Best_etal_Madang_tap.. - Memorial University of Newfoundland
advertisement

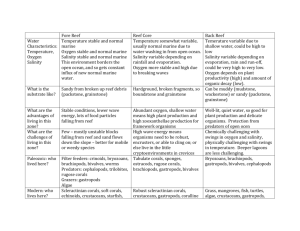
1 EEincorporate the statistical results from adding bivalve total encrustation and total bioerosion data – this was why I did total estimates, but I don’t think I’ve seen the analysis. Best et al., Experimental taphonomy in Madang Lagoon, for PALAIOS Note that some of the nmds ordinations have dispersion among the sediment/water interface specimens, while some have the sediment/water interface specimens clumped, and buried dispersed… Double check names of sites consistent throughout EEUpdate shells in figures MBTable 2? Or in text? Refs, format of citations as well Fig 6 and 8 should be in same format as 7 and 9 Check Format Fig vs fig, Tab… Change abrasion to surface alteration Delete physical vs. biological throughout text Change Exposed vs. buried to sediment/water interface vs. buried Change Mollusks or mollusks for bivalves Update and confirm interpretation of figures in text EXPERIMENTAL TAPHONOMY OF MODERN CORALS AND BIVALVES FROM MADANG LAGOON, PAPUA NEW GUINEA MAIRI M.R. BEST1,2 EVAN N.EDINGER3,4,5*, LAURA M. ESTRADA ALVAREZ5,6 and JOHN M. PANDOLFI7. 1 McGill University Department of Earth and Planetary Sciences, Montreal, QC, Canada; 2 Current Address: University of Victoria, School of Earth and Oceans Sciences, Victoria BC, Canada; 3 Memorial University of Newfoundland, Department of Geography, St. John’s, NL, Canada; 4Memorial University of Newfoundland, Department of Biology, St. John’s, NL, Canada,; 5 Memorial University of Newfoundland, Department of Earth Sciences, St. John’s, NL, Canada; 6Current Address: University of British Columbia, Department of Earth and Ocean Sciences, Vancouver, BC, Canada; 7University of Queensland, Centre for Marine Studies and Department of Earth Sciences, Brisbane, Queensland, Australia. *e-mail: eedinger@mun.ca RRH: EXPERIMENTAL TAPHONOMY OF CORALS AND BIVALVES LRH: BEST ET AL. Keywords: taphonomy, encrustation, bioerosion, exhumation, coral reefs ABSTRACT. This study compared taphonomic alteration of five species of experimentally deployed scleractinian corals and bivalve molluscs on lagoonal coral reefs to assess the influence of burial, onshore-offshore gradients, form, and skeletal microstructure on taphonomic alteration. Artificially killed fresh corals and bivalves were placed on the sediment-water interface, or Best et al., Experimental taphonomy in Madang Lagoon, for PALAIOS 2 buried 10 cm within the reef sediment, at three lagoonal reefs 0.2 - 1.2 km from shore. Experiments were recovered after 1 year, and assessed for physical, chemical, and biological alteration of skeletons. Some buried specimens at all three sites were exhumed, with the highest proportion of exhumation at the most exposed site. Weight loss, surface alteration, and bivalve disarticulation, periostracum and ligament decay, were most strongly influenced by burial/exhumation, with relatively minor differences between reefs. Level of encrustation and bioerosion, and species composition of epibionts and endobionts, were both most strongly influenced by burial, with relatively minor differences between reefs and among experimental taxa. Total encrustation on corals deployed at the sediment water interface was highest on the wave-exposed reef, contrary to results on bivalves and predictions based on productivity of the three sites, and suggesting that burial of sediment/water interface specimens at the inshore site may have restricted encrustation there. Our results emphasize the importance of burial, and of remaining buried, in fossil preservation. Buried corals of all growth forms and both species of bivalves experienced exhumation, confirming that sedimentological time-averaging, involving vertical mixing of bioclasts in the death assemblage, takes place in reef lagoons. Burial Onshore-offshore Form Microstructure Physical, chemical, biological Nutrients (in negative) Best et al., Experimental taphonomy in Madang Lagoon, for PALAIOS INTRODUCTION 3 The degree of taphonomic alteration of skeletal remains depends primarily on rates of burial, frequency of exhumation, and environmental conditions such as temperature, water column productivity and oxygenation, sediment composition and porewater chemistry (Brenchley and Harper, 1998; Pandolfi and Greenstein, 1997a,b; Best and Kidwell 2000a; Lescinsky et al. 2002; Tapanila, et al., 2004; Best et al., 2007). Taphonomic processes in general vary widely between reefal, non-reef carbonate, and siliciclastic environments (Best and Kidwell 2000a; Kidwell et al., 2005; Best et al., 2007), as well as between tropical, temperate, and boreal regions (Best et al. in prep). Furthermore, intrinsic controls on taphonomy stemming from life habit, growth form and skeletal microstructure can be important (Best and Kidwell 2000b; Best 2008; but see also; Kidwell 2005; ). Rapid burial almost always improves quality of preservation, and reduces degree of taphonomic alteration (e.g. Briggs 2000), while highly productive waters favour increased encrustation and bioerosion (Sammarco and Risk 1990; Risk et al., 1995; Holmes et al. 2000; Lescinsky et al. 2002The relative importance of burial, exhumation, sedimentation, and productivity vary among environments, especially along onshore-offshore gradients (Lescinsky et al. 2002; Kidwell et al., 2003; Tapanila et al. 2004; Best et al., 2007; Best, 2008). Terrigenous sedimentation and productivity often co-vary, such that sites in which shells generally experience rapid burial are also likely to have rapid encrustation or bioerosion of sediment/water interface shells (e.g. Holmes et al., 2000; Lescinsky, et al., 2002). Experimental arrays can be a powerful means of determining relatively short-term rates and pathways of skeletal degradation under specific conditions in natural environments. The nature and extent of time-averaging on coral reefs also influences reef taphonomy (Scoffin 1992, Pandolfi 2000). In reefal environments, wave exposure often varies along the same onshore-offshore