Evidence for a gear mechanism in the molecular motor
advertisement

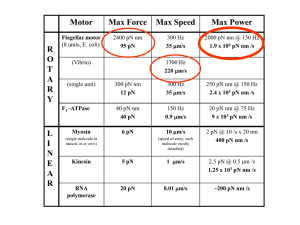
1 Roop Mallik et. al., Cytoplasmic dynein functions as a gear… (Nature Reference Number: 2003-09-09428A) SUPPLEMENTARY MATERIAL (Citations refer to main text) Dynein as a complex and versatile molecular motor Myosin and kinesin each have a single ATP binding/hydrolysis site in the head domain, with the structure of this head being almost identical between the two motors. The dynein heavy chain, which encodes the dynein head domain, is massive (4600 amino acid residues, almost twice the size of myosin II and four times of kinesin3) and is a member of the AAA superfamily of mechanochemical enzymes4 (ATPase associated with diverse cellular activities). In this head domain (Fig 4), there are six tandemly linked AAA-domains (AAA #1 − #6) in a ring conformation, four of which (#1 − #4) are likely to bind ATP. AAA#1 is the site where hydrolysis of ATP generates movement23,24. Further, while in kinesin and myosin the head domain directly binds to the MT or actin filament, dynein uses an ATP-sensitive stalk-like projection on its head to make contact with the MT4,5. The evolutionary cost in design of a motor as massive and complicated as dynein can be understood if one appreciates the wide variety of functions expected from cytoplasmic dynein. While there are many specialized kinesin family members, only a small number of cytoplasmic dynein variants function both during mitosis and in the transport of a multitude of different organelles such as endosomes, lysosomes, Golgi etc. Further thoughts on the proposed gear mechanism in dynein Since the site of ATP hydrolysis (AAA#1) is far separated from the MT binding site (see Fig 4), it is not clear how conformational changes are transmitted back and forth between AAA#1 and the MT binding site4. The intermediate AAA domains (#2−#4) have been implicated in mediating and regulating this transfer of power 3-5,22,25, with AAA#3 being essential for motor function24. The nature of this regulation remains unclear. The mechanism by which nucleotide binding/release at these sites may influence the power stroke and force-production capability of dynein has not been investigated prior to this work. Our work suggests that the regulatory region is not merely a passive linkage, which transmits power from AAA#1 to the MT binding stalk. Rather, it plays a more active role through the binding of ATP on-demand when the motor is straining under load, in such a way that a larger force can be produced to overcome load. Different domains in the regulatory region (AAA#24) could bind nucleotide randomly, or one-by-one in a predetermined way as a function of load, leading to downshifting of gears. Determination of the exact correlation between ATP binding at specific sites in the regulatory domain and observed step sizes (8, 15, 24 and 32 nm) will be a subject of future investigation. Dynein function is not impaired at low ATP To exclude the possibility that the lower stall force is an artefact of protein degradation at low ATP levels, we measured the run length of dynein coated beads at 100 μM ATP. The distribution of run lengths (see supplement, Fig 3) is well fit by a single exponential decay with decay constant = 571±11 nm. This value is comparable to the mean run length of 700 nm reported30 for cytoplasmic dynein at 2 mM ATP. Thus, while the force-production capability of cytoplasmic dynein is inhibited by decreased levels of ATP, its performance in producing processive motion remains unchanged. In this respect, cytoplasmic dynein differs from Tetrahymena ciliary dynein, which is processive17 only at ATP concentrations less than 20 uM. Coupling of steps to ATP hydrolysis Video tracking of dynein-driven bead position at low ATP was used to estimate the residence time of the motor at each step position. The observed distribution of waiting times between steps was found to be a single decaying exponential (Supplement, Fig. 4). This suggests a single rate-limiting step in the enzymatic cycle. Since ATP concentration is low, each ATP arrival constitutes a unique event. Thus a single rate-limiting step suggests that only one ATP is required for the motor to advance by one step. This method was previously used to show that kinesin has a 1:1 coupling between ATP hydrolysis and stepping19. Validation of video tracking results The single exponential decay of residence times (supplement, Fig 4) suggests that motor motion is governed by a Poissondependent process, with a constant probability of ending residence at a particular position (through ATP binding). Since such a process is stochastic, successive steps may sometimes occur in quick succession, even at low ATP concentrations. When tracking motor position with the limited time resolution of video tracking (30 frames/second), several rapid steps of a smaller size could be hidden in a measured step. Reliable estimation of step size is further complicated by the floppiness and large size of the dynein molecule, which results in a spread (s.d. ≈ ±10 nm) of data points for the measured bead position along MT axis 2 (see Figure 3a). This floppiness is also evident from the large sideways excursions 7 of ~100 nm perpendicular to the MT (data not shown) of the bead during dynein-driven motion. To verify that the histogram of step sizes at no load obtained from video tracking (Fig. 3b, main text) reflects the underlying periodicity of step sizes, we simulated