Surfactant and its role in pulmonary inflammation.
advertisement

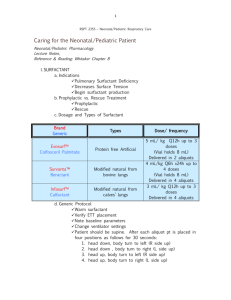
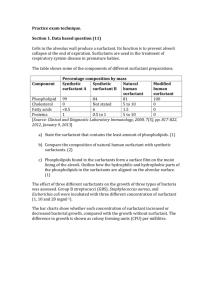
SURFACTANT AND ITS ROLE IN PULMONARY DISEASE Professor Ron Slocombe, School of Veterinary Science. University of Melbourne, Victoria, Australia. Abstract: Over 40 years ago it was recognized that the qualities of lung epithelial lining fluid in reducing surface tension were essential for life. A recent word search using “PubMed” revealed over 8 thousand citations for pulmonary surfactant. Early research was directed toward the phospholipid components of surfactant and its effects on surface tension. In the last decade, an increasingly diverse number of important biologic processes involving surfactant components have been identified. Surfactant lipids and proteins are now known to have roles in modulating fluid flux across air-tissue interfaces, in the recognition of pathogens and in either inducing or suppressing inflammation. An extensive series of reviews on pulmonary surfactant was published in November, 1998 and much of the emerging information about the surfactant proteins is published here. (see Biochimica et Biophysica Acta (1998), 1408:1-289.) In addition, the isolation and elucidation of roles for surfactant proteins in health and disease offers new hope for serologic testing to diagnose interstitial lung disease, as the appearance of these proteins in increasing amounts in the blood correlates with interstitial lung injury. Currently the application of these methods to veterinary medicine is limited by the lack of suitable reagents, and most current data on animals is restricted to experimental laboratory animals, reptiles and horses. The measurement and use of surfactant components in the diagnosis and therapy of lung disease presents exciting new possibilities within the next decade. _____________________________ The nature and function of surfactant lipids. Pulmonary epithelial lining fluid (ELF), surfactant, in mammals has a relatively constant composition of about 90% lipid and 10% protein. Biophysical properties relating to surface tension reduction reside principally with the phospholipids in surfactant, and of these dipalmitoylphosphatidylcholine (DPPC) is the most important, constituting about 50% of the total phospholipid and responsible for most of the surface-tension reducing capacity. Other phospholipids are present in smaller amounts and in some species there are significant alterations in minor phospholipids with maturation from the fetal state to the adult. Neutral lipids, including cholesterol and esterified cholesterol, and fatty acids are present in surfactant in low amounts, but have no known function. It has been postulated that these and the minor phospholipids might enhance the reconstitution of DPPC if displaced from surface films. The study of surfactant lipids has been hampered by difficulties in accurately and easily determining individual lipid components. In addition, for an adequate understanding of the role of surfactant lipids in vivo, integrated studies that analyze biochemical composition and relate these to biophysical properties are needed. To date, this has not been done in animals. Production, use and degredation of surfactant lipids. Surfactant lipids are produced by alveolar type II pneumocytes and to a lesser extent by bronchiolar non-ciliated epithelial cells. These lipids are synthesized into preformed intracytoplasmic lamellar bodies, secreted into the aqueous subphase of ELF, where they unravel to form tubular myelin figures, and from tubular myelin are exchanged with surface film phospholipids. The movement of DPPC from lamellar bodies to surface active film is too rapid for simple diffusion. Neutral lipids and hydrophobic surfactant proteins are though to facilitate this process. This is essential because cyclic stretching and compression of ELF during rhythmic breathing causes displacement of surface lipids back into the subphase, particularly during lung collapse, and without rapid restitution of the surface-active molecules, successive cycles of lung deflation would inexorably lead to further lung collapse. The surfactant lipids, dispersed in the subphase, may also be degraded by both alveolar macrophages and pneumocytes, due to the action of the serine protease, surfactant convertase. In lavage fluids, dense aggregates of surfactant are thought to represent tubular myelin and light unilamellar vesicles degraded, inactive surfactant. Surfactant may be lost by airway removal, further degraded or resynthesised into lamellar bodies along with surfactant proteins. This intimate relationship between recycled components of ELF, macrophages and epithelial cells provides pathways in addition to those involved in surface tension for antigen processing and recognition. Manipulation of surfactant lipids. Immature fetal lungs and those severely damaged by acute interstitial lung disease may develop oedema and collapse because of surfactant dysfunction. The influence of corticosteroids in promoting maturation of type II pneumocytes in preterm lungs is well known. However, exogenous administration of surfactant lipids in cases of lung disease is not widely practiced in veterinary medicine. Purified DPPC has marginal effectiveness as a surface tension reductant, and although significant improvements in activity are produced by the addition of hydrophobic surfactant proteins, these proteins differ sufficiently between species and therefore exogenous complex surfactants designed for human use may not be as effective in animals. Several studies have investigated whether dietary manipulations can alter surfactant lipid composition. Manipulation of dietary lipids or vitamin A in general has minimal effect on surfactant composition, and presumably has no functional effect on biophysical properties. Alteration of the lipid composition of surfactant with changing temperature occurs in reptiles but the function of such changes remains unclear. Surfactant lipids, inflammation and infectious agents. Inactive or inadequate surfactant lipids lead to lung collapse and predisposes to pulmonary edema. Surfactant is not only subject to degradation by surfactant convertase, but oxygen radicals, phospholipases, leukocyte elastases