gradient, and may influence the residence time of bioclasts in the taphonomically active zone by burial and exhumation (Kidwell and Flessa, 1995). Wave energy may also affect the temporal scale of time-averaging, with more hydrodyanimic environments predicted to experience a greater degree of time-averaging than more quiescent environments (Kidwell and Flessa, 1995, Kowalewski, 1996). While coral reefs have been considered to be environments dominated by in-situ preservation, and ecological time-averaging (Edinger et al. 2007), the extensive sediment aprons surrounding the reefs in lagoons may be dominated by taphonomic processes more similar to those in siliciclastic environments (Kidwell 2000), In particular, the extent to which reef bioclasts are subjected to burial –exhumation cycles may be The purpose of this paper is to report on the first combined coral-bivalve experiments and the first taphonomy field experiments on reef corals, and to evaluate the importance of the results for interpreting the quality of fossil preservation in the fossil record. Here, we approach the relative importance of (1) burial/exhumation, (2) onshore-offshore gradients in wave energy and sedimentation rates, and (3) form and skeletal microstructure, on taphonomy of scleractinian corals and bivalve molluscs. We treat the question experimentally, using buried and surface-deployed samples in the modern reef lagoon setting of Madang Lagoon, Papua New Guinea. Although actualistic taphonomic studies of bivalve molluscs in temperate siliciclastic environments are more common (Kidwell refs, Flessa refs, ), there are relatively few tropical studies of modern coral (Kiene and Hutchings 1994; Chazottes et al., 1995; Pandolfi and Greenstein, 1997a,b; Greenstein and Pandolfi 1997) or bivalve (Parsons-Hubbard, 2005; Best, 2008) taphonomy, and no previous experimental studies on tropical coral taphonomy. Best et al., Experimental taphonomy in Madang Lagoon, for PALAIOS 4 We predicted that skeletons deployed at the sediment/water interface would be more affected by biological and physical processes such as encrustation, bioerosion, fragmentation, and surface alteration than buried skeletons. Those buried should have been more affected by chemical processes, such as precipitation and dissolution due to the early diagenetic evolution of sediment porewaters. We also predicted differences between sites, expecting highest levels of encrustation and bioerosion at the nearshore site with highest productivity, and highest levels of surface alteration at the offshore site with greatest wave exposure. Finally, we expected differences in taphonomic alteration among corals and bivalves based on density, surface to volume ratio, and organic content, for example expecting a lesser degree of alteration to massive corals than to branching and free-living corals, based on their more compact and denser morphology and apparently greater resistance to fragmentation, and expecting that bivalves with high organic content in their skeletons would experience more rapid weight loss and greater endobiont infestation than those with lower organic content. Our results emphasize the primacy of burial, and the importance of exhumation in dynamic environments of reef lagoons. MATERIALS AND METHODS Field sites. Experiments were deployed in three shallow reef settings along an onshore-offshore gradient within Madang Lagoon, Papua New Guinea (Fig. 1). The mixed carbonate-siliciclastic fringing reef at the furthest inshore site, Nagada Harbour, experiences low wave exposure, moderate siliciclastic sedimentation, moderate turbidity, and moderate productivity under outer estuarine conditions. The back lagoon fringing reef at Gosem Island experiences moderate levels of wave exposure, low-moderate productivity, and relatively little siliciclastic sedimentation. Paddoz Reef is a mid-lagoon patch reef that experiences moderate to high levels of wave exposure, low productivity, and virtually no siliciclastic sedimentation (cf. Langer and Lipps, 2003). Experiments were deployed on reef slopes at 4 m water depth in Nagada Harbour and Gozem Island, and on a reef crest at 2 m water depth at Paddoz Reef (Table 1). Paddoz reef experiments were deployed at 2 m, rather than 4 m, because the steep reef slope between 2 and 6 m precluded burial experiments at 4 m depth. Experimental design and taxa The experiment followed a balanced 2-way ANOVA design. At each site, four replicate pairs of each of the experimental species were deployed both (a) tethered at the sediment water interface and (b) tethered and buried 10 cm below the sediment/water interface. Each design was replicated 4 times, for a total of 72 corals and 48 articulated bivalve shells (96 valves). Three growth forms of corals were deployed: branching (Acropora pulchra), massive (Goniastrea retiformis), and free-living (Fungia sp.; Figure 2). Two bivalve forms were studied: Gafrarium tumidum, which has low organic microstructure, compact form, ribs, and colouration; and Geloina coaxans, which is a large, thin-shelled bivalve with high organic microstructure,periostracum, and a higher surface to volume ratio. Coral samples were killed in freshwater bleached, and air dried. Bivalve samples were shucked, and shells maintained in salt water prior to deployment to avoid breakdown of organic microstructure . All were tagged, weighed, and photographed prior to deployment. Coral and bivalve specimens were checked after 1.5 months, and were retrieved after 12 months. Taphonomic characters assessed. Best et al., Experimental taphonomy in Madang Lagoon, for PALAIOS 5 Degree of taphonomic alteration of both corals and bivalves was assessed based on weight change, discolouration, surface alteration, fragmentation, encrustation, and bioerosion. In addition, disarticulation and loss of periostracum were also assessed for the bivalves. Weight loss was calculated as the difference in sample weight before and after experimental deployment divided by the initial weight. Discolouration and surface alteration on experimental corals and bivalves