References Cited
advertisement

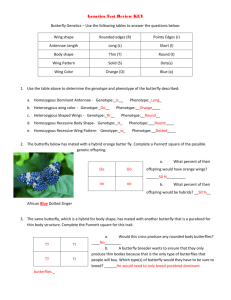
Butterfly Wing Pattern Morphology. E. J. G. May 27, 2004 Biology 454; Term Paper The study of butterfly wing pattern morphology has been critical to our understanding of the developmental mechanisms involved in morphological evolution. The modes by which this morphological variation takes place are often initiated by seasonal, environmental, and geographical features of the individual’s environment. In particular we will discuss how variation of color patterns can affect the fitness and survivability of species, the effects of ecdysteroids on the maturing pupae, and the process of mimicry. Introduction Butterflies are amongst the most visually stunning and unique organisms known to man, with approximately 17,000 known species each of which can be distinguished by its intricate wing color patterns (Caroll et al., 1994). These wing patterns are different from the patterns found on other animals (such as zebras, leopards and lionfish) in that they lack the randomness. While stripes and patterns of other animals are unique to each individual like the fingerprint of a human, the individual elements of the butterfly wing patterns can be found in the same place in all individuals of a species and even across genera (Nijhout, 2001a). Butterfly wings are comprised of a forewing and aft wing with ventral and dorsal surfaces often displaying different patterns. These patterns are based on several systems of symmetry and in most species consist of serial repetition of similar pattern elements within individual wing cells (Carol et al., 1994). The systems of symmetry can be broken down into three distinct systems known as the basal symmetry system, the central symmetry system and the border symmetry system (Nijhout, 2001a). Each system is characterized by a pair of parallel pigmented bands that run from the anterior to the posterior of each wing surface. The bands are interrupted by crossing wing veins giving a segmented appearance of individual wing cells. Wing veination patterns are remarkably constant within taxon resulting in an equal number of wing cells for all species of butterflies (Nijhout, 2001a). Small pigmented marks are often found within the border and central symmetry systems that add another dimension to the possible pattern variation. Change in wing patterns is determined by hormonal control the initiation of which occurs through several different modes of selection including seasonal polyphenism, mimicry and sexual selection. Each of the pattern elements that comprise the wings distinctive color patterns has evolved the ability to change independent of the other elements. The diverse pattern variation resulting from this independent change provides scientists with an opportunity to study the morphological evolution as well as its developmental mechanisms of an incredibly diverse taxon of which essentially all species are known (Nijhout, 1991). H. Frederick Nijhout, a Zoology professor at Duke University, is the world’s foremost expert on the evolution and development of butterfly wing pattern morphology which he has been studying for over thirty years. Building on the findings of Schwanwitsch, Suffert and Kuhn, he has made great advances in our understanding of the evolutionary patterning mechanisms and what is commonly refers to as the nymphalid ground plan. Through his research with the genetics of mimicry and pattern formation, symmetry system hierarchy, and the evolution of serial homology, we have been able to identify the specific modes of evolutionary pattern development that produces the many species-specific patterns found in butterflies. Results The individuated color pattern elements of butterfly wings have evolved from developmentally autonomous, compartmentalized cells within the wings leading to the emergence of serial homology as well as the release from developmental correlation of individual pattern elements (Nijhout, 2001b). It is this loss of developmental correlation that allows the individual butterflies to change one pattern element without affecting another, in turn leading to the great diversity of available patterns we see today. Just as the appearance of the environment can change with the seasons, so to can the appearance of butterfly wing patterns, a phenomenon known as seasonal polyphenism. Many species of butterfly exhibit this form of variation through varying levels of wing melanization in different seasons (Brakefield, 1999). Kingsolver has shown that variation of color can lead to increased relative fitness within Pontia occidentalis, a North American species. In his experiments, Kingsolver (1995a, b, 1996) found that those individuals with darker wing patterns exhibited a higher level of fitness under cooler conditions, where as the opposite was true as the temperature increased. In separate experiments with Precis coenia, Nijhout showed that day length can also lead to selection for specific color patterns. Animals reared under long-day conditions developed a much lighter tan pigmentation (known as the linea form) when compared with the dark reddish- brown coloration (the rosa form) of those reared under short-day conditions. The cause of this variation was determined to be the presence of the pigments xanthommatin in the linea form and dihydro-xanthommatin and ommatin-D in the rosa form. Concentrations of each pigment were then extracted from the wings using an acid methanol solution. The pigments were then analyzed using Thin Layer Chromotography (TLC) and spectroscopy. The presence of each pigment was found to be under control of a developmental biochemical switch that is programmed by the timed release of ecdysteroids during the onset of adult development (Nijhout, 1997). Rountree and Nijhout (1995) discovered that the presence of the ecdysteroids between 24 and 48 hours following pupation would lead to conditions favoring the development of the light tan or linea form while an absence of ecdysteroids during this period produces individuals of the reddish-brown or rosa form (Nijhout, 1997). Aside from thermoregulation, seasonal