dynein-driven motion by using a nanometer-resolution piezo stage to step a dyneinbound bead in known displacements. Dynein coated beads were allowed to bind to immobilized MTs in the absence of ATP and at high (1mM) ADP concentrations. Under these conditions, the enzymatic cycle cannot be completed, and the motor is expected to remain tightly bound to MTs. Video tracking showed that such beads were not motile, but reproduced the observed floppiness of dynein-driven beads. These beads were then moved with known step sizes in a Poisson-distributed way using the piezo-electric stage. Because the observed average bead velocities during low-ATP driven dynein motility were typically 3 to 5 nm/sec we set the mean velocity of such Poisson runs to 5 nm/sec. Bead positions (and step sizes) in video records were estimated by finding the mean position of a bead between steps. Means were taken only over periods of residence exceeding 10 video frames (= 300 ms). This criterion was found to give a reliable estimate of the stepping behaviour. Fiduciary markers were also placed in the field of view in the form of beads firmly stuck to the coverslip. First, we generated Poisson runs on ADP-bound beads with a fixed step size chosen from 8, 15, 24 or 32 nm. Steps of 15, 24 and 32 nm are easily detected with the video tracking (data not shown). In video tracks for Poisson runs with 8 nm steps (supplement Fig. 5a), we find that the steps are more difficult to detect. This difficulty arises from the floppy nature of the dynein linkage, and not due to problems with the video tracking system (See supplementary Fig. 5a: note that 8 nm Poisson steps of a rigidly fixed fiduciary marker bead in the same field of view is reproduced cleanly, while the same is not true for a dynein-bound bead). To estimate the error arising out of this floppiness of the bead, we extracted step size (see supplement Fig. 5b) from the 8 nm step Poisson run of the dynein bound bead using the criterion mentioned above. We find a spread of step sizes about the expected mean value of 8 nm, with occasional steps larger than 10 nm being detected. However, the detected steps are rarely larger than 15 nm. This shows that the 15 nm, 24 nm and 32 nm steps detected in dynein-driven motion (Fig. 3a, 3b) are indeed single steps, and not artefacts of multiple 8 nm steps. Certain segments of data (for example, 6 sec to 14 sec in supplementary Fig. 5a, marked by red vertical arrows) show exaggerated flop in the bead. Such data segments are not used in the step size estimation. We observe such unusable segments of data for dynein-driven motion also, and reject the same when looking for steps. Next, we asked the question: does the histogram of step sizes in Fig 3b truly reflect the underlying periodicity of motion ? To investigate this, we stepped the piezo-electric motor in a Poisson-dependent way, but with a weighed mixture of step sizes of 15 nm, 24 nm and 32 nm. The probability of choosing steps of 15, 24 and 32 nm was respectively 19%, 41% and 40%. These percentages were obtained by grouping the counts in Fig. 3b into intervals of 8 nm centred at 15 nm (19%), 24 nm (41%) and 32 nm (40%). Steps close to the expected values were detected in such mixed Poisson runs (supplement Fig. 6a). The histogram of step sizes obtained from such mixed step size Poisson runs looks similar to that of dynein-driven motion (Fig. 3b). This provides strong evidence in favour of dynein-driven motion consisting of a mixture of 15, 24 and 32 nm steps under no applied load. Other reports of step size larger than 8 nm in dynein Large steps of ~ 24 and 32 nm in cytoplasmic dynein are surprising, considering that only 8 nm steps have been reported for axonemal dynein12,17. However, there have been indications about a possibility of larger step size for dynein in the literature. Electron microscopic studies15 on axonemal dynein predict a 15 nm powerstroke, with the possibility of a range of values around this mean due to flexibility of the dynein molecule. Of particular interest is a report18 where axonemal dynein arms produce oscillatory displacement of a singlet MT with an amplitude of ~30 nm, which was interpreted by the authors as multiple 8 nm steps of the dynein head. These oscillations could also be interpreted as large back-and-forth single steps of the axonemal motor. Such steps would be consistent with the unloaded stepsizes (24 and 32 nm) we observe for cytoplasmic dynein, and the 30 nm displacement could be an indication of the intermediate step discussed earlier (see main text). In the axonemal dynein experiment, however, the geometry of the assay is different from what is reported in the present investigation. The 30 nm oscillations are observed when the dynein head sits on one MT and generates force against a second MT which is almost perpendicularly oriented. In this perpendicular geometry, there is limited scope for dynein-driven processive sliding of MTs, thus the motor might be constrained to take single backand-forth steps. Interestingly, if these two MTs are oriented parallel (or close to parallel) to each other, then the back-and-forth oscillations cease, and the second MT glides over long distances. Shingyoji et. al. do not report the “step size” of the gliding molecule in the parallel oriented case. In light of present work, it might be worthwhile to see if the 30 nm oscillatory behaviour changes to successive steps of ~24 or 32 nm when the geometry is favourable for processive motion.