were measured using a 5-point ordinal scale (Pandolfi and Greenstein 1997; Estrada et al. 2004; Best and Kidwell 2000a; Best et al. 2004). Each coral or shell was determined to be whole or fragmented by visual comparison with its pre-experimental photograph. Total encrustation was measured as percent cover of individual encrusting taxa and as total percent area encrusted. Encrustors were identified to relatively high taxonomic level: spirorbid and serpulid polychaete worms, bryozoans, bivalves, sponges, coralline algae, foraminifera, coral spat, vermetid gastropods. Endolithic bioerosion was measured as the frequency of boreholes visible on the external surface of each specimen. Boreholes were identified to the following taxonomic groups: unlined and lined polychaete worms, clionid sponges, and endolithic bivalves Lithophaga sp. and Gastrochaena sp. The borer count data were square-root transformed prior to ANOVA in order to reduce and homogenize the variance. Data analysis. Presence-absence of each encruster or macroborer taxon was recorded on each replicate coral or mollusc valve, and the average proportion of samples for each treatment-reef-species block of the experiment encrusted (bored) by that epibiont was calculated on a scale of zero to 1 (i.e. zero if no samples encrusted or bored, 1 if all samples encrusted (bored)). Encrusting organisms were scored as presence/absence to equalize the treatment of large and small encrustors. Individual increments in figures 7 and 9 indicate relative frequency of occurrence of each encruster or borer taxon on each substrate type for each treatment (0-1), and the sum of all the different encrusters or borers represented in each treatment-reef-species block. Average species richness of epibionts and endobionts was compared using 2-way ANOVA, testing for effects of treatment (sediment/water interface, buried), reef (Paddoz Rf. Gosem Is., Nagada Hr.), and experimental taxon and interactions. Variation in encrustor or macroborer species composition between treatments, and among reefs and experimental taxa, was assessed using non-parametric ordination (non-metric multidimensional scaling) and analysis of similarity (ANOSIM; Clarke and Warwick 1993) in PRIMER 6.0. Bray-Curtis similarity among treatments, reefs, and taxa was calculated using the proportional occurrence data, based on presence-absence on replicates. Relative contribution of different encrustor or macroborer taxa to the differences between treatments, reefs, or experimental taxa were measured using the similarity percentage (SIMPER) routine in PRIMER 6.0 RESULTS Experimental recovery. All coral and bivalve specimens were present after 1.5 months. After one year, however, 63 of 72 (87%) coral specimens were recovered. Most of the corals lost were from the most exposed site, Paddoz Reef. Among bivalves, only 76 of the 96 (79%) bivalve shells were recovered after 1 year, with all losses at the high energy Paddoz Reef site (Table 2). Best et al., Experimental taphonomy in Madang Lagoon, for PALAIOS 6 Net weight change. Weight change (as % of initial weight) was dominated by weight loss, which was generally greater in sediment/water interface specimens (Fig.4). Sediment/water interface coral specimens showed greater weight loss than buried specimens (sediment/water interface: avg. = -11.51%, buried avg. = -4.92%, ANOVA, F(1,57) = 5.83, p<0.02. Data suggesting weight gain observed among sediment/water interface Acropora corals at Paddoz Reef and buried Acropora corals at Nagada Harbour is limited by sample size; weight change in many of the corals was not significantly different from zero. Weight change among the bivalves was not significantly different from zero in any case except for sediment/water interface Gaffrarium in Nagada Harbour, which experienced slightly greater weight loss than buried Gaffrarium. A tendency towards weight gain in individual specimens as a result of encrustation was not resolved given low n. Due to lost specimens, no error bars could be calculated around weight change in buried Geloina at Paddoz Reef. Bivalve fragmentation and disarticulation data are presented in Table 2. Fragmentation only occurred with Geloina coaxans at Paddoz, after one year of deployment. Disarticulation of the valve pairs had occurred for most samples after one year, with a slightly lower occurrence among buried pairs in the lower energy sites. Surface alteration. Surface alteration, the result of a combination of microboring, dissolution, and abrasion, of corals and bivalves entailed loss of surface detail in corals, and chalkiness to erosion in bivalves. Surfaces can also suffer secondary discolouration. Figure 4 shows details of surface alteration of buried and sediment/water interface pairs of each experimental species of coral and bivalve. Table 2 provides information on loss of periostracum in Geloina coaxans, note that … Figure 5 compares the average degree of surface alteration of each species on each reef, in both sediment/water interface and buried treatments. Sediment/water interface corals experienced significantly more surface alteration than buried corals (1-way ANOVA, F(1,57) = 28.23, p<<0.001, Fig. 5), but there were no significant differences in surface alteration among sites or coral growth forms, nor interactions between those variables and burial. Surface alteration was not discernable in most of the sediment/water interface bivalves, because the original shell interior surfaces were obscured by encrustation, so statistical treatment of surface alteration data in the bivalves was not possible. Initially buried specimens deployed at both Paddoz and Nagada showed surface alteration, whereas those at Gosem were pristine. Discolouration was higher in buried corals than in sediment/water interface corals at the innermost site (Nagada Harbour), but the sediment/water interface corals were more discoloured than buried corals at the other two sites (2-way ANOVA, effect of treatment F=5.07, p=0.028; interactions F=3.73, p=0.031). Surface alteration (mainly abrasion) was significantly