as well as to create educational images [1, 13, 19–21, 45]. In the Internet version of this article, movie animations demonstrating cardiovascular development are presented. Embryonic folding, heart tube looping, and development of systemic venous drainage are demonstrated in different movie animations. These images were created using current information about the development of these structures. On the other hand, a different animation shows a process that can be used to create 3-D objects using histological slices from human embryos. Stage 14 sliced embryos from the Carnegie collection of human embryos from the National Library of Medicine in Washington, DC, were digitized, the cardiovascular structures were traced, and the various slices were then stacked up using special computer software. This animation demonstrates how actual 3-D structures can be scientifically reassembled for better understanding 192 Pediatric Cardiology Vol. 25, No. 3, 2004 ential growth causing the embryo to fold in two different dimensions: 1. Craniocaudal axis due to the more rapid growth of the neural tube forming the brain at its cephalic end. Growth in this direction will cause the embryo to become convex shaped. 2. Lateral folding, causing the two lateral edges of the germ disk to fold forming a tube-like structure. Fig. 1. The Carnegie collection of embryos includes various stages of whole and sliced embryos. Digital images of slides of sliced embryos are made, with various structures traced using specialized software. Subsequently, 3-D images are electronically reconstructed. This image depicts a slice from a stage 14 embryo with 3-D reconstruction, demonstrating the dorsal half of the embryo (white) as well as a 3-D reconstruction of the heart. See animation of this process in the Web version of this issue. (Fig. 1). After 3-D cardiac structures from sequentially staged embryos are created, the images can serve as templates for the animation process. These can then be studied from various vantage points and provide embryologically correct teaching tools to facilitate the comprehension of cardiac development (Fig. 2). The first indication of any cardiovascular development occurs on approximately day 18 or 19. Prior to embryonic folding, angiogenic cell clusters on either side of the neural crest coalesce to form capillaries in the mesoderm of the germ disk. These capillaries then join to form a pair of blood vessels on each side of the neural crest (total of four blood vessels). These blood vessels run along the long axis of the germ disk, with one pair of blood vessels at the lateral edge of the embryo (one on each edge) and the other pair more medially on either side of the neural tube. The blood vessels on either side of the neural tube join at their cranial end. As the embryo folds in its lateral dimension, it causes the lateral edges of the germ disk to approach each other until they meet, causing the embryo to acquire a tubular form [16, 25]. The two outer endocardial tubes will come close to each other in the median of the embryo, ventral to the primitive gut, and start fusing cranially to caudally, thus forming a single median tube—the primitive heart tube [16, 41]. The Primitive Heart Embryonic Folding Early in the third week of development, the germ disk has the appearance of a flat oval disk and is composed of two layers: the epiblast and the hypoplast. The first faces the amniotic cavity and the latter faces the yolk sac. A primitive groove, ending caudally with the primitive pit surrounded by a node, first appears at approximately 16 days of development and extends half the length of the embryo. The primitive groove serves as a conduit for epiblast cells that detach from the edge of the groove and migrate inwards toward the hypoblast and replace it to form the endoderm. After the endoderm is formed, cells from the epiblast continue to migrate inwards to infiltrate the space between the epiblast and the endoderm to form the intraembryonic mesoderm. After this process is complete, the epiblast is termed the ectoderm [16, 25, 37] (Fig. 3). The flat germ disk transforms into a tubular structure during the fourth week of development [16, 25, 35]. This is achieved through a process of differ- The first intraembryonic blood vessels are noted on day 20, and 1–3 days later the formation of the single median heart tube is complete. The heart starts to beat on day 22, but the circulation does not start until days 27–29 [35]. The single tubular heart