oxidative stress induced alterations of gene expression in human
advertisement

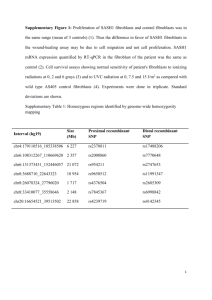
OXIDATIVE STRESS INDUCED ALTERATIONS OF GENE EXPRESSION IN HUMAN FIBROBLASTS HARBORING A8344G MITOCHONDRIAL DNA MUTATION Y.S. Ma1, C.Y. Lu1, Y.C. Chen1, C.Y. Liu1, T. Yang2, Y.H. Wei1 1 Department of Biochemistry and Center for Cellular and Molecular Biology, National Yang-Ming University, Taipei, Taiwan 112, 2CAG Sector of Agilent Technologies, Taiwan, Ltd., Taipei, Taiwan 105 ABSTRACT The effect on gene expression of MERRF (myoclonic epilepsy with ragged-red fibers)-associated A8344G mutation of mtDNA was investigated by Agilent’s human 1 cDNA microarray and RT-PCR. The activation of transcriptional responses, such as the induction of inflammation-related proteins, matrix metalloproteases, and markers of oxidative stress were observed in the patient’s fibroblasts. By contrast, the gene expression of cytoskeleton-related proteins, extracellular matrix proteins, and translation-related proteins were all down-regulated. We also found that the mRNA and activity levels of Mn-SOD, but not catalase and glutathione peroxidase, were significantly increased in skin fibroblasts of the MERRF patient. The intracellular level of hydrogen peroxide was increased in skin fibroblasts of the MERRF patient as compared to those of her family members and age-matched controls. As a result, ROS may not be efficiently removed and in turn elicit an elevation of oxidative stress in skin fibroblasts of the MERRF patient. Our data suggest that the alterations of gene expression may represent the response of the MERRF patient’s fibroblasts to the increased oxidative stress. Taken together, the changes in gene expression imply that inflammation, disruption of mitochondrial reticulum together with degradation of muscle proteins and oxidative damage to tissues are all involved in the pathophysiology of MERRF syndrome. AIM OF STUDY To unravel the genetic and epigenetic factors involved in the phenotypic heterogeneity of skin fibroblasts harboring the A8344G mtDNA mutation. MATERIALS AND METHODS Fig. 1. The pedigree of the MERRF family examined in this study. (A). The patient (IV-1) was a fifteen-year-old female with MERRF syndrome but her younger sister (IV-2) and brother (IV-3) were healthy at the time their skin samples were collected. (B). The proportion of mutant mtDNA of each established skin fibroblasts was determined by PCR-RFLP and analyzed by an Agilent 2100 Bioanalyzer (Lu et al. Clin. Chim. Acta 318: 97-105, 2002). By introducing a mismatched primer, only the PCR products amplified from mutant mtDNA as a template were recognized by Nae I to produce two short restriction fragments of 197 and 26 bp. B, H, and S represent DNA samples purified from whole blood cells, hair follicle cells, and skin biopsy, respectively. Fig. 2. Analysis of differentially expressed genes between the skin fibroblasts from a MERRF patient and an age-matched healthy subject by a cDNA microarray system. Gene expression profiles of these fibroblasts were analyzed by using Agilent’s Human 1 cDNA Microarray Kits (Agilent Technologies, Ltd., CA) that include 12,814 human cDNA clones on an array. The cDNA probes will be made from 20 g of purified total RNA with SuperScript II RT (Life Technologies, Inc., Bethesda, MD) to incorporate the Cy3 or Cy5 dye (NEN). Following purification with the QIAquick spin columns, the purified labeled cDNA probes were concentrated under vacuum in a rotary desiccator until dry. For each microarray to be hybridized, a Cy3-/Cy5-labeled cDNA sample was suspended in deposition hybridization buffer and mixed well with deposition control targets and Cot-1 DNA. After denaturation at 98°C for 2 min, the cDNA probe was transferred to microarray slide, which was then placed onto the base of a hybridization chamber and incubated at 65ºC overnight. The slides was washed sequentially in 2 X SSC, 0.01% SDS; 0.5 X SSC, 0.01% SDS and finally in 0.06 X SSC before subjecting to scan for image analysis using the Agilent's dual-laser Microarray Scanner (G2565AA) combined with Feature Extraction Data Analysis Software (Agilent Technologies, Ltd.). Fig. 3. A comparison between microarray and RT-PCR analysis for several differentially expressed genes between the fibroblasts from normal subjects and MERRF Chen patient. Total RNA were isolated from skin fibroblasts of MERRF patients and age-matched healthy subjects. The cDNA from each of the RNA samples was analyzed by RT-PCR with each gene specific primer pair and quantitated by an Agilent 2100 bioanalyzer. The expression levels of these genes were measured in each run using GAPDH as an internal standard. The results were shown in Table 3.Abbreviation: IL 8, Interleukin 8; CCIS, Complement Component 1s; CC3, Complement Component 3; IFIT1, Interferon Induced Transmembrane Protein 1; IFIP, Interferon, -inducible Protein; IFIP27, Interferon, -inducible Protein 27; IFIPTI, Interferon-induced Protein with Tetratricopeptide repeats 1. Table 1. Clinical features of patients with MERRF syndrome. Patient IV-1 M1 M2 Age / Sex Age of onset Myoclonic epilepsy 15/F 12 ++ 18/F 11 + 14/F 10 - Muscle atrophy +++ + + Cerebellar dysfunction: ataxia Mental retardation Ragged-red fibers MtDNA mutation in fibroblasts A8344G mutation ++ + + + - + + + + + + + 84 86 55 Proportion of the mutant mtDNA (%) IV-1is the patient of the pedigree examined in Figure 1. M1 and M2 are the two unrelated MERRF patients. RESULTS Table 2. Global view of transcriptional alterations in patient with MEERF syndrome compared with normal subject. According to the extraction result created by Feature Extraction software, non-uniform features flagged by the algorithm of software were manually eliminated to increase the reliability of the data. The genes that differentially expressed by 1.5 fold between the skin fibroblasts established from the patient with MERRF syndrome and normal subject were picked up and grouped the genes according to their classification. Table 3. The activities of free radical scavenging enzyme Mn-SOD and catalase and the relative levels of superoxide anions and hydrogen peroxide in the skin fibroblasts from the MERRF patient and her family members. Subject IV-1 III-10 IV-2 IV-3 Activities of free radical scavenging enzyme (mole/mg/mg) Mn-SOD 22732* 14826 12529 13141 Catalase 9.50.7 9.32.4 5.50.6* 11.23.0 Contents of superoxide anions and hydrogen peroxide Superoxide anions Hydrogen peroxide 8112 9627 12135 9038 150 9* 8514 18349* 54 7* CAT activity was determined by monitoring the rate of decomposition of H 2O2 by the decrease in the absorbance at 240 nm (Methods Enzymol 105:121-126, 1985). The total SOD activity was assayed by monitoring nitroblue tetrazolium (NBT) reduction according to Spitz and Oberley (J. Cell. Physiol. 175:359-369, 1998). Western blot for Mn-SOD and CAT in each sample were also carried out and similar results were observed. The contents of superoxide anion and hydrogen peroxide were relative intensities of hydroethidine (HE) and 2’, 7’-dichlorofluorescin diacetate (DCF), respectively, as compared with normal subjects with no mtDNA mutation and measured by staining with HE and DCF and then analyzed by flow cytometry. All the data in this table were obtained by three independent experiments and are expressed as mean SD. *Significantly different from normal subjects, p < 0.01. between three and six population doubling time. The cell lines studied were (A) Intracellular pro-MMP-1 Extracellular pro-MMP-1 GAPDH IV-1 Relative activity (fold to normal control) (B) III-10 IV-2 20 IV-3 N Active MMP-1 Total MMP-1 15 10 5 0 IV-1 III-10 IV-2 IV-3 N Fig. 4. Relative level of secreted MMP-1 in skin fibroblasts from the MERRF patient and her family members. (A) Western blot of intracellular and extracellular pro-MMP-1. (B) Relative activity of MMP-1. The acticvity of MMP-1 in cell culture medium were measured by Fluorokine E Active MMP-1 kit (R & D Systems). This kit allowed us to measure the levels of both endogenous active MMP-1 in cell culture medium based on the fact that MMP-1 can be activated by p-aminophenylmercuric acetate (APMA) in the assay mixture. Fig. 5. The abnormality of the distribution of mitochondria in the skin fibroblasts established from the MERRF patient. The fibroblasts were stained with Mito Tracker Red. Compared with extended and clustered mitochondria of a normal subject, the skin fibroblasts of the MERRF patient showed small and isolated mitochondria are revealed by a confocal microscopy. (A) and (B) represent skin fibroblasts of MERRF patient (IV-1) harboring A8344G point mutation in mtDNA; (C) and (D) indicate skin fibroblasts of a normal subject. DISCUSSION The differentially expressed genes between the skin fibroblasts from a MERRF patient and an age-matched healthy subject were analyzed by a cDNA microarray system developed by Agilent Technologies (Figs. 1-2 and Tables 1-2). To diminish the systematic error, the data were obtained by duplicate experiments and dye swap. Based on the excellent performance of Agilent bioanalyzer on nucleic acid analysis as described recently (Lu et al. Clin. Chim. Acta, 318: 97-105, 2002), we performed RT-PCR on these target genes and quantitated by the Agilent bioanalyzer. We found that the genes of several cytoskeleton-related proteins like actin, integrin, and tubulin were repressed and matrix metalloproteinase (MMPs) gene families were up-regulateded in the fibroblasts from the MERRF patient as compared with the controls. By staining the skin fibroblasts with Mito Tracker Red, extended and clustered mitochondria could be observed in the fibroblasts derived from normal subject but the skin fibroblasts of the MERRF patient showed small and isolated (disconnected) mitochondria scattered in the cytoplasm in a rather random manner (Fig. 5). The matrix-degrading enzymes MMPs, which are activated in tissue remodeling and repair during development and inflammation, as well as many interferon-induced proteins were up-regulated. These results could explain the muscle wasting and weakness-related symptoms in most patients with MERRF syndrome. Moreover, we observed up-regulated transcription of manganese superoxide dismutase (Mn-SOD), which correlates with our previous findings that the protein content and enzyme activity of the enzyme were increased in the MERRF fibroblasts. Based on the cellular and clinical observations, we suggest that the communication between nuclear DNA and mitochondrial genome may play a very important role in the pathogenesis of the MERRF syndrome. Further analysis of differential gene expression in specific signaling and metabolic pathways will be performed by a conjugated database and literature integrated software.