higher in sediment/water interface corals than in buried corals (F=25.7, p<0.0001, but there was no significant effect of reef on surface alteration, nor any interaction among treatment and reefs. Discolouration and surface alteration were difficult to discern in many of the sediment/water interface bivalves because the shell surface was completely Best et al., Experimental taphonomy in Madang Lagoon, for PALAIOS 7 covered by encrusting organisms. Discoloration occurred among buried shells at both Gosem and Nagada. Encrustation and Bioerosion. Total encrustation was significantly higher on sediment/water interface corals and bivalves than buried specimens in all environments (Fig. 6, Table 3). When all specimens were considered together, there was only a weakly significant interaction effect between treatment and reef (Table 6). Among both sediment/water interface and buried corals, encrustation was higher at Paddoz Reef than at the other two sites (1-way ANOVA, sediment/water interface: PR>GI≥NH, F(2,23) = 7.21, p<0.005); buried: PR>GI≥NH, F(2,30) = 8.36, p<0.0015. By contrast, among bivalves, the degree of encrustation was highest at Nagada harbour (2-way ANOVA, F(2,65) = 8.53, p<0.001, encrustation on internal surfaces, F(2,65) = 3.81, p<0.0277, encrustation on external surfaces). Buried shells at Gosem Island remained unencrusted, while buried shells at Nagada and Paddoz displayed encrustation, but at lower coverage levels than for sediment/water interface shells at the same sites (Fig 6, Table 3).. Degree of encrustation was generally higher on bivalves than on corals but was only weakly significantly different among coral growth forms. Among the bivalves, total encrustation on shell exteriors was generally higher on Gaffrarium than on Geloina (2-way ANOVA, F (2,67) = 19.35, p<0.001 for although encrustation on shell interior surfaces was not significantly different (2-way ANOVA, F (2,67) = 0.59, p<0.45). Among corals, Fungia was less encrusted than the other coral species, particularly at Gosem Island, and among buried specimens at Paddoz Reef (Figure 6). Species composition of encrustors. -- The most common epibiont taxa were calcareous worms, especially spirorbids, calcareous algae, and foraminifera, mostly the encrusting red foraminifer Homotrema rubrum and vermetids. Other common epibionts included bryozoans, sponges, and juvenile corals (Fig. 7). Encrusting bivalves were uncommon, and occurred only on the sediment/water interface specimens of massive coral Goniastrea and the free-living coral Fungia (Fig.7); there was no variation among sites in the occurrence of encrusting bivalves. Species richness of encrustor taxa was significantly greater on sediment/water interface corals and bivalves than on buried substrates (2-way ANOVA F(1,2)=52.0, p<0.001, and was higher at Paddoz Reef than at the other two sites (2-way ANOVA, F(2,2)=4.75, p=0.019), but did not differ among taxa of sediment/water interface corals and bivalves, nor were there significant interactions among treatment, reef, or taxon. Among buried corals, species richness was significantly greater at Paddoz Reef than at Gozem Island or Nagada Harbour (2-way ANOVA, F(2,58)=3.146, p=0.034), but this pattern did not hold for buried bivalves, probably because those at Paddoz and Nagada were exhumed. The species composition of encrusting organisms varied significantly among treatments (surface, buried; ANOSIM, R=0.348, p<0.0001), but not among experimental sites (R=0.0026, p=0.44) or forms (R=-0.006, p=0.52). Total bioerosion.-- Total bioerosion was significantly higher on sediment/water interface corals and bivalves than on buried specimens in all environments (Fig. 8; Table 4,). There was a significant difference in bioerosion between sediment/water interface and buried bivalves, at Gozem and Nagada Gafrarium tumidum, Paddoz could not be resolved for bivalves because of low numbers after loss of Best et al., Experimental taphonomy in Madang Lagoon, for PALAIOS 8 sediment/water interface Geloina specimens. There was no effect of reef, and no significant interactions effect between reef and treatment, when both corals and bivalves were analyzed together (Table 4). Among buried corals, bioerosion was higher at the high energy, low nutrient site (Paddoz Reef) than at the other two sites (ANOVA, F(2,23) = 4.58, p<0.02), but among sediment/water interface corals, there was no significant difference in bioerosion among sites. Taxonomic effects on bioerosion were weakly significant. Among buried corals, bioerosion was highest in massive corals, intermediate in branching corals, and lowest in free-living corals (ANOVA, F(2,30) = 5.03, p<0.015), but bioerosion was not significantly different among growth forms of sediment/water interface corals. Gaffarium generally experienced slightly greater bioerosion than Geloina , but these analyses were limited by the loss of all sediment-water interface Geloina specimens at Paddoz Reef. Species composition of internal borers.—Microboring dominated bivalve shells but was not clearly discernable in corals. Internal macroborings were dominantly unlined worm tubes (cf. Trypanites) or worm tubes with calcareous linings (Fig. 9). Sponge borings (Entobia sp.), formed by the endolithic sponge Cliona sp., were next most common, and were found predominantly in exposed specimens – covering extensive areas in bivalves. The endolithic bivalve traces Gastrochaenolites, formed by Lithophaga sp. and Gastrochaena sp., were least common, and occurred only in Goniastrea and Acropora corals at the most exposed site, Paddoz Reef, and did not occur in any of the other species. Bioerosion of bivalve shells was dominated by microborings and worms.,Boring sponges affected a considerable shell area among sediment/water interface shells at Gosem Island and Nagada Harbour, but not among buried shells. Average species richness of epibionts and epibionts was significantly higher among sediment-water interface corals and bivalves than among buried specimens (2-way ANOVA, F(5,29)=18.49, p=0.00025), but there were no statistically