develops many constrictions outlining future structures. The cranialmost area is the bulbus cordis, which extends cranially into the truncus arteriosus. This, in turn, is connected to the aortic sac and through the aortic arches to the dorsal aorta [35]. The primitive ventricle is caudal to the bulbus cordis and the primitive atrium is the caudal-most structure of the tubular heart. The atrium connects to the sinus venosus, which receives the vitelline veins (from the yolk sac) and common cardinal (from the embryo) and umbilical (from primitive placenta) veins. The primitive atrium and sinus venosus lay outside the caudal end of the pericardial sac, and the truncus arteriosus is outside the cranial end of the pericardial sac. Some publications have introduced new terminology describing the segments of the primitive heart. Wenink and Gitten- R. Abdulla et al.: Cardiovascular Embryology 193 Fig. 2. The sequence of events resulting in the union of the two lateral endocardial tubes to form the single endocardial tube. The rest of the embryo is not shown. The embryo starts as a flat disk(A). The lateral endocardial vessels located on either side of a flat embryo disk come closer together as the embryo folds along its long axis to transform a flat structure into a tubular shape (B). As the edges of the flat embryo meet to form this tubular structure, the two lateral endocardial vessels unite (C), forming a single heart tube at the ventral aspect of the embryo (D). This process occurs on approximately day 20 or 21 of development. See animation of this process in the Web version of this issue. Fig. 4. The single heart tube shows constrictions outlining future structures. Fig. 3. Cells from the epiblast detach and migrate through the primitive groove to form the endoderm and mesoderm layers. berger-deGroot [44] support the use of inlet, outlet, and arterial segments as proposed by Anderson and Becker [3, 4, 10] (Fig. 4). Looping of the primitive heart occur on approximately day 23 of development [22]. It was initially suggested that this is due to faster growth of the bulboventricular portion of the heart compared to the pericardial sac and the rest of the embryo [35]. However, it has been shown that the heart will loop even when the pericardial sac is removed, as seen when the heart is cultured in vitro [24, 41]. It seems that the process of looping is a genetic property of the myocardium and not related to differential growth [41]. As the heart tube loops, the cephalic end of the heart tube bends ventrally, caudally, and slightly to the right. The bulboventricular sulcus becomes visible from the outside, and from the inside a primitive interventricular foramen forms. The internal fold formed by the bulboventricular sulcus is known as the bulboventricular fold. The bulboventricular seg- ment of the heart is now U shaped; the bulbus cordis forms the right arm of the U-shaped heart tube and the primitive ventricle forms the left arm. The looping of the bulboventricular segment of the heart will cause the atrium and sinus venosus to become dorsal to the heart loop [41]. At this stage, the paired sinus venosus extends laterally and gives rise to the sinus horns. As the cardiac looping progresses, the paired atria form a common chamber and move into the pericardial sac. The atrium now occupies a more dorsal and cranial position and the common atrioventricular junction becomes the atrioventricular canal, connecting the left side of the common atrium to the primitive ventricle [35]. At this stage, the heart has a smooth lining except for the area just proximal and just distal to the bulboventricular foramen, where trabeculations form. The primitive ventricle will eventually develop into the left ventricle and the proximal portion of the bulbus cordis will form the right ventricle. The distal part of the bulbus cordis, an elongated structure, will form the outflow tract of both ventricles, and the truncus arteriosus will form the roots of both great vessels. The bulbus cordis gradually acquires a more medial position due to the 194 Pediatric Cardiology Vol. 25, No. 3, 2004 Fig. 5. Looping of the single endocardial heart tube transforms it into a complex fourchamber structure. Looping starts on day 23 of development, and the four-chambered heart is evident on day 27. growth of the right atrium, forcing the bulbus to be in the sulcus in between the two atria [42] (Fig. 5). and the right umbilical vein connects to the vitelline system through the ductus