polyphenism plays a critical role in the effectiveness of the tropical satyrine butterflies ability to avoid predation. Since most of these species live close to the ground, it is imperative that they be able to blend in with the seasonal vegetation and leaf liter to avoid would-be predators. During the dry season when the butterflies experience long periods of inactivity, it has been observed that individuals lose many of the conspicuous ventral wing markings such as the marginal eyespots and medial bands that are prominent in the wet season forms (Brakefield, 1999). These medial bands and large eyespots are thought to serve as camouflage by helping to disrupt the outline of the wing when the wing is closed and the butterfly is perched among the leaves and foliage. In the dry season however these traits are thought to attract predatory birds that may be searching for food on the ground and so selection works against them (Brakefield, 1997). To further demonstrate the importance of this variation, Brakefield (1999) released butterflies of the wet season form and dry season forms with paintedon eye spots during the dry season and recorded markedly lower rates of survival for both groups demonstrating the effects of seasonal pattern composition on an individual’s fitness. Mimicry is another form of adaptation undertaken by butterflies and a direct result of their wing pattern morphology. Turner (1984) stated that Batesian and Mullerian mimicry both begin with a single mutation of large effect resulting in convergence on an intermediate pattern by a lessprotected species with a better-protected one (Nijhout, 1991). Many species such as the Papilio dardanus found in sub-Saharan Africa, are polymorphic and have evolved the ability to mimic between four and six different models of unpalatable species. Control of this polymorphism has been identified as a single genetic locus containing 10 alleles of which four are involved in mimicry (Ford, 1936, Clark and Sheppard, 1959, 1960). The significance of the remaining 6 alleles not utilized in mimicry has not yet been identified (Nijhout, 1991). Conclusion The evolution of butterfly wing patterns has been extensively studied and has contributed a great deal to our understanding of morphological evolution and development. I believe that the evolutionary development of butterfly wing patterns is a testament to the effects and importance of evolutionary change in maintaining species survival. Had butterflies not evolved the ability to independently vary their pattern elements they probably would not exist today. The ability of each pattern element to vary independently of the others is the key factor in the development of the wide range of patterns and adaptations on which the fitness and survival of butterflies depends. Butterflies are ideal test subjects in the area of morphological evolution because the are a large group of more than 17,000 known species the majority of which are morphologically similar (Caroll et al., 1994). By studying monophyletic groups of morphologically similar species, it is possible to deduce the path of evolutionary development through the discovery of model parameters representative of specific pattern development (Nijhout, 1991). Once the parameters have been determined, we could experimentally make small changes in the parameters to see if the forms of related species are produced. Groups of parameters can then be used to identify each species and to determine the morphology of their ancestor. Based on the literature I reviewed, I believe that our understanding of butterfly wing pattern morphology is extensive albeit incomplete. Due to the importance of understanding evolutionary processes, I feel that research in this area should remain a primary focus of evolutionary biologists as well as entomologists. It would seems that due to the popularity of butterflies within the general public, it would be easier to gain the backing and financial support required to complete such research. This popularity should be exploited for the good of the scientific community and to further our understanding of evolutionary development. References Cited Brakefield, P.M., French, V., 1999. Butterfly wings: the evolution of development of colour patterns. BioEssays. 21: 391-401 Caroll, S.B., et al., 1994. Pattern formation and eyespot determination in butterfly wings. Science. 265: 109-114 *Clark, C.A., and P.M. Shepard., 1959. The genetics of some mimetic forms of Papilo dardanus, Brown, and Papilio glaucus, Linn. Journal of Genetics. 56: 237-259 *Clark, C.A., and P.M. Shepard., 1960. The evolution of mimicry in the butterfly Papilio dardanus. Heredity. 14: 163-173 *Ford, E.B., 1936. The genetics of Papilio dardanus Brown (Lep.). Transactions of the Royal Entomological Society of London. 85: 435-466 *Kingsolver, J.G., 1995a. Viability selection on seasonal polyphonic traits: wing melanin pattern in Western White Butterflies. Evolution. 49: 932-941 *Kingsolver, J.G., 1995b. Fitness consequences of seasonal polyphenism in Western White Butterflies. Evolution. 49: 942-954 *Kingsolver, J.G., 1996. Experimental manipulation of wing pigment patterns and survival in Western White Butterflies. Am Nat. 147: 296-306 Nijhout, H.F., 2001a. Elements of Butterfly Wing Patterns. J. Exp. Zool. (Molec. Evol. And Dev.) 291: 213-225 Nijhout, H.F., 2001b. Origin of butterfly wing patterns. In: The Character Concept in Evolutionary Biology. (G.A. Wagner ed.), pp. 511-529. Academic Press. Nijhout, H.F., 1991, The Development and Evolution of Butterfly Wing Patterns, Smithsonian Institution Press, Washington, D.C. Nijhout, H.F., 1997. Ommochrome pigmentation of the linea and rosa seasonal forms of Precis coenia (Lepidoptera: Nymphalidae). Arch. Insect Biochem. Physiol. 36: 15-222 *Rountree, D.B., Nijhout, H.F., 1995. Hormonal control of seasonal polyphenism in Precis coenia (Lepidoptera: Nymphalidae). J Insect Physiol. 41: 987-992 *Turner, J.R.G., 1984. Mimicry: The palatable spectrum and its consequences. In The Biology of Butterflies, ed. R.I. Vane-Wright and P.R. Ackery, pp. 141-161. Academic Press, London