significant effects of reef or taxon on epibiont and endobiont diversity, norzere there significant interactions effects). Species composition of epibionts and endobionts was significantly different among treatments, but not among reefs(table 5). Samples left exposed at the sediment-water interface were all fairly similar in their epibiont/endobiont species composition, while the buried samples exhibited much greater variation (Fig 10A). Among experimental taxa, there were significant differences in species composition among endobionts, but not among epibionts (Table 5). When epibionts and endobionts were combined, significant differences were found between Paddoz Reef and Gosem Island, but not between other pairs of sites, and between Fungia and Gaffrarium, but not among other pairs of taxa. The MDS results showed no clear patterns of similarity among experimental taxa, as opposed to a clear difference in the degree of variation between exposed and buried treatments (Figure 10B). Best et al., Experimental taphonomy in Madang Lagoon, for PALAIOS DISCUSSION 9 The importance of burial and re-exhumation. The dominant pattern observed in this study was the importance of burial to limiting most forms of taphonomic alteration of corals and bivalves. Generally, buried samples experienced less loss, fragmentation, weight change, surface alteration, and infestation by epi and endobionts than sediment/water interface samples. Differences among field sites and among coral and bivalve forms were secondary, and were statistically weaker than the dominant effect of burial. A taxon effect between corals and bivalves mostly relates to their differential frequency of exhumation, and to the fact that the regular surfaces of bivalve shells are likely to be more fully encrusted than the irregular surfaces of coral skeletons. An apparent site effect of Paddoz related to the others is mostly due to the more frequent exhumation of buried specimens at that site. By contrast, the higher encrustation of buried bivalves than of buried corals at Nagada Harbour probably reflects preferential exhumation of bivalves at Nagada harbour, where the bivalves were buried on a slight slope. Loss, Fragmentation (one), disarticulation (by one year)…(Table 2) Weight change was highly variable with respect to site and form (Fig.3) and no clear pattern emerged because of high variability and low n. The general tendency was for sediment/water interface corals to lose more weight than their buried counterparts, and for bivalves to lose less weight or even gain weight due to encrustation. Even the shell fragment at Paddoz gained over what was the initial weight of the whole shell. Surface alteration varied very slightly with respect to site, even though wave energy was significantly different at the Paddoz site (Pandolfi and Minchin 1995), no consistent pattern was observed among corals and sediment/water interface bivalves (Fig.4,5). Buried bivalves showed adifference between sites, but higher alteration correlated with exhumation of buried specimens at Paddoz and Nagada. Discolouration also varied slightly with site, but differences among sites were only significant in buried specimens. Discolouration was lowest at the most exposed site (Paddoz Reef), and highest at the protected site (Nagada Harbour). High degrees of discolouration at Nagada Harbour are probably a result of organic-rich siliciclastic influence (Langer and Lipps, 2003) leading to surficial iron staining, probably with limonitic iron oxides (cf. Lescinsky et al. 2002). Burial plays a crucial role in determining the amount of encrustation and bioerosion experienced by bioclasts, since encrustation and bioerosion are limited or terminated by burial (Bromley and Asgaard, 1993; Chazottes et al., 1995; Perry, 1998; Lescinsky et al., 2002; Perry and Hepburn 2008; Best 2008). Differences in alteration among sites, and among host taxa, were primarily related to the frequency of exhumation. There was a difference between corals and bivalves in that the latter had more total surface area covered, which is likely due to the higher surface/volume ratio of the bivalves, and the smoothness of their surface. Wave energy at Paddoz actually resulted in lower total coverage of bivalves than at the other sites, with signs of erosion of the encrusted taxa. Also at Paddoz, buried samples were more similar to the sediment/water interface samples than at the other two reefs and reflects greater exhumation of buried samples. Note however exhumation of bivalves also occurred through slumping in the lowest energy site (Nagada), so it is not strictly correlated with wave energy, and in fact requires a setting where there is accumulation of sediments. Exhumation of buried shells is common in shallow marine environments, and tends to occur more frequently in shallower or more exposed environments than in deeper or more protected settings (e.g. Meldahl et al. 1997). In contrast to our results, exhumed brachiopod shells were apparently not encrusted or bored much, with nearly all encrustation occurring in the brief window between shell death and initial burial (Rodland et al. 2006). Best et al., Experimental taphonomy in Madang Lagoon, for PALAIOS 10 The fact that total bioerosion among buried corals was highest in massive Goniastrea, intermediate in branching Acropora, and lowest in disc-shaped free-living Fungia, without a similar pattern among sediment/water interface corals, probably results from differential susceptibility to exhumation among coral growth forms. The round, relatively smooth colonies of Goniastrea would likely move through sediment more readily than branching or highly irregular colony shapes found in Acropora or Fungia. The bivalve species were probably exhumed at equal frequencies, and presumably more readily than the corals, at Paddoz Reef and Nagada Harbour.. Where significant differences in intensity of encrustation and internal bioerosion occurred among reefs, if anything these parameters were highest at Paddoz Reef -PR-(lowest nutrient level, highest exposure level), and lowest at Nagada Harbour-NG-(highest nutrient level, lowest exposure level), although among the buried bivalve specimens the degree of encrustation and bioerosion were lowest at Gosem Island. Buried bivalves at Nagada Harbour apparently experienced a high degree of exhumation due to their deployment on a slope. The effect of wave energy (burial-exhumation cycles) largely overprinted any effect primary productivity and net sedimentation might have on the biological activity (Best and Kidwell, 2000a; Lescinsky et al., 2002). Species Composition of Epibionts and Endobionts on sediment/water interface and buried substrates.