venosus (which is derived from the vitelline veins) [26] (Fig. 6). Systemic Venous System Pulmonary Circulation On day 21, there is a common atrium as a result of fusion of the two endocardial tubes. The common atrium communicates with two sinus horns, a left and a right horn, representing the unfused ends of the endocardial tubes [16]. These two horns will form the sinus venosus. The sinus venosus is located dorsal to the atria. The following veins drain into the sinus venosus on each side: the common cardinal vein, which drains from the anterior cardinal vein (draining the cranial part of the embryo); the posterior cardinal vein (draining the caudal part of the embryo); the umbilical vein (connecting the heart to the primitive placenta); and the vitelline vein (draining the yolk sac, gastrointestinal system, and the portal circulation). On week 4, the sinus venosus communicates with the common atrium. During week 7, the sinoatrial communication becomes more right sided, connecting it to the right atrium. At 8 weeks, the distal end of the left cardinal vein degenerates, and the more proximal portion of it now connects through the anastomosing vein (left brachiocephalic vein) to the right anterior cardinal vein (right brachiocephalic vein), thus forming the superior vena cava. The left posterior cardinal vein also degenerates, and the left sinus horn receiving venous blood from the heart becomes the coronary sinus. The right vitelline vein becomes the inferior vena cava, and the right posterior cardinal vein becomes the azygos vein. All this is completed in week 8 of development. The left umbilical vein degenerates Airways, Lung Parenchyma, and Distal Pulmonary Arteries On day 21 of development, a groove forms in the floor of the foregut just dorsal to the heart. This is termed the pharyngeal groove, which develops to form the pharynx. On day 23, the laryngotracheal groove, a median structure in the pharyngeal region, develops. The edges of the laryngotracheal tube fuse to form the larynx and trachea cranially and the right and left main bronchi and right and left lung buds distally. The growth and branching of the lung buds, together with the surrounding mesoderm, form the distal airways, lung parenchyma, and pulmonary blood vessels. By week 16 of gestation, a full complement of preacinar airways and blood vessels have formed. The pulmonary arteries in utero are muscular, similar to that of the aorta. The thick, muscular walls of pulmonary arteries extend much further into distal arteries than what is seen in adults. Thinning of distal pulmonary arteries occurs postnatally as the pulmonary vascular resistance decreases after the onset of breathing and improved oxygenation [29]. Proximal Pulmonary Arteries The proximal main pulmonary artery develops from the truncus arteriosus, whereas the distal main pul- R. Abdulla et al.: Cardiovascular Embryology 195 Fig. 6. Development of the systemic venous drainage. These schematics represent dorsal views of the heart. (a) At week 4 of development, there is symmetrical systemic venous drainage into the two sinus venosus horns. (b) At week 7 of development, there is degeneration of some of the systemic veins. (c) At week 8 of development, the central systemic venous anatomy as seen in a term infant. Normal and abnormal development of systemic venous drainage are shown in movie clips in the Web version of this issue. IVC, inferior vena cava; SVC, superior vena cava. monary artery and the proximal right pulmonary artery develop from the ventral sixth aortic arch artery. The distal right pulmonary artery and the left pulmonary arteries form from the post branchial arteries, which develop from the lung buds and surrounding mesoderm. The ductus arteriosus develops from the distal left sixth aortic arch artery. Pulmonary Venous System A primitive vein sprouts out of the left atrium, which bifurcates twice to give four pulmonary veins that grow toward the developing lungs. The lung buds develop from the foregut. A plexus of veins is formed in the mesoderm enveloping the bronchial buds; these veins will meet with the developing pulmonary veins out of the left atrium to establish a connection during week 5 of gestation. As the left atrium develops, it progressively incorporates the common pulmonary vein into the left atrial wall until all four pulmonary veins enter the posterior wall of the left atrium separately. The incorporated pulmonary veins form the smooth posterior wall of the left atrium, whereas