and other proteolysins generated during inflammation can reduce its activity. Inhibition of proteolysis by addition of 1 antitrypsin prolongs surfactant function and improves oxygenation in surfactant deficient rats. Inflammation associated with allergic lung disease has also been shown to increase ELF protein and inversely correlate with surfactant activity, leading to increased airway obstruction. Surfactant dysfunction often occurs following lung transplantation, ARDS and in ischemia-reperfusion injury (IRI). Previously, it has been assumed that surfactant was inactivated by proteolysins in edema fluid that developed as a consequence of inflammation, but recently experimental IRI studies found that surfactant dysfunction preceded protein leak and edema formation. Surfactant dysfunction occurs with septic pneumonia, and replacement improves lung function and speeds clearance of organisms. Some of these effects may be indirect, since better reduction in surface tension in pneumonic lung lessens edema, improves oxygenation, and facilitates mucociliary clearance. Surfactant has additional beneficial effects, suppressing the growth of bacteria directly, serving as an opsonin for phagocytosis and affecting the generation of nuclear factor B, critical in the regulation of many cytokines. Some of these functions may be more closely related to actions of surfactant proteins rather than lipid components. Surfactant dysfunction also occurs in horses that develop EIPH, and in horses following transport stress, when it is associated with sepsis. The hydrophilic Surfactant Proteins. There are 2 proteins in this group, SP-A and SP-D. Both belong to the collectin family of proteins, responsible in part for innate immunity and mucosal defences. These proteins may be especially important in preventing infections in neonates prior to the development of aquired immunity. SP-A is secreted independently of lamellar bodies, is also secreted by Clara cells and to a lesser extent by conducting airway epithelium. It appears to be under feedback control and influences the amount of total surfactant present. SP-A is the most abundant surfactant protein and is required for surfactant to effectively lower surface tension. Excess SP-A in the subphase hastens lipid degradation and recycling and suppresses further lipid secretion by Type II cells. Receptors for SP-A and SP-D are present on both type II cells and alveolar macrophages and SP-A enhances the chemotaxis, phagocytosis and killing of micro-organisms in the lungs. There are conflicting reports of the influence of SP-A, -D on cytokine production, the range of inflammatory cells that carry receptors for it and whether inflammatory cells exposed to SP-A, -D release oxygen radicals and enzymes. SP-A may also assist in protecting surfactant from inactivation by plasma proteins during inflammation. SP-A and SP-D bind carbohydrates and lipids and this binding has been associated with antimicrobial functions, including a range of viruses, bacteria, endotoxin, fungi and Pneumocystis. SP-A deficient animals are susceptible to lung infections but do not develop respiratory failure through surface tension abnormalities. Both SP-A and SP-D bind pollen and mite antigens, influence lymphocyte function and therefore may modulate allergic reactions in the lungs. Serum levels of these proteins rise with alveolar injury, infection and SP-A also in association with lung carcinoma. Equine SP-D has recently been isolated. The hydrophobic Surfactant Proteins. There are 2 proteins in this group, SP-B belonging to the saposin family of proteins, and SP-C. These proteins are thought to have similar functions and are critical for the effective secretion, maintenance of tubular myelin and surface films, and recycling of the surface-active lipid components of surfactant. Inherited deficiencies in SP-B are associated with fatal neonatal respiratory failure. Incorporation of synthetic SP-B into artificial surfactant preparations dramatically improves efficacy. Reviews 1. Pulmonary surfactant. (1998) Biochimica et Biophysica Acta 1408:1-289. 2. Doyle, IR., Nicholas, TE., and Bersten, AD. (1999) Partitioning lung and plasma proteins: Circulating surfactant proteins as biomarkers of alveolocapillary permeability. Clin Exp Pharmacol Physiol. 26: 185-197. 3. Van Iwaarden, JF. (1993) Surfactant and the pulmonary defense system. In B Robertson, LMG Golde and JJ Batenburg, editors. Pulmonary surfactant: from molecular biology to clinical practice. Elsevier, Amsterdam, 215-225. Recent articles on surfactant in animals 1. Belai, Y. et al (1999) Addition of 1-antitrypsin to surfacatant improves oxygenation in surfactant deficient rats. Am J Crit Care Med 159: 917-923. 2. Gunther, A et al (1999) Surfactant subtype conversion is related to loss of surfacatant Apoprotein B and surface activity in large surfacatant aggregates: experimental and clinical studies. Am J Crit Care Med 159: 244-251. 3. Harrod, KS. et al (1999) SP-A enhances viral clearance and inhibits inflammation after pulmonary adenoviral infection. AJP 277: L580-L588. 4. Herting, E. et al (1999) Combined treatment with surfactant and specific immunoglobulin reduces bacterial proliferation in experimental neonatal group B streptococcal pneumonia. Am J Crit Care Med 159: 1862-1867. 5. Hickman-Davis, J. et al (1999) Surfactant protein A mediated mycolplasmacidal activity of alveolar macrophages by peroxynitrite. Proc Nat Acad Sci 96: 4953-4958. 6. Hobo S. et al (1997) Effect of transportation on the composition of broncho-alveolar lavage fluid obtained from horses. Am J Vet Res 58: 531-534. 7. Hobo S. et al (1999) Purification and biochemical characterization of equine pulmonary surfactant protein D. Am J Vet Res 60: 368-372. 8. Morrison KE. et al (1999) Functional and compositional changes in pulmonary surfactant in response to exercise. Equine Vet J Suppl 30: 62-66. 9. Ochs M. et al (1999) Ultrastructural alterations in intra-alveolar surfacatant subtypes after experimental ischemia and reperfusion. Am J Crit Care Med 160: 718-724. 10. Orgeig S. et al (1997) Surfactant regulates pulmonary fluid balance in reptiles. Am J Physiol 273/42: R2013-R2021. 11. Savov, J. et al (1999) Mechanical ventilation of rat lung: effect of surfactant forms. AJP 277: L320-L326.