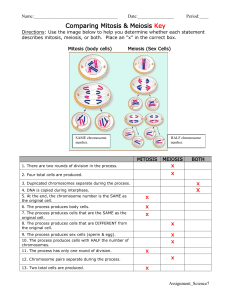
Chapter 7
advertisement

E S S E N T I A L Biology C A M P B E L L R E E C E CHAPTER 7 The Cellular Basis of Reproduction and Inheritance You began life as a single cell, but there are now more cells in your body than stars in the Milky Way. Overview: What Cell Reproduction Accomplishes 3 Passing On the Genes from Cell to Cell The Reproduction of Organisms Just in the past second, millions The Cell Cycle and Mitosis 4 Eukaryotic Chromosomes The Cell Cycle Mitosis and Cytokinesis Cancer Cells: Growing out of Control divided in two. Meiosis, the Basis of Sexual Reproduction 10 Homologous Chromosomes Gametes and the Life Cycle of a Sexual Organism The Process of Meiosis Review: Comparing Mitosis and Meiosis The Origins of Genetic Variation When Meiosis Goes Amok of your cells have The dance of the chromo- somes in a dividing cell is so precise that only one error occurs in 100,000 cell divisions. Each sperm or egg Evolution Link: New Species from Errors in Cell Division 21 produced in your reproductive organs carries one of over 8 million possible combinations of parental chromosomes. 2 C onsider the skin on your arm. The surface is a protective layer of dead cells, but underneath are layers of living cells busy carrying out the chemical reactions you studied in Unit One. The living cells are also engaged in another vital activity: They are reproducing themselves. The new cells are moving outward toward the skin’s surface, replacing dead cells that have rubbed off. This renewal of your skin goes on throughout your life. And when your skin is injured, additional cell reproduction helps heal the wound. In this chapter, you’ll learn what actually happens inside reproducing cells and how cell reproduction functions in the perpetuation of all life. Overview: What Cell Reproduction Accomplishes The replacement of lost or damaged cells is just one of the important roles that cell reproduction—or cell division, as we commonly call it—has played in your life. Another is growth. All of the trillions of cells in your body result from repeated cell divisions that began in your mother’s body with a single fertilized egg cell. (Figure 7.1 shows a human embryo at the eight-cell stage, after three rounds of cell division.) Figure 7.1 Human embryo at the eight-cell stage, the result of three rounds of cell division. The eight cells have identical sets of genes, as will the cells that they give rise to. As development continues, the ball of cells will transform itself into a baby and eventually into an adult. Passing On the Genes from Cell to Cell When a cell divides, the two “daughter” cells that result are ordinarily genetically identical to each other and to the original “parent” cell. (Biologists traditionally use the word daughter in this context; it does not imply gender.) Before the parent cell splits into two, it duplicates its chromosomes, the DNA-containing structures that carry the organism’s genes. Then, during the division process, the two sets of chromosomes are distributed to the daughter cells. As a rule, the daughter cells receive identical sets of chromosomes, with identical genes. Some organisms reproduce by simple cell division. Single-celled organisms such as amoebas reproduce this way, and the offspring are replicas of the parent (Figure 7.2). Because it does not involve fertilization of an egg by a sperm, this type of reproduction is called asexual reproLearn more about duction. Offspring produced by asexual reproduction inasexual and sexual herit all their chromosomes from a single parent. In addireproduction in tion to single-celled creatures, many multicellular Web/CD Activity 7A. organisms can also reproduce asexually. For example, some sea stars can divide into two pieces that regrow into two whole new individuals. And if you’ve ever grown an African violet from a clipping, you’ve observed asexual reproduction in plants. In asexual reproduction, there is one simple principle of inheritance: The parent and each of its offspring have identical genes. Sexual reproduction, which requires fertilization of an egg by a sperm, is different. The production of egg and sperm cells involves a special type of cell division, called meiosis, that occurs only in reproductive organs (such as testes and ovaries). As we’ll discuss later, a sperm or egg cell has only half as many chromosomes as the parent cell that gave rise to it. So two kinds of LM 300 The Reproduction of Organisms Figure 7.2 Asexual reproduction of an amoeba. This single-celled organism is reproducing by dividing in half. Its chromosomes have been duplicated, and the two identical sets of chromosomes have been allocated to opposite sides of the parent. When division is complete, the two daughter amoebas will be genetically identical to each other and to their parent. CHAPTER 7 The Cellular Basis of Reproduction and Inheritance 3 CheckPoint LM 600 cell division—meiosis and ordinary cell division—are involved in the lives of sexually reproducing organisms. The remainder of the chapter is divided into two main sections. The first section deals with the cell cycle and mitosis, the type of cell division responsible for asexual reproduction and for the growth and maintenance of multicellular organisms. The second section focuses on meiosis, the special type of cell division that produces sperm and eggs for sexual reproduction. Ordinary cell division produces two daughter cells that are genetically identical. Give three functions of this type of cell division. Answer: Cell replacement, growth of an organism, asexual reproduction of an organism Figure 7.3 A plant cell just before division. The colors result from staining. The purple threads are the chromosomes. (The thinner red threads in the surrounding cytoplasm are the cytoskeleton.) TEM 29,000× The Cell Cycle and Mitosis Almost all the genes of a eukaryotic cell—around 75,000 genes in humans —are located on chromosomes in the cell nucleus. (The main exceptions are genes on small DNA molecules found in mitochondria and chloroplasts.) As leading players in cell division, the chromosomes deserve a little more of our attention before we broaden our focus to the cell as a whole. Centromere Figure 7.4 Electron micrograph of a duplicated chromosome. The two identical copies of the chromosome are called sister chromatids. The constricted region is the centromere. The fuzzy appearance comes from the intricate twists and folds of the chromatin fibers. Sister chromatids Chromosome duplication Sister chromatids Figure 7.5 Chromosome dupliChromosome distribution to daughter cells 4 UNIT TWO cation and distribution. In the course of cell reproduction, the cell duplicates each chromosome and distributes the two copies to the daughter cells. This diagram focuses on a single chromosome. Genetics Eukaryotic Chromosomes Chromosomes get their name (Greek chroma, colored, and soma, body) from their attraction for certain stains used in microscopy. In the micrograph of a plant cell in Figure 7.3, each dark purple thread is an individual chromosome. Chromosomes are clearly visible under the light microscope as structures like these only when a cell is in the process of dividing. The rest of the time, the chromosomes exist as a diffuse mass of very long fibers that are too thin to distinguish in a light micrograph. The chromosomal material, called chromatin, is a combination of DNA and protein molecules. As a cell prepares to divide, its chromatin fibers coil up, forming compact, distinct chromosomes. Each eukaryotic chromosome contains one long DNA molecule, typically bearing thousands of genes. The attached protein molecules help organize the chromatin and help control the activity of its genes. The number of chromosomes in a eukaryotic cell, like the number of genes, depends on the species. For example, human body cells generally have 46 chromosomes. Well before a cell begins the division process, it duplicates all of its chromosomes. The DNA molecule of each chromosome is replicated, and new protein molecules attach as needed. The result is that each chromosome now consists of two copies called sister chromatids, which contain identical genes. Figure 7.4 is an electron micrograph of a human chromosome that has been duplicated. The two chromatids are joined together especially tightly at a region called the centromere. When the cell divides, the sister chromatids of a duplicated chromosome separate from each other, as shown in the simple diagram in Figure 7.5. Once separated from its sister, each chromatid is considered a fullfledged chromosome, and it is identical to the chromosome we started with. One of the new chromosomes goes to one daughter cell, and the other goes to the other daughter cell. In this way, each daughter cell receives a complete and identical set of chromosomes. A dividing human skin cell, for example, has 46 duplicated chromosomes, and each of the two daughter cells that results from it has 46 single chromosomes. Let’s now summarize how cell division fits into the life of an organism. Cell division is the basis of reproduction for every organism, and it enables a multicellular organism to grow from a single cell. It also replaces wornout or damaged cells, keeping the total cell number in a mature individual relatively constant. In your own body, for example, millions of cells must divide every second to maintain the total number of about 60 trillion cells. G1 The Cell Cycle How do chromosome duplication and cell division fit into the life of a cell? How often a cell divides depends on the role of the cell in the organism’s body. Some cells divide once a day, others less often, and highly specialized cells, such as mature muscle cells, not at all. Eukaryotic cells that do divide un- Cytokinesis dergo a cell cycle, an orderly sequence of events that extends from the time a cell first arises from cell division until it itself divides. As Figure 7.6 shows, most of the cell cycle is spent in interphase. This is a time when a