-Species composition of epibionts and endobionts reflected significant differences between sediment/water interface and buried treatments, some differentiation between coral growth forms and bivalve shell structures, but very little differentiation among sites. The significant difference in endobiont composition among forms may largely stem from an inability of endolithic bivalves to bore into bivalves substrates because of size limitations. As with encruster species composition, the endobiont species composition on Paddoz reef buried corals and shells was largely the same as that on the sediment/water interface shells from that reef. Again, this similarity reflects the importance of exhumation of buried corals and shells at Paddoz Reef. The degree of exhumation was apparently lowest at Gosem Island, and likely explains the The differences in species composition between sediment/water interface and buried corals and shells may also reflect the speed with which epibionts settle and grow on substrates. Worms, algae, and forams were common on both sediment/water interface and buried substrates, reflecting their rapid colonization abilities. Bryozoans, sponges, coral spat, vermetids, and bivalves were all more common on sediment/water interface than on buried substrates, reflecting the greater time window required for settlement and growth in these organisms. Similarly, borers were dominated by microborings and worms and sponges, which rapidly colonize substrates, while bivalves were less common, as found in bioerosion experiments (Kiene and Hutchings 1994; Chazottes et al. 1995; see Edinger 2000 for review). The relatively minor role of bivalves in bioerosion is partially a consequence of the short duration of the experiment. Differences in species composition of epibionts might help to indicate the rapidity with which fossil bioclasts were buried, or the duration for which they were exposed during burial-exhumation cycles. Unless separated from the effects of burial, differences in species composition of epibionts are unlikely to yield information on paleoproductivity (Lescinsky et al. 2002; Tapanila et al. 2004). Environments shielded from the effects of burial, such as hardgrounds and rocky shores, are most likely to yield useful paleoproductivity data through encruster and bioeroder abundance and species composition (Johnson. These non-depositional environments have low preservation potential, and may still be affected by differences in length of exposure before sedimentation resumes after facies change. Best et al., Experimental taphonomy in Madang Lagoon, for PALAIOS 11 The effect of form on endobiont species richness and composition was more limited. The massive coral Goniastrea had more macroborers than the other two coral growth forms, particularly more sponges and bivalves. This pattern likely results from higher skeletal density than the other coral growth forms (cf. Highsmith 1983; Tapanila et al. 2004), but also from the fact that the round shape of Goniastrea skeletons, and their possible longer exposure may render them a more suitable substrate for boring bivalves (cf. Risk et al. 1995). Among the bivalves, larger Geloina coaxans skeletons, with high organic content, had higher rates of shell loss and disarticulation than low organic, smaller Gafrarium tumidum. These differences may relate to its organic matrix content. By contrast, Gaffrarium had consistently higher rates of encrustation, among both sediment-water inferface and buried shells,than Geoloina.. Implications for the nature of time-averaging on coral reefs. Time-averaging is a fundamental stratigraphic process affecting the temporal resolution of the fossil record in all depositional environments (Kowalewski, 1996). Time-averaging can be divided into sedimentological and ecological time-averaging. Sedimentological time-averaging results from vertical mixing of bioclasts throughout the taphonomically active zone, resulting in an exponentially young-skewed) age distribution within a single stratigraphic horizon (cf. Kidwell and Flessa, 1995; Meldahl et al., 1997). By contrast, ecological time-averaging results from co-occurrence of organisms with radically different life-spans, such that a single stratigraphic horizon contains a wide-spectrum of bioclast ages, often but not necessarily with a young-skewed age distribution (Kowalewski 1996). These patterns to date have been observed predominantly in bivalves. Wide ranges in coral lifespans, combined with broad differences in coral carbonate production rates, can lead to large discrepancies between the relative abundance of coral growth forms in coral reef life assemblages, death assemblages, and fossil assemblages (James and Bourque, 1992; Zuchsin, 2000; Edinger et al., 2001). Reef crests are generally thought to be environments without abundant unconsolidated sediment, where ecological time-averaging predominates over sedimentological time-averaging (Edinger et al., 2001; Edinger et al., 2007). However reefal deposits have shown evidence of high degrees of time averaging due to local slumping and wave energy (Kidwell et al 2005, Best 2008). Exhumation of corals and molluscs on the lagoonal reefs in this study necessarily reflects vertical bioclast movement through the sediment, or sedimentological time-averaging, (cf. Kowalewski, 1996; Meldahl et al., 1997). There is also evidence for differential movement between taxa, with the massive coral Goniastrea apparently