the trabeculated portion of the left atrium comes to occupy a more ventral aspect [16, 35]. Atrioventricular Canal The atrioventricular valves form during the fifth to eighth week of development [26]. Initially, endocardial cushion tissue forms bulges at the atrioventricular junction. These bulges have the appearance of valves, and although such tissue may play an im- portant role in the eventual formation of the atrioventricular valves, endocardial cushion tissues are not the precursors of the mitral and tricuspid valves [16, 43]. The atrioventricular junction is guarded by two masses of endocardial cushions—a superior and inferior cushion. These two masses will meet in the middle, thus dividing the common atrioventricular canal into right and left atrioventricular orifices. The process through which these two cushions fuse is not clear [18], and the role of apoptosis in this process is debatable. The fusion of the two endocardial cushions results in the formation of two atrioventricular orifices. In addition, the atrioventricular cushion appears to play a role in the closure of the interatrial communication at the edge of the primum atrial septum. This septum grows toward the atrioventricular endocardial cushion and fuses with it [41]. The formation of the atrioventricular valve starts when the atria and inlet portion of the ventricle enlarge; the atrioventricular junction (or canal) lags behind. Such a process causes the sulcus tissue to invaginate into the ventricular cavity, forming a hanging flap. The endocardial cushion tissue is located at the tip of this flap, which is formed from three layers—the outer layer from atrial tissue, the inner layer from ventricular tissue, and the middle layer from invaginated sulcus tissue. The inlet portion of the ventricles then becomes undermined, forming the tethering cords holding the newly formed valve leaflets. The inner sulcus tissue will eventually come in contact with the cushion tissue at the tip of valve leaflets, thus interrupting the muscular continuity between the atria and ventricles [16] (Fig. 7). 196 Fig. 7. Formation of atrioventricular valves. The Atria and Atrial Septum The atria of the mature heart have more than one origin. The trabeculated portions (appendages) of the right and left atria are from the primitive atria, whereas the smooth-walled posterior portions of the left and right atria originate from the incorporation of venous blood vessels. The posterior aspect of the left atrium is formed by the incorporation of the pulmonary veins, whereas the posterior smooth portion of the right atrium is derived from the sinus venosus. The two sinus horns are initially paired structures; later, they fuse to give a transverse sinus venosus. The entrance of the sinus venosus shifts rightward to eventually enter into the right atrium exclusively. The veins draining into the left sinus venosus (left common cardinal, umbilical, and vitelline veins) eventually degenerate. The left sinus venosus will become smaller because it will drain only the venous circulation of the heart, becoming the coronary sinus. The sinus venosus orifice of the right atrium is slit-like and to the right of the undeveloped septum primum [16]. The sinus venosus now connecting to the right atrium will assume a more vertical position. The sinoatrial junction will become guarded by two valve-like structures, resulting from the invagination of the atrial wall at the right and left sinoatrial junction. This orifice enlarges, with the superior and inferior vena cavae and the coronary sinus opening separately and directly into the right atrium. The right and left sinoatrial valves join at the top, forming the septum spurium. This septum and the two sinoatrial valve-like structures obliterate and are not appreciated in the mature heart [41]. Atrial septation starts when the common atrium becomes indented externally by the bulbus cordis and truncus arteriosus. This indentation will correspond internally with a thin sickle-shaped membrane developing in the common atrium on day 35 [39]. This membrane divides the atrium into right and left chambers. It grows from the posterosuperior wall and extends toward the endocardial cushion of the atrio- Pediatric Cardiology Vol. 25, No. 3, 2004 ventricular canal. This is the septum primum. The septum primum initially has a concave-shaped edge growing toward the atrioventricular canal. This orifice connecting the two atria is called the ostium primum. As the superior and inferior