cell metabolizes and performs its various other functions within the organism. For example, a cell in Mitosis your stomach lining might be making and releasing enzyme molecules that aid in digesting the food you eat. During interphase, a cell roughly doubles everything in its cytoplasm. It increases its supply of proteins, increases the number of many of its organelles (such as mitochondria and ribosomes), and grows in size. Typically, interphase lasts for at least 90% of the cell cycle. From the cell reproduction standpoint, the most important event of interphase is chromosome duplication, when the DNA in the nucleus is precisely doubled. This occurs approximately in the middle of interphase, and the period when it is occurring is called the S phase (for DNA synthesis). The interphase periods before and after the S phase are called the G1 and G2 phases, respectively (G stands for gap). During G2, each chromosome in the cell consists of two identical sister chromatids, and the cell is preparing to divide. The part of the cell cycle when the cell is actually dividing is called the mitotic phase (M phase). It includes two overlapping processes, mitosis and cytokinesis. In mitosis, the nucleus and its contents, notably the duplicated chromosomes, divide and are evenly distributed to form Test your knowledge two daughter nuclei. In cytokinesis, the cytoplasm is diof the cell cycle with vided in two. Cytokinesis usually begins before mitosis is Web/CD Activity 7B . completed. The combination of mitosis and cytokinesis produces two genetically identical daughter cells, each with a single nucleus, surrounding cytoplasm, and plasma membrane. Mitosis is a remarkably accurate mechanism for allocating identical copies of a large amount of genetic material to two daughter cells. Experiments with yeast cells, for example, indicate that an error in chromosome distribution occurs only once in about 100,000 cell divisions. Mitosis is unique to eukaryotes. Prokaryotes have only a single small chromosome (see Chapter 9) and use a simpler mechanism for allocating DNA to daughter cells. Mitotic phase (M) Interphase S (DNA synthesis— chromosome duplication) G2 Figure 7.6 The eukaryotic cell cycle. The cell cycle extends from the “birth” of a cell as a result of cell reproduction to the time the cell itself divides in two. The cell spends most of the cycle in interphase. A key event of interphase is the duplication of the chromosomes; the period during which this occurs is called the S phase (for DNA synthesis). Before S, the cell is said to be in the G1 phase; after S, the cell is in G2. The cell metabolizes and grows throughout interphase. The actual division process occurs during the mitotic phase (M phase), which includes mitosis (the division of the cell’s nucleus) and cytokinesis (the division of the cytoplasm). During interphase, the chromosomes are diffuse masses of thin fibers; they do not actually appear in the rodlike form you see here. CHAPTER 7 The Cellular Basis of Reproduction and Inheritance 5 Mitosis and Cytokinesis Figure 7.7 Cell reproduction: A dance of the chromosomes. After the chromatin doubles during interphase, the elaborately choreographed stages of mitosis—prophase, metaphase, anaphase, and telophase—distribute the duplicate sets of chromosomes to two separate nuclei. Cytokinesis then divides the cytoplasm, yielding two genetically identical daughter cells. The micrographs here show cells from a fish. The drawings include details not visible in the micrographs. For simplicity, only four chromosomes appear in the drawings. The light micrographs in Figure 7.7 show the cell cycle for an animal cell, with most of the figure devoted to the mitotic phase. With the onset of mitosis, striking changes are visible in the nucleus and other See mitosis animacellular structures. The text under the figure describes the tions and videos in events occurring at each stage. Mitosis is a continuum, Web/CD Activities 7C but biologists distinguish four main stages: prophase, and 7D. metaphase, anaphase, and telophase. The chromosomes are the stars of the mitotic drama, and their movements depend on the mitotic spindle, a football-shaped structure of microtubules that guides the separation of the two sets of daughter chromosomes. The spindle microtubules grow from two centrosomes, clouds of Prophase Interphase Centrosomes (with centriole pairs) Chromatin Early mitotic Centrosome Centromere spindle Fragments of nuclear envelope Nucleolus Nuclear envelope Plasma membrane Interphase Interphase is the period of cell growth, when the cell makes new molecules and organelles. At the point shown here, late interphase (G2), the cytoplasm contains two centrosomes. Within the nucleus, the chromosomes are duplicated, but they cannot be distinguished individually because they are still in the form of loosely packed chromatin fibers. The prominent nucleolus is an indication that the cell is making ribosomes. 6 UNIT TWO Genetics Chromosome, consisting of two sister chromatids Spindle microtubules Prophase During prophase, changes occur in both nucleus and cytoplasm. In the nucleus, the chromatin fibers coil, so that the chromosomes become thick enough to be seen with the light microscope. The nucleoli disappear. Each chromosome appears as two identical sister chromatids joined together, with a narrow “waist” at the centromere. In the cytoplasm, the mitotic spindle begins to form as microtubules grow out from the centrosomes, which are moving away from each other. Late in prophase, the nuclear envelope breaks up. The spindle microtubules can now reach the chromosomes, which are thick and have a protein structure (black dot) at their centromeres. Some of the spindle microtubules capture chromosomes by attaching to these structures, throwing the chromosomes into agitated motion. Other microtubules make contact with microtubules coming from the opposite spindle pole. The spindle moves the chromosomes toward the center of the cell. cytoplasmic material that in animal cells contain centrioles. (Centrioles are can-shaped structures made of microtubules; microtubules and centrioles were introduced in Chapter 3.) The role of centrioles in cell division is a mystery; destroying them experimentally does not interfere with normal spindle formation, and plant cells lack them entirely. Cytokinesis, the actual division of the cytoplasm into two cells, typically occurs during telophase. In animal cells, the cytokinesis process is known as cleavage. The first sign of cleavage is the appearance of a cleavage furrow, an indentation at the equator of the cell (Figure 7.8a). A ring of microfilaments in the cytoplasm just under the plasma membrane is responsible for the cleavage furrow. The ring contracts like the pulling of a drawstring, deepening the furrow and pinching the parent cell in two. Microfilaments are made of actin, a protein that also enables muscle cells to contract. Metaphase Anaphase Telophase and Cytokinesis Nucleolus forming Cleavage furrow Nuclear envelope forming Daughter chromosomes Spindle Metaphase Anaphase Telophase and Cytokinesis The mitotic spindle is now fully formed. The chromosomes convene on an imaginary plate equidistant from the two poles of the spindle. The centromeres of all the chromosomes are lined up at this plate. For each chromosome, the spindle microtubules attached to the two sister chromatids pull toward opposite poles. This tug of war keeps the chromosomes in the middle of the cell. Anaphase begins suddenly, when the sister chromatids of each chromosome separate. Each is now considered a fullfledged (daughter) chromosome. Motor proteins at the centromeres “walk” the daughter chromosomes along their microtubules toward opposite poles of the cell (see motor proteins in Figure 5.8). Meanwhile, these microtubules shorten. However, the microtubules not attached to chromosomes lengthen, pushing the poles farther apart and elongating the cell. Telophase begins when the two groups of chromosomes have reached the cell poles. Telophase is the reverse of prophase: Nuclear envelopes form, the chromosomes uncoil, nucleoli reappear, and the spindle disappears. Mitosis, the division of one nucleus into two genetically identical daughter nuclei, is now finished. Cytokinesis, the division of the cytoplasm, usually occurs with telophase. In animals, a cleavage furrow pinches the cell in two, producing two daughter cells. CHAPTER 7 The Cellular Basis of Reproduction and Inheritance 7 SEM 100× Cancer Cells: Growing out of Control Cleavage furrow For a plant or animal to grow and develop normally and maintain its tissues once full grown, it must be able to control the timing of cell division in different parts of its body. The sequential events of the cell cycle are directed by a cell cycle control system that consists of special proteins within the cell. When this control system malfunctions, cells may reproduce at the wrong time or in the wrong place. This can result in a benign tumor, an abnormal mass of essentially normal cells. Benign tumors can cause problems if they grow in certain organs, such as the brain, but usually they can be completely removed by surgery. They always remain at their original site in the body. Cleavage furrow Contracting ring of microfilaments What Is Cancer? Cancer, which currently claims the lives of one out of Daughter cells (a) Animal cell Cell plate forming Daughter nucleus LM 850× Wall of parent cell Cell wall Vesicles containing cell wall material Cytokinesis in a plant cell occurs differently (Figure 7.8b). A cell plate, a membranous disk containing cell-wall material, forms inside the cell and grows outward. Eventually the new piece of wall divides the cell in two. every five people in the United States and other developed nations, is a serious disease of the cell cycle. Unlike normal cells of the body, cancer cells have a severely deranged cell cycle control system; not only do they divide excessively, but they also exhibit other kinds of bizarre behavior. A