experiencing more frequent exhumation than the branching or free-living corals, and with the smaller lighter bivalves showing more evidence of burial/exhumation cycles than the corals. The high frequency of exhumation of both corals and bivalves on all three sites suggests that natural burial-exhumation cycles are commonplace in these shallow reef lagoon settings. The exhumation of buried corals and molluscs attests to the importance of sedimentological time-averaging in shallow reef lagoon facies. Fore-reef slopes may also be prone to sedimentological time-averaging due to off-reef transport and slumping (James and Bourque 1992). The dominance of ecological time-averaging is therefore likely limited to reef facies without abundant loose sediment, such as reef front and crests. Within the broad spectrum of reef facies, these are volumetrically the minority, therefore sedimentological time-averaging is probably the norm for much of the reef record. SIGNIFICANCE FOR THE FOSSIL RECORD This study corroborates the primary importance of burial in preservation quality of skeletal remains Best et al., Experimental taphonomy in Madang Lagoon, for PALAIOS 12 (e.g. Lazo 2004, Best et al, 2007, Best 2008). As such, secondary influences on fossil preservation such as wave exposure and paleoproducitivity are likely to be generally obscured, except in those cases where limited sediment supply prevents rapid burial, or where exhumation of buried skeletons is common. Use of epibiont and endobiont abundance, diversity, and species composition to aid in fossil reef and peri-reefal carbonate facies intepretation will likely be most successful along gradients of wave energy as moderated by exhumation of buried bioclasts. Large biovolume encrusters and macroborers indicative of longer duration of exposure will likely be more useful as indicators of exposure than small encrusters and micrborers, which can colonize exposed bioclasts more rapidly (cf. Bromely and Asgaard, 1993; Lescinksy et al. 2002). The study also emphasizes the rapidity with which alteration of fossil remains takes place. Early checking of specimens after 1.5 months of deployment showed that modification began immediately (Best et al., 2004). Exquisitely preserved fossils not only require rapid burial, but also require a lack of exhumation associated with wave energy, sediment mass-movement, or biological mixing of sediments (e.g. Best 2008). Our results also highlight the importance of re-exhumation. This has also been reported in siliciclastic settings, and can lead to the strongly skewed age distributions of shells exposed at the sediment-water interface (e.g. Meldahl et al., 1997; Kowalewski et al. 1998; Carroll et al. 2003; Kidwell et al. 2005). While it was not surprising that exhumation of buried corals and shells was highest at the site with the greatest wave exposure, the patterns of coral and shell exhumation, associated not only with wave energy gradients but also slumping, are likely to overwhelm onshore-offshore trends in sedimentation and nutrient availability (cf. Lescinsky et al. 2002). Thus patterns of paleoproductivity may be obscured by patterns of wave energy, except where productivity is driven exclusively by upwelling, independent of gradients in sedimentation and wave exposure. Exhumation at Nagada Harbour may have been moderately important, particularly for bivalves; buried specimens at Nagada Harbour were deployed on a sloping muddy-sand bottom, which likely experienced some slumping during the course of the experiment. Again this emphasizes that bottom steepness, not only wave energy, has a strong effect on sediment movement, and therefore skeletal transport and burial (e.g. Kidwell et al. 2001). Reefs in nearshore areas with high sedimentation rates are likely to experience high quality of preservation primarily due to the rapidity with which corals are buried, and the infrequency with which they are exhumed (cf. Best and Kidwell 2000a; Perry and Smithers 2006; Best 2008) The high incidence of experimental coral and shell loss at the exposed site, Paddoz Reef, emphasize the importance of bioclast transport, and not just burial and re-exhumation. We observed this differential transport mostly strongly between corals and bivalves, and to a lesser extent among the coral growth forms. Buried corals and shells are likely to be moved if exhumed by sediment mass-moved events induced by storms or slumping. While reef crest deposits are generally assumed to be largely untransported, reef lagoon deposits are likely to reflect some degree of in habitat transport, with differential transport among different clast size (cf. James and Bourque 1992). Diffierences in the total surfaces infested by encrustors and borers follows differences in burial exhumation (clast size), surface/volume ratios, and surface smoothness. Differences in encrustor and borer species composition between corals and bivalves were relatively minor, and were consistently less important than the effect of burial. The similarity in degree and type of taphonomic alteration driven by burial/exhumation emphasizes the high quality of the fossil record, and the extent to which ecological patterns of biodiversity in the reef fossil record are not strongly biased by differential preservation along lines of morphology, shell microstructure or composition (cf. Best and Kidwell 2000 Best et al., Experimental taphonomy in Madang Lagoon, for PALAIOS a&b) 13 CONCLUSIONS (1) The specimens that were deployed at the sediment/water interface or buried and exhumed generally showed a higher degree of taphonomic alteration than those that remained burid., Exceptions are higher discolouration in buried specimens, and encrustation cover for bivalves. The impact of burial/exhumation was much stronger than differences in morphology, microstructure, or form. (2) The effect of varying exposure at the sediment-water interface overprints the influence of nutrients and primary productivity on skeletal alteration (Lescinsky et al., 2002; Best 2008). (4) In this study, differences among coral