endocardial cushions fuse, thus dividing the atrioventricular canal into a right and left orifice, the concave lower edge of the septum primum fuses with it, obliterating the ostium primum. However, just before this happens fenestrations appear in the posterosuperior part of the septum forming the ostium secundum, thus maintaining a communication between the two atria [41]. The ostium secundum and superior vena cava later acquire a more anterosuperior position, although they maintain their relationship with each other; this is achieved through the growth of the atria [41]. These fenestrations then coalesce and form a larger fenestration. Meanwhile, another sickle-shaped membrane develops on the anterosuperior wall of the right atrium, just right of the septum primum and left of the sinus venosus valve. It grows and covers the ostium secundum, which continues to allow blood passage since the two membranes do not fuse. The septum secundum grows toward the endocardial cushion, leaving only an area at the posterosuperior part of the interatrial septum where the septum primum continues to exist as the foramen ovale membrane. The septum primum disappears from the posterosuperior portion of interatrial septation and the edge of the septum secundum forms the rim of the fossa ovalis [44] on approximately day 42 of development (Figs. 8 and 9). Ventricular Septation Ventricular septation is a complex process involving different septal structures from various origins and positioned at various planes [2, 27, 28, 31]. These structures eventually meet to complete the separation of the right and left ventricles. Muscular Interventricular Septum During the fifth week, on approximately day 30, a muscular fold extending from the anterior wall of the ventricles to the floor appears at the middle of the ventricle near the apex and grows toward the atrioventricular valves with a concave ridge. Most of the initial growth is achieved by growth of the two ventricles on either side of the ventricular septum. In addition, trabeculations from the inlet region coalesce to form a septum, which grows into the ventricular cavity at a slightly different plane than that of the primary septum; this is the inlet interventricular septum, which is in the same plane of that of the R. Abdulla et al.: Cardiovascular Embryology 197 Fig. 8. The atrial septum is formed by the septum primum and septum secundum. A movie clip depicting this process can be viewed in the Web version of this issue. AV, atrioventricular; IVC, inferior vena cava; SVC, superior vena cava. Outflow Tract Septum Fig. 9. 3-D depiction of atrial septum formation. See animation in Web version of this issue. atrial septum. The point of contact between these two septa will cause the edge of the primary septum to protrude slightly into the right ventricular cavity, forming the trabecular septomarginalis. The fusion of these two septa forms the bulk of the muscular interventricular septum. This septum will then come into contact with the outflow septum (Fig. 10). The interventricular foramen, which is bordered by the concave upper ridge of the muscular interventricular septum, the fused atrioventricular canal endocardial tissue, and the outflow tract septation ridges, never actually closes. Instead, communication between the left ventricle and the right ventricle is closed at the end of week 7 by growth of three structures—the right and left bulbar ridges and the posterior endocardial cushion tissue—that baffle the left ventricular output through a newly formed left ventricular outflow tract (LVOT). The LVOT is posterior to a right ventricular outflow tract, connecting the right ventricle to the pulmonary trunk. The cardiac outflow tract includes the ventricular outflow tract and the aortopulmonary septum. There has been much debate regarding this process. This section provides a summary of various theories [9, 36, 40]. In 1942, Kramer suggested that there are three embryological areas: the conus, the truncus, and the pulmonary arterial segments. Each segment develops two opposing ridges of endocardial tissue; the opposing pairs of ridges and those from various segments meet to form a septum separating two outflow tracts and aortopulmonary trunks. The aortopulmonary septum is formed by ridges separating the fourth (future aortic arch) and the sixth (future pulmonary arteries) aortic arches. The truncus ridges are formed in the area where the semilunar valves are destined to be formed, thus forming the septum