lump resulting from the reproduction of a cancer cell is called a malignant tumor. The most dangerous attribute of cancer cells is their ability to spread into neighboring tissues and often to other parts of the body. Like a benign tumor, a malignant tumor displaces normal tissue as it grows (Figure 7.9). But if a malignant tumor is not killed or removed, it can spread into surrounding tissues. More alarming still, cells may split off from the tumor, invade the circulatory system (lymph vessels and blood vessels), and travel to new locations, where they can form new tumors. The spread of cancer cells beyond their original site is called metastasis. Cancers are named according to where they originate. They are grouped into four categories. Carcinomas are cancers that originate in the external or internal coverings of the body, such as the skin or the lining of the intestine. Sarcomas arise in tissues that support the body, such as bone and muscle. Cancers of blood-forming tissues, such as bone marrow and lymph nodes, are called leukemias and lymphomas. Cancer Treatment In addition to removing malignant tumors with surgery, physicians combat cancers in two other ways: radiation therapy and chemotherapy. Both types of cancer treatment attempt to stop cancer cells from dividing. In radiation therapy, parts of the body that have cancerous Cell plate New cell wall Figure 7.8 Cytokinesis in animal and plant cells. (a) In a dividing animal Daughter cells (b) Plant cell 8 UNIT TWO Genetics cell, a contracting ring of microfilaments pinches the cell in two. (b) In a plant cell, the development of a new piece of cell wall brings about cytokinesis. First, membrane-enclosed vesicles containing cell wall material collect at the middle of the cell. The vesicles gradually fuse, forming a membranous disk called the cell plate. The cell plate grows outward, accumulating more cell wall material as more vesicles join it. Eventually, the membrane of the cell plate fuses with the plasma membrane, and the cell plate’s contents join the parental cell wall. The result is two daughter cells, each bounded by its own continuous plasma membrane and a cell wall. CheckPoint tumors are exposed to high-energy radiation, which disrupts cell division. Because cancer cells divide more often than most normal cells, they are more likely to be dividing at any given time. So radiation can often destroy cancer cells without seriously injuring the Tumor normal cells of the body. However, there is sometimes enough damage to normal body cells to produce bad side effects. For example, Glandular damage to cells of the ovaries or testes can tissue cause sterility. Chemotherapy generally uses the same A tumor grows from a strategy as radiation; in this case, drugs that single cancer cell. disrupt cell division are administered to the patient. These drugs work in a variety of ways. Some, called antimitotic drugs, prevent cell division by interfering with the mitotic spindle. One antimitotic drug, vinblastine, prevents the spindle from forming in the first place; another, Taxol, freezes the spindle after it forms, keeping it from functioning. Vinblastine was first obtained from the periwinkle, a flowering plant native to tropical rain forests in Madagascar. Taxol is made from a chemical found in the bark of the Pacific yew, a tree found mainly in the northwestern United States. Taxol has fewer side effects than many anticancer drugs and seems to be effective against some hard-to-treat cancers of the ovary and breast. In the laboratory, researchers can grow cancer cells in culture. The cells are placed in a glass container, and nutrients are provided by an artificial liquid medium (Figure 7.10). Normal mammalian cells grow and divide in culture for only about 50 cell generations. But cancer cells are “immortal” —they can go on dividing indefinitely, as long as they have a supply of nutrients. It is by studying cancer cells in culture that researchers are learning about the molecular changes that make a cell cancerous. We will return to the topic of cancer in Chapter 10, after learning more about genes. 1. When in the cell cycle does each chromosome consist of two chromatids? 2. An organism called a plasmodial slime mold is one huge cytoplasmic mass with many nuclei. Explain how this “monster cell” could arise. Lymph vessels Metastasis Cancer cells invade neighboring tissue. Cancer cells spread through lymph and blood vessels to other parts of the body. Figure 7.9 Growth and metastasis of a malignant (cancerous) tumor of the breast. Figure 7.10 Growing cancer cells in the lab. This researcher is working under a fume hood to help prevent contamination of the cells by microbes from the air. 3. In what sense are the two daughter cells produced by mitosis identical? 4. When a cancer patient is treated with vinblastine, which prevents the mitotic spindle from forming, in what stage of mitosis are dividing cells trapped? Answers: 1. During G2, the last part of interphase, and during prophase and metaphase of the next mitosis 2. Mitosis occurs repeatedly without cytokinesis. 3. They have identical genes (DNA). 4. Prophase CHAPTER 7 The Cellular Basis of Reproduction and Inheritance 9 Meiosis,the Basis of Sexual Reproduction Figure 7.11 The varied products of sexual reproduction. Eddie Murphy, his wife Nicole, and their children pose for a family snapshot. Each child has inherited a unique combination of genes from the parents and displays a unique combination of traits. Pair of homologous chromosomes Centromere LM 3600 Sister chromatids Only maple trees produce more maple trees, only goldfish make more goldfish, and only people make more people. These simple facts of life have been recognized for thousands of years and are reflected in the age-old saying “Like begets like.” But in a strict sense, “Like begets like” applies only to asexual reproduction, such as the reproduction of the amoeba in Figure 7.2. In that case, because offspring inherit all their DNA from a single parent, they are exact genetic replicas of that one parent and of each other, and their appearances are very similar. The family photo in Figure 7.11 makes the point that in a sexually reproducing species, like does not exactly beget like. You probably resemble your parents more closely than you resemble a stranger, but you do not look exactly like your parents or your siblings. Each offspring of sexual reproduction inherits a unique combination of genes from its two parents, and this combined set of genes programs a unique combination of traits. As a result, sexual reproduction can produce great variation among offspring. Notice in the photograph that despite the family resemblances, each Murphy child has a unique appearance. You’ll find the same sort of similarities and differences in pictures of your own relatives. Long before anyone knew about genes or chromosomes, people recognized that individuals of sexually reproducing species are highly varied. What’s more, they learned to develop domestic breeds of plants and animals by controlling sexual reproduction. A domestic breed displays particular traits from among the great variety of traits found in the species as a whole. All dachshunds, for instance, have sausage-shaped bodies and short legs, whereas Saint Bernards are much taller and bulkier. The ancestry of dog breeds can be traced back for many generations, during which breeders reduced variability in the breed by mating only those dogs with specific traits. In a sense, selective breeding is an attempt to make like beget like more than it does in nature. Sexual reproduction depends on the cellular processes of meiosis and fertilization. But before discussing these processes, we return to chromosomes and their role in the life cycles of sexually reproducing organisms. Homologous Chromosomes Figure 7.12 Pairs of homologous chromosomes. To make this karyotype (chromosome display) of a man, a scientist broke open a cell in metaphase of mitosis, stained the released chromosomes with special dyes, made a micrograph, and then arranged the chromosome images in matching pairs. The result: 22 well-matched pairs (autosomes) and a twenty-third pair that consists of an X chromosome and a Y chromosome (sex chromosomes). Each chromosome consists of two sister chromatids closely attached all along their lengths. Notice that with the exception of X and Y, the homologous chromosomes of each pair match in size, centromere position, and staining pattern. 10 UNIT TWO Genetics If we examine a number of cells from any individual organism, we discover that virtually all of them have the same number and types of chromosomes. Likewise, if we examine cells from different individuals of a single species— sticking to one gender, for now—we find that they have the same number and types of chromosomes. Viewed with a microscope, your chromosomes would look just like those of Queen Elizabeth (if you’re a woman) or Michael Jordan (if you’re a man). A typical body cell, called a somatic cell, has 46 chromosomes in humans. If we break open a human cell in metaphase of mitosis, make a micrograph of the chromosomes, and arrange the chromosome images in an orderly array, we produce a display called a karyotype (Figure 7.12). Every (or almost every) duplicated chromosome has a twin that resembles it in size and shape. The two chromosomes of such a matching pair, called homologous chromosomes, carry the same sequence of genes controlling the same inherited characteristics. For example, if a gene influencing eye color is located at a particular place on one chromosome—for example, within the yellow band in the Figure 7.12 inset—then the homologous chromosome has a similar gene for eye color there. However, the two genes may be slightly different versions, unlike the ones on sister chromatids, which are identical. Altogether, we humans have 23 homologous pairs of chromosomes. Other species have different numbers of chromosomes, but these, too, usually match in pairs. For a human female, the 46 chromosomes fall neatly into 23 homologous pairs, with the members of each pair essentially identical in appearance. For a male, however, one pair of chromosomes do not look alike (see Figure 7.12). The nonmatching pair, called the sex chromosomes, determines the person’s gender. Like all mammals, human males have one X chromosome and one Y chromosome. Only small parts of the X and Y are homologous; most of the genes carried on the X chromosome do not have counterparts on the tiny Y, and the Y has genes lacking on the X. Females have two X chromosomes. The remaining chromosomes, found in both males and females, are called autosomes. For both autosomes and sex chromosomes, we inherit one chromosome of each pair from our mother and the other from our father. Haploid gametes (n = 23) n Egg cell n Sperm cell Fertilization Meiosis Diploid zygote (2n = 46) Multicellular diploid adults (2n = 46) 2n Gametes and the Life Cycle of a Sexual Organism Having two sets of chromosomes, one inherited from each parent, is a key Mitosis and factor in the human life cycle, outlined in Figure 7.13, and in the life cycles development of all other species that reproduce sexually. The life cycle of a multicellular organism is the sequence of stages leading from the adults of one generaFigure 7.13 The human life cycle. In each generation, the tion to the adults of the next. Let’s follow the human chromosomes through doubling of chromosome number that results from fertilization is offthe human life cycle. set by the halving of chromosome number that occurs in meiosis. Humans are said to be diploid organisms because almost all our cells are For humans, the number of chromosomes in a haploid cell (sperm or diploid: They contain two homologous sets of chromosomes. The total egg) is 23 (that is, n 23). The number of chromosomes in the number of chromosomes, 46 in humans, is the diploid diploid zygote and all somatic cells arising from it is 46 (2n 46). Review sexual life number (abbreviated 2n). The exceptions are the egg and cycles in Web/CD sperm cells, known as gametes. Made by meiosis in an Activity 7E. ovary or testis, each gamete has a single set of chromosomes: 22 autosomes plus a single sex chromosome, X or Y. A cell with a single chromosome set is called a haploid cell; it has only one member of each homologous pair. For humans, the haploid number (abbreviated n) is 23. In the human life cycle, sexual intercourse allows a haploid sperm cell from the father to reach and fuse with a haploid egg cell of the mother in the process known as fertilization. The resulting fertilized egg, called a zygote, is diploid. It has two homologous sets of 2 Homologous chromosomes, one set from each par1 Chromosomes chromosomes duplicate separate ent. The life cycle is completed as a sexually mature adult 3 Sister chromatids develops from the zyseparate gote. Mitotic cell division ensures that all Homologous Homologous Sister somatic cells of the pair of pair of chromatids human body receive chromosomes duplicated in diploid chromosomes copies of all of the zyparent cell gote’s 46 chromosomes. All sexual life cycles Interphase before meiosis Meiosis I Meiosis II involve an alternation of diploid and haploid homologous pair, packaging them in separate (haploid) daughFigure 7.14 How meiosis halves chromosome stages. Producing haploid number. This simplified diagram tracks just one pair of hoter cells. But each chromosome is still doubled. 3 Meiosis II gametes by meiosis keeps mologous chromosomes. 1 Each of the chromosomes is separates the sister chromatids. Each of the four daughter the chromosome number duplicated during the preceding interphase. 2 The first divicells is haploid and contains only one single chromosome from from doubling in every the homologous pair. sion, meiosis I, segregates the two chromosomes of the generation (Figure 7.14). CHAPTER 7 The Cellular Basis of Reproduction and Inheritance 11 The Process of Meiosis Meiosis, the process that produces haploid gametes in diploid organisms, resembles mitosis, but with two special features. The first is the halving of the number of chromosomes. In meiosis, a cell that has duplicated its chromosomes undergoes two consecutive divisions, called meiosis I and meiosis II. Because the two divisions of meiosis are preceded by only one Watch an animation duplication of the chromosomes, each of the four daughter of meiosis in Web/CD cells resulting from meiosis has only half as many chromoActivity 7F. somes as the starting cell—a haploid set of chromosomes. The chromosome number is actually haploid by the end of meiosis I, although there are still two sister chromatids per chromosome at that point. Figure 7.15 The stages of meiosis. The drawings here show the two cell divisions of meiosis, starting with a diploid animal cell containing four chromosomes. Each homologous pair consists of a red chromosome and a blue chromosome of the same size. The colors remind us that the members of a homologous pair were inherited from different parents and carry different versions of some genes. Meiosis I: Homologous chromosomes separate Interphase Centrosomes (with centriole pairs) Nuclear envelope Chromatin Like mitosis, meiosis is preceded by an interphase during which the chromosomes duplicate. Each chromosome then consists of two identical sister chromatids. Genetics Microtubules attached to chromosome Anaphase I Sister chromatids remain attached Spindle Interphase UNIT TWO Metaphase I Sites of crossing over Chromosomes duplicate 12 Prophase I Sister chromatids Tetrad Homologous chromosomes pair and exchange segments Centromere Tetrads line up Pairs of homologous chromosomes split up Meiosis I Prophase I Prophase I is the most complicated stage of meiosis. As the chromatin condenses, special proteins cause the homologous chromosomes to stick together in pairs. The resulting structure has four chromatids and is called a tetrad. Within each tetrad, chromatids of the homologous chromosomes exchange corresponding segments—they “cross over.” Because the versions of the genes on a chromosome (or one of its chromatids) may be different from those on its homologue, crossing over rearranges genetic information. As prophase I continues, the chromosomes condense further, a spindle forms, and the tetrads are moved toward the center of the cell. Metaphase I At metaphase I, the tetrads are aligned in the middle of the cell. The sister chromatids of each chromosome are still attached at their centromeres, where they are anchored to spindle microtubules. Notice that for each tetrad, the spindle microtubules attached to one homologous chromosome come from one pole of the cell, and the microtubules attached to the other chromosome come from the opposite pole. With this arrangement, the homologous chromosomes of each tetrad are poised to move toward opposite poles of the cell. Anaphase I As in anaphase of mitosis, chromosomes now migrate toward the poles of the cell. But in contrast to mitosis, the sister chromatids migrate as a pair instead of splitting up. They are separated not from each other, but from their homologous partners. So in the drawing, you see two still-doubled chromosomes moving toward each pole. The second special feature of meiosis is an exchange of genetic material —pieces of chromosomes—between homologous chromosomes. This exchange, called crossing over, occurs during the first prophase of meiosis. We’ll look more closely at crossing over later. For now, study Figure 7.15 and the text below it, which describe the stages of meiosis in detail. As you go through Figure 7.15, keep in mind the difference between homologous chromosomes and sister chromatids: The two chromosomes of a homologous pair are individual chromosomes that were inherited from different parents. Homologues appear alike in the microscope, but they have different versions of some of their genes (for example, a gene for freckles on one chromosome and a gene for the absence of freckles at the same place on the homologue). The homologues in Figure 7.15 (and later figures) are colored red and blue to remind you that they differ in this way. In the interphase just before meiosis, each homologue replicates to form sister chromatids that remain together until anaphase of meiosis II. Before crossing over occurs, sister chromatids are identical and carry the same versions of all their genes. Meiosis II: Sister chromatids separate Telophase I and Cytokinesis Prophase II Metaphase II Telophase II and Cytokinesis Anaphase II Cleavage furrow Sister chromatids separate Two haploid cells form; chromosomes are still double Telophase I and Cytokinesis In telophase I, the chromosomes arrive at the poles of the cell. When they finish their journey, each pole has a haploid chromosome set, although each chromosome is still in duplicate form. Usually, cytokinesis occurs along with telophase I, and two haploid daughter cells are formed. Depending on the species, the nuclei may or may not return to an interphase state. But in either case, there is no further chromosome duplication. Haploid daughter cells forming During another round of cell division, the sister chromatids finally separate; four haploid daughter cells result, containing single chromosomes Meiosis II Meiosis II is essentially the same as mitosis. The important difference is that meiosis II starts with a haploid cell. During prophase II, a spindle forms and moves the chromosomes toward the middle of the cell. During metaphase II, the chromosomes are aligned as they are in mitosis, with the microtubules attached to the sister chromatids of each chromosome coming from opposite poles. In anaphase II, the centromeres of sister chromatids finally separate, and the sister chromatids of each pair, now individual daughter chromosomes, move toward opposite poles of the cell. In telophase II, nuclei form at the cell poles, and cytokinesis occurs at the same time. There are now four daughter cells, each with the haploid number of single chromosomes. CHAPTER 