between the ascending aorta and the main pulmonary artery. The conus ridges form just below the semilunar valves and from the septation between the right and left ventricular outflow tracts. Van Mierop [41] agreed that there are three pairs of ridges forming in the aortopulmonary, truncus, and conus regions. However, he stated that the pairs of ridges fuse independently and later on fuse with each other to complete the septation. His theory indicates that the truncus ridges form first, and as they fuse they form a truncal septum. This septum then fuses with the aortopulmonary septum, which is formed by invagination of the dorsal wall of the aortic sac between the fourth and the sixth aortic arch arteries (Fig. 11). Asami [7], Pexieder [36, 37], and Orts Llorca et al. [7], concur with Van Mierop’s theory; however, Asami believes that these ridges fuse in the opposite direction of that indicated by Van Mierop (i.e., from the outflow tract to the aortopulmonary region). On the other hand, Pexieder and Orts Llorca believe that 198 Fig. 10. Formation of ventricular septum. there are only two septa—a conotruncal (or bulbar) and an aortopulmonary septum. In 1989, Bartlings et al. introduced a new theory. They stated that the septation process of the ventricular outflow tracts, pulmonary and aortic valves, and the great vessels is mostly caused by a single septation complex, which they termed aortopulmonary septum. This septation complex develops at the junction of the muscular ventricular outflow tract with the aortopulmonary vessel. This junction has a saddle shape, allowing the right ventricular outflow tract to be long with a short main pulmonary artery, whereas the left ventricular outflow tract becomes short with a long ascending aorta (Fig. 12). The ventricular outflow septation is formed by condensed mesenchyme, embedded in the endocardial cushion tissue just proximal to the level of the aortopulmonary valves. The condensed mesenchyme will come in close contact with the outflow tract myocardium, from the area just above the bulboventricular fold, and participate in the septation of the outflow tract by providing an analogue to muscle tissue [6–9]. Myocardium in contact with the mesenchymal arch grows rapidly and forms the bulk of the outflow septum, continuous with the primary fold on the parietal wall of the right ventricle and the myocardium on the right side of the primary septum. Conduction System Primary myocardium, found in the early heart tube, gives rise to the contracting myocardium (of the atria and ventricles) and the conducting myocardium (nodal and ventricular conducting tissue). Conducting myocardial tissue is frequently referred to as being highly specialized tissue, implying that it has a homogenous function. In reality, some portions, such Pediatric Cardiology Vol. 25, No. 3, 2004 Fig. 11. One theory of formation of the outflow tract and vascular septation. LV, left ventricular; LVOT, left ventricular outflow tract; RV, right ventricle; RVOT, right ventricular outflow tract. Fig. 12. Diagram depicting the theory of ventricular outflow and great vessels’ septation by Bartlings et al. [9]. Numbers indicate specific aortic arch arteries. as nodal tissue, are slow conducting and resemble less developed primary myocardium, whereas other portions, such as ventricular conduction tissue, are fast conducting [30]. The embryological origin and formation of the sinus and atrioventricular nodal tissue is not clear. The ventricular conduction system formation is better known. The latter starts with the formation of an encircling ring of conducting myocardial tissue around the bulboventricular foramen. The dorsal portion of the ring will become the bundle of His. The portion of the ring covering the septum will become the left and right bundle branches. The anterior portion of the ring is called the septal branch and it disappears during normal embryological development. Other portions of this specialized tissue that form and later disappear are the right atrioventricular ring bundle and the retroartic branch. The right atrioventricular ring forms due to the rightward shift of the common atrioventricular valve, which originally connects the common atrium to the primitive R. Abdulla et al.: Cardiovascular Embryology Fig. 13. Degenerated aortic arch arteries (AAA) and the final great vessels anatomy. (left) ventricle. This results in a shift of the specialized myocardium rightward in a ring shape around the right atrioventricular