7 The Cellular Basis of Reproduction and Inheritance 13 Mitosis Meiosis Parent cell (before chromosome duplication) Duplicated chromosome (two sister chromatids) Chromosome duplication Chromosome duplication Site of crossing over Meiosis I Tetrad formed by pairing of homologous chromosomes Pairing of homologous chromosomes Prophase Prophase I 2n = 4 Chromosomes align at the middle of the cell Metaphase Anaphase Telophase Sister chromatids separate during anaphase 2n Daughter cells of mitosis 2n Tetrads align at the middle of the cell Metaphase I Homologous chromosomes separate during anaphase I; sister chromatids remain together Anaphase I Telophase I Daughter cells of meiosis I Haploid n=2 No further chromosomal duplication; sister chromatids separate during anaphase II Meiosis II n n n Daughter cells of meiosis II Figure 7.16 Comparing mitosis and meiosis. The events unique to meiosis occur during meiosis I: In prophase I, duplicated homologous chromosomes pair to form tetrads, and crossing over occurs between homologous (nonsister) chromatids. In metaphase I, tetrads (rather than 14 UNIT TWO Genetics individual chromosomes) are aligned at the center of the cell. During anaphase I, sister chromatids of each chromosome stay together and go to the same pole of the cell as homologous chromosomes separate. At the end of meiosis I, there are two haploid cells, but each chromosome still has two sister n chromatids.Meiosis II is virtually identical to mitosis and separates sister chromatids. But unlike mitosis, meiosis II yields daughter cells with a haploid set of chromosomes. Review: Comparing Mitosis and Meiosis We have now described the two ways that cells of eukaryotic organisms divide. Mitosis, which provides for growth, tissue repair, and asexual reproduction, produces daughter cells genetically identical to the parent cell. Meiosis, needed for sexual reproduction, yields haploid daughter cells— cells with only one member of each homologous chromosome pair. For both mitosis and meiosis, the chromosomes duplicate only once, in the preceding interphase. Mitosis involves one division of the nucleus, and it is usually accompanied by cytokinesis, producing two diploid cells. Meiosis entails two nuclear and cytoplasmic divisions, yielding four haploid cells. Figure 7.16 (at the left) compares mitosis and meiosis, tracing these two processes for a diploid parent cell with four chromosomes. As before, homologous chromosomes are those matching in size. Notice that all the events unique to meiosis occur during meiosis I. The Origins of Genetic Variation As we discussed earlier, offspring that result from sexual reproduction are genetically different from their parents and from one another. When we discuss natural selection and evolution in Unit Three, we will see that this genetic variety in offspring is the raw material for natural selection. For now, let’s take another look at meiosis and fertilization to see how genetic variety arises. Possibility 1 Possibility 2 Metaphase of meiosis I Independent Assortment of Chromosomes Figure 7.17 illustrates one way in which meiosis contributes to genetic variety. The figure shows how the arrangement of homologous chromosome pairs at metaphase of meiosis I affects the resulting gametes. Once again, our example is from an organism with a diploid chromosome number of 4, with red and blue used to differentiate homologous chromosomes. These colors highlight the fact that homologous chromosomes differ genetically, although the two look alike under a microscope. (For example, the chromosome 3 you inherited from your mother undoubtedly carries many genes that are slightly different from those on the homologous chromosome 3 you received from your father.) Combination 1 The orientation of the homologous pairs of chromosomes (tetrads) at metaphase I is a matter of chance, like the flip of a coin. In this example, there are two possible ways that the two tetrads can align during metaphase I. In possibility 1, the tetrads are oriented with both red chromosomes on the same side. In this case, each of the gametes produced at the end of meiosis II has only red or only blue chromosomes (combinations 1 and 2). In possibility 2, the tetrads are oriented differently. This arrangement produces gametes that each have one red and one blue chromosome. Furthermore, half the gametes have a big blue chromosome and a small red one (combination 3), and half have a big red one and a small blue one (combination 4). So we see that for this example, a total of four chromosome combinations is possible in the gametes, and the organism will in fact produce gametes of all four types. This variety in gametes arises because each homologous pair Metaphase of meiosis II Gametes Combination 2 Combination 3 Combination 4 Figure 7.17 Results of alternative arrangements of chromosomes at metaphase of meiosis I. In this figure, we consider the consequences of meiosis in a diploid organism with four chromosomes (two homologous pairs). The positioning of each homologous pair of chromosomes (tetrad) at metaphase of meiosis I is random; the two red chromosomes can be on the same side (possibility 1) or on opposite sides (possibility 2). The arrangement of chromosomes at metaphase I determines which chromosomes will be packaged together in the haploid gametes. Because possibilities 1 and 2 are equally likely, the four possible types of gametes will be made in approximately equal numbers. CHAPTER 7 The Cellular Basis of Reproduction and Inheritance 15 Tetrad Prophase I of meiosis Chiasma, site of crossing over Metaphase I Spindle microtubules Metaphase II Gametes of chromosomes orients itself at metaphase I independently of the other pair. For a species with more than two pairs of chromosomes, such as the human, every chromosome pair orients independently of all the others at metaphase I. (Chromosomes X and Y behave as a homologous pair in meiosis.) For any species, the total number of chromosome combinations that can appear in gametes is 2n, where n is the haploid number. For the organism in this figure, n 2, so the number of chromosome combinations is 22, or 4. For a human (n 23), there are 223, or about 8 million, possible chromosome combinations! This means that every gamete a human produces contains one of about 8 million possible combinations of maternal and paternal chromosomes. Random Fertilization How many possibilities are there when a gamete from one individual unites with a gamete from another individual during fertilization? A human egg cell, representing one of about 8 million possibilities, is fertilized at random by one sperm cell, representing one of about 8 million other possibilities. By multiplying 8 million by 8 million, we find that a man and a woman can produce a diploid zygote with any of 64 trillion combinations of chromosomes! So we see that the random nature of fertilization adds a huge amount of potential variability to the offspring of sexual reproduction. These large numbers suggest that independent orientation of chromosomes at metaphase I and random fertilization could account for all the variety we see among people. Actually, these two events are only part of the picture, as we see next. Crossing Over So far, we have focused on genetic variability in gametes and Recombinant chromosomes Figure 7.18 The results of crossing over during meiosis. This diagram focuses on a single pair of homologous chromosomes (a tetrad). Early in prophase I of meiosis, homologous (nonsister) chromatids exchange corresponding segments, remaining attached at the crossover points. Sister chromatids are joined at their centromeres. Following these chromosomes through the rest of meiosis, we see that crossing over gives rise to recombinant chromosomes—individual chromosomes that combine genetic information originally derived from different parents. With multiple pairs of homologous chromosomes, the result is a huge variety of gametes. 16 UNIT TWO Genetics zygotes at the whole-chromosome level. We have ignored crossing over, the exchange of corresponding segments between two homologous chromosomes, which occurs during prophase I of meiosis. Figure 7.18 shows crossing over between two homologous chromosomes and the results in the gametes. At the time that crossing over begins, homologous chromosomes are closely paired all along their lengths, with a precise gene-by-gene alignment. The sites of crossing over appear as X-shaped regions; each is called a chiasma (Greek for cross; plural, chiasmata.) The homologous chromatids remain attached to each other at chiasmata until anaphase I. The exchange of segments by homologous chromatids adds to the genetic variety that results from the independent orientation of chromosome pairs at metaphase I. In Figure 7.18, if there were no crossing over, meiosis could produce only two types of gametes. These would be the ones ending up with the “parental” types of chromosomes, either all blue or all red (as in Figure 7.17). With crossing over, gametes arise that have chromosomes that are part red and part blue. These chromosomes are called “recombinant” because they result from genetic recombination, the production of gene combinations different from those carried by the parental chromosomes. Because most chromosomes contain thousands of genes, a single crossover event can affect many genes. When we also consider that multiple crossovers can occur in each tetrad, it’s no wonder that gametes and the offspring that result from them can be so varied. In fact, it’s surprising that even siblings resemble one another as much as they do. We have now examined three sources of genetic variability in sexually reproducing organisms: crossing over during prophase I of meiosis, independent orientation of chromosome pairs at metaphase I, and random fer- tilization. When we take up molecular genetics in Chapter 9, we will see yet another source of variation—mutations, which are rare changes in the DNA of genes. The different versions of genes found on homologous