orifice, only later to disappear. The retroarotic branch is formed as a result of the leftward shift of the outflow tract, causing some of the specialized conducting tissue to move and to be situated behind the aorta. Development of Pericardial Sac The right and left intracelomic cavities approach the midline as the two heart tubes are fusing into a medial tube (day 21). The two cavities approach each other and surround the heart tube. The ventral mesoderm is immediately absorbed and the two cavities communicate. The dorsal mesoderm persists until day 25. After the mesoderm is absorbed, the heart becomes suspended from the cranial and caudal ends. A band of connective tissue grows from the epicardium into the atrioventricular junction when the heart is four chambered, resulting in separation of atrial and ventricular myocardium. The bundle of His remains the only means of electrical conduction from atria to ventricles. The sinoatrial node, atrioventricular node, and the bundle of His receive sympathetic and parasympathetic nervous supply throughout the rest of gestation and even after birth to complete the development of the cardiac conduction system. Aortic Arches The first pair of aortic arches is formed by the curving of the ventral aorta to meet the dorsal aorta; these 199 will eventually contribute to the external carotid arteries (Fig. 13). The second pair of aortic arch arteries appears in week 4. These regress rapidly and only a portion remains, which forms the stapedial and hyoid arteries. The third pair of the aortic arch arteries appears at approximately the end of the fourth week; these will give rise to the common carotid arteries and the proximal portion of the internal carotid arteries. The distal portion of the internal carotid arteries is formed by the cranial portions of the dorsal aorta. The fourth aortic arch arteries develop soon after the third arch arteries. Their development differs on the left from that on the right. On the left side, they persist, connecting the ventral aorta to the dorsal aorta and forming the aortic arch. On the right, they form the proximal portion of the right subclavian artery. The fifth pair of aortic arch arteries is rudimentary and does not develop into any known vessels; this pair of aortic arch arteries is not seen in many embryo specimens. The sixth aortic arch arteries develop in the middle of the fifth week. The proximal portions develop into the main and right pulmonary arteries, whereas the distal portion of the left aortic arch artery develops into the ductus arteriosus (Fig. 13). References 1. Abdulla R (2000) The three dimensional heart. Pediatr Cardiol 21:409 2. Anderson RH (1986) Description of ventricular septal defect—or how long is a piece of string? Int J Cardiol 13:267–278 3. Anderson RH (1991) Simplifying the understanding of congenital malformation of the heart. Int J Cardiol 32:131–142 4. Anderson RH, Wilkinson JL, Rosenquist GC, Bergsma D (1978) The bulbus cordis. Morphogenesis and Malformation of the Cardiovascular System. Liss, New York, pp 1–28 5. Anderson PAW (1995) The molecular genetics of cardiovascular disease. Curr Opin Cardiol 10:33–43 6. Bartelings MM (1989) The outflow tract of the heart—embryologic and morphologic correlations. Int J Cardiol 22:289– 300 7. Bartelings MM (1990) The Outflow Tract of the Heart - embryologic and morphholayre correlations. Fnt. J Condcol 22: 289–300 8. Bartelings MM, Gittenberger-deGroot AC (1988) The arterial orifice level in the early human embryo. Anat Embryol 177: 537–542 9. Bartelings MM, et al. (1986) Contribution of the aortopulmonary septum to the muscular outlet septum in the human heart. Acta Morphol Neerl-Scand 24:181–192 10. Becker AE, Anderson RH (1984) Cardiac embryology. In: Nora JJ, Talao A (Eds.), Congenital Heart Disease: Causes and Processes. Futura, New York, pp 339–358 11. Benson DW, et al. (1996) New understanding in the genetics of congenital heart disease. Curr Opin Pediatr 8:505–511 12. Bockman ED (1987) Effect of neural crest on the development of the heart and arch arteries in the chick. Am J Anat 180:332– 341 200 13. Bolender D, Holterman ML (2001) Animated thoughts on teaching human development. FASEB J 15:Abstract 793.1 14. Burn J, Goodship J (1996) Developmental genetics of the heart. Curr Opin Genet Dev 6:322–326 15. Clark EB (1984) Hemodynamic control of the chick embryo cardiovascular system. In: Nora JJ, Talao A (Eds.), Congenital Heart Disease: Causes and Processes. Futura, New York, pp 337–386 16. Colvin EV (1998) Cardiac embryology. In: Garson A Jr (Eds.), The Science and Practice of Pediatric Cardiology, 2nd ed. Williams & Wilkins, Baltimore, pp 91–126 17. Creazzo TL, et al. (1998) Role of cardiac neural crest in cardiovascular development. Ann Rev Physiol 60:267–286 18. Hay DA (1978) Development and fusion of the endocardial cushion. In: Rosenquist GC, Bergsma D (Eds.), Morphogenesis and Malformations of the Cardiovascular System. Liss, New York, pp 69–90 19. Holterman MJ, et al (1999) The Visible Embryo Project: new approaches to embryology education [abstract]. American Academy of Pediatrics Surgical section meeting, Washington DC 20. Holterman MJ, Blew G, Bolender D, Abdulla R (2001) Clinical education development using multimedia [abstract]. Biannual meeting of the American Association of Clinical Anatomists and the British Association of Clinical Anatomists, Cambridge, UK 21. Holterman MJ, Oladapo A, Abdulla R (1999) Clinically relevant embryology: new approach to education [abstract]. American Academy of Pediatrics, Washington, DC 22. Kathiriya IS, Srivastava D (2000) Left–right asymmetry and cardiac looping: implications for cardiac development and congenital heart disease. Am J Med Genet 97:271–279 23. Kirby ML (1989) Plasticity and predetermination of mesencephalic and trunk neural crest transplanted into the region of the cardiac neural crest. Dev Biol 134:402–412 24. Kirby ML, Waldo KL (1990) Role of neural crest in congenital heart disease. Circulation 82:232–340 25. Larsen WJ (1997a) Human Embryology, 2nd edn. Churchill Livingstone, New York, pp 49–61 26. Larsen WJ (1997b) Human Embryology, 2nd edn. Churchill Livingstone, New York, pp 151–188 27. Los JA (1978) Cardiac septation and development of the aorta, pulmonary trunk, and pulmonary veins. In: Rosenquist GC, Bergsma D (Eds.), Morphogenesis and Malformations of the Cardiovascular System. Liss, New York, pp 109–138 28. McBride RE (1981) Development of the outflow tract and closure of the interventricular septum. Am J Anat 106:309–331 29. McGowan Jr FX (1992) Cardiovascular and airway interactions. Int Anesthesiol Clin 30:21–44 Pediatric Cardiology Vol. 25, No. 3, 2004 30. Moorman AF, de Jong F, Denyn MM, et al. (1998) Development of the cardiac conduction system. Circ Res 82:629– 644 31. Morse DE (1978) Scanning electron microscopy of the developing septa in the chick heart.. In: Rosenquist GC, Bergsma D (Eds.), Morphogenesis and Malformations of the Cardiovascular System. Liss, New York, pp 91–107 32. Nadal-Ginard B, Mahdavi V (1993) Basic mechanism of cardiac gene expression. Eur Heart J 14:suppl2–11 33. Nora J, Berg K, Nora AH (1991) Cardiovascular Disease: Genetics, Epidemiology and Prevention. Oxford University Press, New York 34. Olson EN, Srivastava D (1996) Molecular pathways controlling heart development. Science 272:671–675 35. Pensky B (1982) Review of Medical Embryology. McMillan, New York, pp 291–355 36. Pexieder T (1978) Development of the outflow tract of the embryonic heart. In: Rosenquist GC, Bergsma D (Eds.), Morphogenesis and Malformation of the Cardiovascular System. Liss, New York, pp 29–68 37. Pexieder T, Janecek P (1984) Organogenesis of the human embryonic and early fetal heart as studied by microdissection and SEM. In: Nora JJ, Talao A (Eds.), Congenital Heart Disease: Causes and Processes. Futura, New York, pp 401– 422 38. Srivastava D (2001) Genetic assembly of the heart: implications for congenital heart disease. Annu Rev Physiol 63:451– 469 39. Steding G, Seidl W (1984) Cardiac septation in normal development. In: Nora JJ, Talao A (Eds.), Congenital Heart Disease: Causes and Processes. Futura, New York, pp 481– 500 40. Thompson RP (1985) Morphogenesis of human cardiac outflow. Anat Rec 213:578–586 41. Van Mierop LHS (1979) Morphological development of the heart. In: Berne RM (Eds.), Handbook of Physiology, the Cardiovascular System. American Physiology Society, Bethesda, MD, pp 1–28 42. Van Mierop LHS (1986) Cardiovascular anomalies in DiGeorge syndrome and importance of neural crest as a possible pathogenetic factor. Am J Cardiol 58:133–137 43. Wenink ACG (1986) Embryology of the mitral valve. Int J Cardiol 11:75–84 44. Wenink ACG, Gittenberger-deGroot AC (1985) The role of atrioventricular endocardial cushion in the septation of the heart. Int J Cardiol 8:25–44 45. Whiten S, Smart SD, McLachlan JC, Aiton JF (1998) Computer-aided interactive three-dimensional reconstruction of the embryonic human heart. J Anat 193:337–345