chromosomes originally arose from mutations, and it is mutations that are ultimately responsible for the genetic diversity in living organisms. To help you understand the origins of genetic variability, go to Web/CD Activity 7G. When Meiosis Goes Amok Down Syndrome: An Extra Chromosome 21 Figure 7.12 showed a normal human complement of 23 pairs of chromosomes. Figure 7.19 is different; besides having two X chromosomes (because it’s from a female), it has three number 21 chromosomes. This condition is called trisomy 21. In most cases, a human embryo with an abnormal number of chromosomes is spontaneously aborted (miscarried) long before birth. However, some aberrations in chromosome number, including trisomy 21, seem to upset the genetic balance less drastically, and individuals carrying them survive. These people usually have a characteristic set of symptoms, called a syndrome. A person with trisomy 21, for instance, is said to have Down syndrome (named after John Langdon Down, who described it in 1866). Trisomy 21 is the most common chromosome number abnormality. Affecting about one out of every 700 children born, it is the most common serious birth defect in the United States. Chromosome 21 is one of our smallest chromosomes, but an extra copy produces a number of effects. Down syndrome includes characteristic facial features—frequently a fold of skin at the inner corner of the eye (epicanthic fold), a round face, flattened nose bridge, and small, irregular teeth—as well as short stature, heart defects, and susceptibility to respiratory infection, leukemia, and Alzheimer’s disease. People with Down syndrome usually have a life span shorter than normal. They also exhibit varying degrees of mental retardation. However, individuals with the syndrome may live to middle age or beyond, and many are socially adept and able to hold a job. A few women with Down syndrome have had children, though most people with the syndrome are sexually underdeveloped and sterile. Half the eggs produced by a woman with Down syndrome will have an extra chromosome 21, so there is a 50% chance that she will transmit the syndrome to her child. As indicated in Figure 7.20, the incidence of Down syndrome in the offspring of normal parents increases markedly with the age of the mother. Down syndrome strikes less than 0.05% of children (fewer than one in 2000) born to women under age 30. The risk climbs to 1% for mothers in their late 30s and is even higher for older mothers. Because of this relatively high risk, pregnant women over 35 are candidates for fetal testing for trisomy 21 and other chromosomal abnormalities (see Chapter 8). What causes trisomy 21? We address that question next. Figure 7.19 Trisomy 21 and Down syndrome. The karyotype (left) shows trisomy 21; notice the three copies of chromosome 21. The child displays the characteristic facial features of Down syndrome. Infants with Down syndrome (per 1000 births) So far, our discussion of meiosis has focused on the process as it normally and correctly occurs. But what happens when an error occurs in the process? 90 80 70 60 50 40 30 20 10 0 20 25 30 35 40 Age of mother 45 50 Figure 7.20 Maternal age and Down syndrome. The chance of having a baby with Down syndrome rises with the age of the mother. CHAPTER 7 The Cellular Basis of Reproduction and Inheritance 17 (a) (b) Meiosis I How Accidents During Meiosis Can Alter Chromosome Number Meiosis occurs repeatedly in our lifetime as our testes or ovaries produce gametes. Almost always, the meiotic spindle distributes chromosomes to daughter cells withNondisjunction out error. But occasionally there is an accident, called a nondisjunction, in which the members of Meiosis II a chromosome pair fail to separate at anaphase. Nondisjunction can occur in meiosis I or II (Figure 7.21). In either case, gametes with abnormal numbers of chromosomes result. Nondisjunction Figure 7.22 shows what can happen when an abnormal gamete produced by nondisjunction Gametes unites with a normal gamete in fertilization. When a normal sperm fertilizes an egg cell with an extra chromosome, the result is a zygote with a total of 2n 1 chromosomes. Mitosis then transn+1 n+1 n–1 n–1 n+1 n–1 n n mits the abnormality to all embryonic cells. If the Number of chromosomes organism survives, it will have an abnormal karyFigure 7.21 Two types of nondisjunction. In both parts of otype and probably a syndrome of disorders the figure, the cell at the top is diploid (2n), with two pairs of homolocaused by the abnormal number of genes. gous chromosomes. (a) A pair of homologous chromosomes fails to Nondisjunction can lead to an abnormal chromosome number in either separate during anaphase of meiosis I, even though the rest of sex of any sexually reproducing, diploid organism, including humans. If, for meiosis occurs normally. In this case, all the resulting gametes end example, there is nondisjunction affecting human chromosome 21 during up with abnormal numbers of chromosomes. (b) Meiosis I is normal, meiosis I, half the resulting gametes will carry an extra chromosome 21. but a pair of sister chromatids fail to move apart in one of the cells Then if one of these gametes unites with a normal gamete in fertilization, during anaphase of meiosis II. In this case, two gametes have the trisomy 21 will result. normal complement of two chromosomes each, but the other two Nondisjunction explains how abnormal chromosome numbers come gametes are abnormal. about, but what causes nondisjunction in the first place? We do not know the answer, nor do we fully understand why offspring with trisomy 21 are more likely to be born as a woman ages. We do know, however, that meiosis begins in a woman’s ovaries before she is born but is not completed until years later, at the time of an ovulation. Because only one egg cell usually matures each month, a cell might remain arrested in the middle of meiosis Egg for decades. Perhaps damage to the cell during this time leads to meiotic ercell rors. It seems that the longer the time lag, the greater the chance that there will be errors such as nondisjunction when meiosis is completed. n+1 Sperm cell n (normal) Zygote 2n + 1 Figure 7.22 Fertilization after nondisjunction in the mother. Assuming that the organism has a diploid number of 4 (2n 4), the sperm is a normal haploid cell (n 2). The egg cell, however, contains an extra copy of the larger chromosome as a result of nondisjunction in meiosis; it has a total of n 1 3 chromosomes. When the sperm and egg fuse during fertilization, the result is an abnormal zygote with an extra chromosome; it has 2n 1 5 chromosomes. 18 UNIT TWO Genetics Abnormal Numbers of Sex Chromosomes Nondisjunction in meiosis does not affect just autosomes, such as chromosome 21. It can also lead to abnormal numbers of sex chromosomes (X and Y). Unusual numbers of sex chromosomes seem to upset the genetic balance less than unusual numbers of autosomes. This may be because the Y chromosome is very small and carries fewer genes than other chromosomes. Also, most of the genes on the Y chromosome affect maleness but not functions that are essential to the person’s survival. A peculiarity of X chromosomes in humans and other mammals also helps an individual tolerate unusual numbers of X chromosomes: In mammals, the cells usually operate with only one functioning X chromosome because extra copies of the chromosome become inactivated in each cell (see Chapter 10). Table 7.1 lists the most common sex chromosome abnormalities. An extra X chromosome in a male, making him XXY, occurs approximately once Table 7.1 Sex Chromosomes Abnormalities of Sex Chromosome Number in Humans Syndrome Origins of Nondisjunction Frequency in Population XXY Klinefelter syndrome (male) Meiosis in egg or sperm formation 1 2000 XYY Normal male Meiosis in sperm formation 1 2000 XXX Metafemale Meiosis in egg or sperm formation 1 1000 XO Turner syndrome (female) Meiosis in egg or sperm formation 1 5000 in every 2000 live births (once in every 1000 male births). This disorder is called Klinefelter syndrome. Men with this disorder have male sex organs, but the testes are abnormally small and the individual is sterile. The syndrome often includes breast enlargement and other feminine body contours (Figure 7.23, left). The person is usually of normal intelligence. Klinefelter syndrome is also found in individuals with more than one additional sex chromosome, such as XXYY, XXXY, or XXXXY. These abnormal numbers of sex chromosomes probably result from multiple nondisjunctions. Such men are more likely to be mentally retarded than XY or XXY individuals. Human males with a single extra Y chromosome (XYY) do not have any well-defined syndrome, although they tend to be taller than average. Females with an extra X chromosome (XXX) are called metafemales. They have limited fertility but are otherwise apparently normal. Females who are lacking an X chromosome are designated XO; the O simply indicates the absence of a second sex chromosome. These women have Turner syndrome. They have a characteristic appearance, including short stature and often a web of skin extending between the neck and shoulders (Figure 7.23, right). Women with Turner syndrome are sterile because their sex organs do not fully mature at adolescence, and they have poor development of breasts and other secondary sex characteristics. However, they are usually of normal intelligence. The XO condition occurs in about one in 5000 babies born (about one in 2500 female births). The sex chromosome abnormalities described here illustrate the crucial role of the Y chromosome in determining a person’s sex. In general, a single Y chromosome is enough to produce “maleness,” even when it is combined with several X chromosomes. The absence of a Y chromosome results in “femaleness.” Breast development Poor beard growth Web of skin Constriction of aorta Poor breast development Underdeveloped testes A man with Klinefelter syndrome (XXY) Underdeveloped ovaries A woman with Turner syndrome (XO) Figure 7.23 Syndromes associated with unusual numbers of sex chromosomes. Alterations of Chromosome Structure Even if all chromosomes are pres- ent in normal numbers, abnormalities in chromosome structure may cause disorders. Breakage of a chromosome can lead to a variety of rearrangements CHAPTER 7 The Cellular Basis of Reproduction and Inheritance 19 Deletion (a) Duplication Homologous chromosomes (b) Inversion (c) Reciprocal translocation Nonhomologous chromosomes (d) Figure 7.24 Alterations in chromosome structure. (a) A deletion is the removal of a chromosome segment. (b) A duplication is the repetition of a segment. When a duplication results from the movement of a chromosomal segment to a homologous chromosome, as shown here, the “duplicates” may be slightly different. (c) An inversion is the reversal of a segment within a chromosome. (d) A translocation is the movement of a segment to a nonhomologous chromosome. In the case shown here, the two nonhomologous chromosomes trade segments, producing a reciprocal translocation. affecting the genes of that chromosome. Figure 7.24 shows four types of rearrangement. If a fragment of a chromosome is lost, the remaining chromosome has a deletion. If a fragment from one chromosome is inserted into a homologous chromosome, it produces a duplication there. If a fragment reattaches to the original chromosome but in the reverse direction, an inversion results. Inversions are less likely than deletions or duplications to produce harmful effects, because in inversions, all genes are still present in their normal number. Deletions, especially large ones, tend to have the most serious effects. One example in humans is a specific deletion in chromosome 5 that causes the cri du chat (“cat-cry”) syndrome. A child born with this syndrome is mentally retarded and has a small head and a cry like the mewing of a cat. Death usually occurs in infancy or early childhood. Another type of chromosome change is chromosomal translocation, the attachment of a chromosome fragment to a nonhomologous chromosome. Figure 7.24d shows a translocation that is reciprocal; that is, two nonhomologous chromosomes exchange segments. Like inversions, translocations may or may not be harmful. Some people with Down syndrome have only part of a third chromosome 21; as the result of a translocation, it is attached to another (nonhomologous) chromosome. Whereas chromosome alterations in sperm or egg can cause congenital disorders, such changes in a somatic cell may contribute to the development of cancer. For example, a chromosomal translocation in somatic cells in the bone marrow is associated with chronic myelogenous leukemia (CML). CML is the most common of the leukemias, the cancers affecting cells that give rise to white blood cells (leukocytes). In the cancerous cells of most CML patients, a part of chromosome 22 has switched places with a small fragment from chromosome 9 (Figure 7.25). This reciprocal translocation activates a gene that leads to leukemia. The chromosome ending up with the activated cancer-causing gene is called the “Philadelphia chromosome,” after the city where it was discovered. Chromosome 9 Reciprocal translocation Chromosome 9 Activated cancer-causing gene Chromosome 22 “Philadelphia chromosome” Figure 7.25 The translocation associated with a type of leukemia. This is a reciprocal translocation in which one end of chromosome 9 trades places with about half of chromosome 22. One of the genes moved from chromosome 9 is activated to become a cancer-causing gene in its new home, called a “Philadelphia chromosome.” When a Philadelphia chromosome arises in a bone marrow cell that is destined to develop into a white blood cell (leukocyte), the result is a type of leukemia called chronic myelogenous leukemia (CML). 20 UNIT TWO Genetics CheckPoint 1. is to somatic cells as haploid is to . 2. If a diploid cell with 18 chromosomes undergoes meiosis, the resulting gametes will each have chromosomes. 3. Explain how mitosis conserves chromosome number while meiosis reduces the number in half. 4. In what important way is anaphase of meiosis II like anaphase of mitosis? 5. Name two events during meiosis that contribute to genetic variety among gametes. 6. How does the karyotype of a human female differ from that of a male? 7. What is the chromosomal basis of Down syndrome? 8. Explain how nondisjunction in meiosis could result in a diploid gamete. 9. How is reciprocal translocation different from normal crossing over? Answers: 1. Diploid; gametes. 2. 9 3. In mitosis, a single replication of the chromosomes is followed by one division of the cell. In meiosis, a single replication of the chromosomes is followed by two cell divisions. 4. Sister chromatids separate. 5. Crossing over between homologous chromosomes during prophase I and independent orientation of tetrads at metaphase I 6. A female has two X chromosomes; a male has an X and a Y. 7. Three copies of chromosome 21 (trisomy 21) 8. A diploid gamete would result if there were nondisjunction of all the chromosomes during meiosis I or II. 9. Normal crossing over is an exchange of segments between homologous chromosomes; reciprocal translocation occurs between nonhomologous chromosomes. Evolution Link: New Species from Errors in Cell Division Errors in meiosis or mitosis do not always lead to problems. In fact, biologists believe that such errors have been instrumental in the evolution of many species. Numerous plant species, in particular, seem to have originated from accidents during cell division that resulted in extra sets of chromosomes. The new species is polyploid, meaning that it has more than two sets of homologous chromosomes in each somatic cell. At least half of all species of flowering plants are polyploid, including such useful ones as wheat, potatoes, apples, and cotton. Let’s consider one scenario by which a diploid (2n) plant species might generate a tetraploid (4n) plant. Imagine that, like many plants, our diploid plant produces both sperm and egg cells and can selfLearn more about fertilize. If meiosis fails to occur in the plant’s reproductive new species resulting organs and gametes are instead produced by mitosis, the from errors in cell digametes will be diploid. The union of a diploid (2n) sperm vision in the Web/CD with a diploid (2n) egg in self-fertilization will produce a Evolution Link and The Process of Science. tetraploid (4n) zygote, which may develop into a mature tetraploid plant that can itself reproduce by self-fertilization. The tetraploid plants will constitute a new species, one that has evolved in just one generation. Although polyploid animal species are less common than polyploid plants, they are known to occur among the fishes and amphibians. Recently, researchers in Chile have identified the first candidate for polyploidy among CHAPTER 7 The Cellular Basis of Reproduction and Inheritance 21 the mammals, a rat whose cells seem to be tetraploid (Figure 7.26). Tetraploid organisms are sometimes strikingly different from their recent diploid ancestors—larger, for example. Scientists don’t yet understand exactly how polyploidy brings about such differences. You’ll learn more about the evolution of polyploid species in Chapter 13. In Chapter 8, we continue our study of genetic principles by looking at the rules governing the inheritance of biological traits and the connection between these traits and the organism’s chromosomes. Figure 7.26 Chock full of chromosomes+a tetraploid mammal? The somatic cells of this red viscacha rat from Argentina have about twice as many chromosomes as those of closely related species. (Interestingly, the heads of its sperm are unusually large, presumably a necessity for holding all that genetic material.) Scientists think that this rat is a tetraploid species that arose when an ancestor somehow doubled its chromosome number, probably by errors in mitosis or meiosis within the animal’s reproductive organs. Researchers are studying the rat’s chromosomes to verify that it actually has four homologous sets. Chapter Review Summary of Key Concepts Overview: What Cell Reproduction Accomplishes Cell reproduction, usually called cell division, enables a multicellular organism to grow and develop and to replace damaged or lost cells. Passing On the Genes from Cell to Cell Most cell division involves a duplication of all the chromosomes, followed by the distribution of the two identical sets of chromosomes to two “daughter” cells when the cell divides in two. The daughter cells are genetically identical. ● The Reproduction of Organisms Some organisms use ordinary cell division to reproduce. Their offspring are therefore genetically identical to the one parent and to each other. Organisms that reproduce sexually, by the union of a sperm with an egg cell, carry out another type of cell division in their reproductive organs. This process, meiosis, yields sperm and egg cells with only half as many chromosomes as ordinary body cells. Web/CD Activity 7A Asexual and Sexual Reproduction ● ● The Cell Cycle and Mitosis ● Eukaryotic Chromosomes The many genes of a eukaryotic cell are grouped into multiple chromosomes in the nucleus. Each chromosome contains a very long DNA molecule with thousands of genes. Individual chromosomes are visible only when the cell is in the process of dividing; otherwise, they are in the form of thin, loosely packed chromatin fibers. Before a cell starts dividing, the chromosomes duplicate, producing sister chromatids (containing identical DNA) joined together at the centromere. Cell division involves the separation of sister chromatids and results in two daughter cells, each containing a complete and identical set of chromosomes. ● The Cell Cycle Cell division is only one phase, called the mitotic phase, of the eukaryotic cell cycle. Most of the cycle is spent in interphase, when meta- 22 UNIT TWO Genetics bolic activity is high, chromosomes duplicate, many cell parts are made, and the cell grows in size. Eukaryotic cell division consists of two processes: mitosis and cytokinesis. Mitosis is the process that distributes the duplicated chromosomes to daughter nuclei; cytokinesis is the division of the cytoplasm to create two daughter cells. Web/CD Activity 7B The Cell Cycle ● Mitosis and Cytokinesis At the start of mitosis, the chromosomes coil up, becoming thick enough to be visible with a light microscope. The nuclear envelope breaks down, and a mitotic spindle made of microtubules moves the chromosomes to the middle of the cell. The sister chromatids then separate and are moved to opposite poles of the cell, where two new nuclei form. Cytokinesis overlaps the end of mitosis. Mitosis and cytokinesis produce genetically identical cells. In animals, cytokinesis occurs by cleavage, which pinches the cell apart. In plants, a membranous cell plate splits the cell in two. Web/CD Activity 7C Mitosis and Cytokinesis Animation Web/CD Activity 7D Mitosis and Cytokinesis Video ● ● ● ● Cancer Cells: Growing out of Control When the cell cycle control system malfunctions, a cell may divide excessively and form a tumor. Cancer cells have highly abnormal cell cycles. They can grow to form malignant tumors, invade other tissues (metastasize), and even kill the organism. Radiation and chemotherapy are effective as treatments because they interfere with cell division. Meiosis, the Basis of Sexual Reproduction Homologous Chromosomes The somatic cells (body cells) of each species contain a specific number of chromosomes; for example, human cells have 46, making up 23 pairs (two sets) of homologous chromosomes. The chromosomes of a homologous pair carry genes for the same characteristics at the same places. In mammalian males, one pair of chromosomes are only partially homologous: the sex chromosomes X and Y. Females have two X chromosomes. ● Gametes and the Life Cycle of a Sexual Organism Cells with two sets of homologous chromosomes are said to be diploid. Gametes—eggs and sperm —are haploid cells. Each gamete contains a single set of chromosomes. At fertilization, a sperm fuses with an egg, forming a diploid zygote. Repeated mitotic cell divisions lead to a multicellular adult made of diploid cells. The diploid adult produces haploid gametes by meiosis, the kind of cell division that reduces the chromosome number by half. Although sexual life cycles differ with the species, they all involve the alternation of haploid and diploid stages. Web/CD Activity 7E Sexual Life Cycles ● ● The Process of Meiosis Meiosis, like mitosis, is preceded by chromosome duplication. But in meiosis, the cell divides twice to form four daughter cells. The first division, meiosis I, starts with the pairing of homologous chromosomes. In crossing over, homologous chromosomes exchange corresponding segments. Meiosis I separates the members of the homologous pairs and produces two daughter cells, each with one set of (duplicated) chromosomes. Meiosis II is essentially the same as mitosis; in each of the cells, the sister chromatids of each chromosome separate. Web/CD Activity 7F Meiosis Animation ● ● Review: Comparing Mitosis and Meiosis See Figure 7.16 for a review and comparison of the two processes. ● The Origins of Genetic Variation Because the chromosomes of a homologous pair come from different parents, they carry different versions of many of their genes. The large number of possible arrangements of chromosome pairs at metaphase of meiosis I leads to many different combinations of chromosomes in eggs and sperm. This is one source of the variation in offspring that results from sexual reproduction. Random fertilization of eggs by sperm greatly increases the variation. Crossing over during prophase of meiosis I increases variation still further. Web/CD Activity 7G The Origins of Genetic Variation ● ● ● When Meiosis Goes Amok Sometimes a person has an abnormal number of chromosomes, which causes problems. Down syndrome is caused by an extra copy of chromosome 21. The abnormal chromosome count is a product of nondisjunction, the failure of a homologous pair of chromosomes to separate during meiosis I or of sister chromatids to separate during meiosis II . Nondisjunction can also produce gametes with extra or missing sex chromosomes, which lead to varying degrees of malfunction in humans but do not usually affect survival. Chromosome breakage can lead to rearrangements—deletions, duplications, inversions, and translocations—that can produce genetic disorders or, if the changes occur in somatic cells, cancer. Evolution Link: New Species from Errors in Cell Division New species can arise very quickly when errors in meiosis or mitosis create polyploid cells, which have more than two sets of homologous chromosomes. Many plants are polyploid, as well as certain animals. Web/CD Evolution Link New Species from Errors in Cell Division Web/CD The Process of Science Polyploid Plants ● ● Self-Quiz 1. If an intestinal cell in a grasshopper contains 24 chromosomes, a grasshopper sperm cell would contain chromosomes. a. 3 d. 24 b. 6 e. 48 c. 12 2. Which of the following phases of mitosis is essentially the opposite of prophase in terms of nuclear changes? a. telophase d. interphase b. metaphase e. anaphase c. S phase 3. A biochemist measures the amount of DNA in cells growing in the laboratory and finds that the quantity of DNA in a cell has doubled a. between prophase and anaphase of mitosis. b. between the G1 and G2 phases of the cell cycle. c. during the M phase of the cell cycle. d. between prophase I and prophase II of meiosis. e. between anaphase and telophase of mitosis. 4. Which of the following is not a function of mitosis in humans? a. repair of wounds b. growth c. production of gametes from diploid cells d. replacement of lost or damaged cells e. multiplication of somatic cells 5. A micrograph of a dividing cell from a mouse shows 19 chromosomes, each consisting of two sister chromatids. During which of the following stages of cell division could this picture have been taken? (Explain your answer.) a. prophase of mitosis b. telophase II of meiosis c. prophase I of meiosis d. anaphase of mitosis e. prophase II of meiosis 6. Cytochalasin B is a chemical that disrupts microfilament formation. This chemical would interfere with a. DNA replication. b. formation of the mitotic spindle. c. cleavage. d. formation of the cell plate. e. crossing over. 7. It is difficult to observe individual chromosomes during interphase because a. the DNA has not been replicated yet. b. they are in the form of very long, thin strands. c. they leave the nucleus and are dispersed to other parts of the cell. d. homologous chromosomes do not pair up until division starts. e. the spindle must move them to the center of the cell before they become visible. 8. A fruit fly somatic cell contains eight chromosomes. This means that different combinations of chromosomes are possible in its gametes. a. 4 d. 32 b. 8 e. 64 c. 16 CHAPTER 7 The Cellular Basis of Reproduction and Inheritance 23 9. If a fragment of a chromosome breaks off and then reattaches to the original chromosome but in the reverse direction, the resulting chromosome abnormality is called a. a deletion. d. a nondisjunction. b. an inversion. e. a reciprocal translocation. c. a translocation. 10. Why are individuals with an extra chromosome 21, which causes Down syndrome, more numerous than individuals with an extra chromosome 3 or chromosome 16? a. There are probably more genes on chromosome 21 than on the others. b. Chromosome 21 is a sex chromosome and 3 and 16 are not. c. Down syndrome is not more common, just more serious. d. Extra copies of chromosomes 3 or 16 are probably fatal. e. Nondisjunction of chromosome 21 probably occurs much more frequently. LM 600× 11. The following light micrograph shows dividing cells near the tip of an onion root. Identify the stage of mitosis for each of the outlined cells, a–d. (b) (a) (c) (d) ● Go to the website or CD-ROM for more self-quiz questions. The Process of Science 1. A mule is the offspring of a horse and a donkey. A donkey sperm contains 31 chromosomes and a horse egg 32 chromosomes, so the zygote contains a total of 63 chromosomes. The zygote develops normally. The combined set of chromosomes is not a problem in mitosis, and the mule combines some of the best characteristics of horses and donkeys. However, a mule is sterile; meiosis cannot occur normally in its testes or ovaries. Explain why mitosis is normal in cells containing both horse and donkey chromosomes but the mixed set of chromosomes interferes with meiosis. 2. Explore how new species can result from errors in cell division in The Process of Science activity available on the website and CD-ROM. Biology and Society Every year about a million Americans are diagnosed as having cancer. This means that about 75 million Americans now living will eventually have cancer, and one in five will die of the disease. There are many kinds of cancers and many causes of the disease. For example, smoking causes most lung cancers. Overexposure to ultraviolet rays in sunlight causes most skin cancers. There is evidence that a high-fat, low-fiber diet is a factor in breast, colon, and prostate cancers. And agents in the workplace such as asbestos and vinyl chloride are also implicated as causes of cancer. Hundreds of millions of dollars are spent each year in the search for effective treatments for cancer; far less money is spent on preventing cancer. Why might this be the case? What kinds of lifestyle changes could we make to help prevent cancer? What kinds of prevention programs could be initiated or strengthened to encourage these changes? What factors might impede such changes and programs? Should we devote more of our resources to treating cancer or preventing it? Why? Go to www.essentialbiology.com to explore the media for Essential Biology. Sample chapter from Essential Biology by Neil A. Campbell and Jane B. Reece © 2001 by Benjamin Cummings, an imprint of Addison Wesley, Inc. 24 UNIT TWO Genetics