growth forms played a relatively unimportant role in taphonomic alteration, only having minor effects on bioerosion and surface alteration. Similar minor differences show for organic-rich bivalve shells which degraded more rapidly than organic-poor shells. The taxonomic difference was between corals and bivalves, where differences in size, surface/volume ratios, and surface smoothness did result in significant differences. (5) Encrustation was the predominant form of biological alteration. Initial colonization by macroborers is usually slow, and might become more important over a longer period of time. (6) Taphonomic alteration of corals and bivalves on reefs proceeds rapidly (within 1 month), with measurable alteration of bioclasts occurring within one year. ACKNOWLEDGEMENTS This research was conducted during a Christensen Research Fellowship awarded to JMP and was also supported by grant no. AD9031784 to JMP from the Australian Research Council. We are grateful to V. Baker, C.E. Lovelock, and R. Whelan for diving assistance. M. Jebb and the personnel of the Christensen Research Institute provided an introduction to the environments of Madang Lagoon and assisted in all logistical aspects of the fieldwork. MB thanks P. Burniaux for the bivalve data collection, and AIMS for the student visiting fellowship. EE thanks University of California Davis for a student travel grant that enabled his participation in the field work. Further support was provided through the ARC Centre of Excellence for Coral Reef Studies (http://www.coralcoe.org.au/index.html). REFERENCES Behrensmeyer, A.K.and S. M. Kidwell.1985. Taphonomy’s contributions to paleobiology. Paleobiology, 11:105-119. Best et al., Experimental taphonomy in Madang Lagoon, for PALAIOS 14 Best, M.M.R. 2008. Contrast in preservation of bivalve death assemblages in siliciclastic and carbonate tropical shelf settings. Palaios 23: 796–809. Best, M.M.R and S.M. Kidwell. 2000a. Bivalve taphonomy in tropical mixed siliciclastic-carbonate settings: I. Environmental variation in shell condition. Paleobiology, 26: 80-102. Best, M.M.R. and S.M. Kidwell, 2000b. Bivalve taphonomy in tropical mixed siliciclastic-carbonate settings: II. Effect of bivalve life habits and shell shapes. Paleobiology 26: 103-115. Best, M.M.R., P. Burniaux, and J.M. Pandolfi, 2004. Experimental bivalve taphonomy in reefs of Madang Lagoon, Papua New Guinea. In, M.M.R. Best, and J-B. Caron, eds., Proceedings, Canadian Paleontology Conference, Proceedings #2, p. 8-12. Best, M.M.R., T.C.W. Ku, S.M. Kidwell, and L.M. Walter, 2007. Carbonate preservation in shallow marine environments; unexpected role of tropical siliciclastics. Journal of Geology 115: 437-456. Brenchley, P. J and D. A. T. Harper. 1998. Palaecology: Ecosystems, environments and evolution. Hall, Thomson Science, UK, Ch. 3. Chapman & Brett, C. E. and G. C. Baird. 1986. Comparative Taphonomy: A Key to Paleoenvironmental Intrepretation Based on Fossil Preservation. Palaios, 1: 207-227. Briggs, D.E.G, 2000. Exceptionall preserved fossils. .—In: Briggs, D.E.G., and Growther, P.R. (eds.): Palaeobiology II.—328–332, Oxford, (Blackwell). Bromley, R. G. and U. Asgaard. 1993. Two bioerosion ichnofacies produced by early and late burial associated with sea-level change. Geol Rundsch, 82: 276-280. Chazottes, V., T. Le Campion-Alsumard and M. Peyrot-Clausade. 1995. Bioerosion rates on coral reefs: interactions between macroborers, microborers and grazers (Moorea, French Polynesia). Palaeogeography, Palaeoclimatology, Palaeoecology, 113: 189-198. Clarke & Warwick 1993. Edinger, E.N., 2001. Bioerosion.—In: Briggs, D.E.G., and Growther, P.R. (eds.): Palaeobiology II.—273–277, Oxford, (Blackwell). Edinger, E. N., and M. J. Risk. 1997. Sponge borehole size as a relative measure of bioerosion and paleoproductivity. Lethaia, 29: 275-286. Edinger, E.N., J.M. Pandolfi, and R.A. Kelley, 2001. Community structure of Quaternary reefs compared to Recent life and death assemblages. Paleobiology 26(4):669-694. Edinger, E.N., G.S. Burr, J.M. Pandolfi, and J.C. Ortiz, 2007. Age accuracy and resolution of Quaternary corals used as proxies for sea level. Earth and Planetary Science Letters 253: 37-49. Estrada Alvarez, L.M., E.N. Edinger, and J.M. Pandolfi, 2004. Taphonomy of modern corals from Madang Lagoon, Papua New Guinea. In, M.M.R. Best, and J-B. Caron, eds., Proceedings, Canadian Paleontology Conference, Proceedings #2, p. 23-28. Greenstein, B. J. and H. A. Moffat. 1996. Comparative Taphonomy of Modern and Pleistocene Corals, San Salvador, Bahamas. Palaios, 11: 57-63. Greenstein and Pandolfi 1997. Best et al., Experimental taphonomy in Madang Lagoon, for PALAIOS 15 Greenstein, B.J., and J.M. Pandolfi, 2003. Taphonomic alteration of reef corals: effects of reef environment and coral growth form. II: the Florida Keys. Palaios 18: 495-509. Highsmith 1983. Holmes, K.E., E.N. Edinger, Hariyadi, G.V. Limmon, and M.J. Risk, 2000. Bioerosion of live massive corals and branching coral rubble on Indonesian coral reefs. Marine Pollution Bulletin 40: 606-617. James and Bourque 1992 Jebb, M., 1991. The Madang Lagoon: A natural history (in ms.). Kidwell, S.M., M.M.R. Best, and D.S. Kaufman, 2005. Taphonomic trade-offs in tropical marine death assemblages: differential time averaging, shell loss, and probable bias in siliciclastic vs. carbonate facies. Geology 33: 729-732. Kidwell, S.M., 2005. Shell composition has no net impact on large-scale evolutionary patterns in molluscs. Science 307: 914-917. Kidwell, S.M., 2000. Major biases in the fossil record. .—In: Briggs, D.E.G., and Growther, P.R. (eds.): Palaeobiology II.—297-303, Oxford, (Blackwell). Kidwell, S.M. and K.W. Flessa, 1995. The quality of the fossil record; populations, species, and communities. Annual Review of Earth and Planetary Sciences 24: 433-464. Kiene, W.E. and P.A. Hutchings, 1994. Bioerosion experiments at Lizard Island, Great Barrier Reef. Coral Reefs 13: 91-98. Kowalewski, M., 1996. Time-averaging, overcompleteness, and the geological record. Journal of Geology 104: 317-326. Kowalewski, M., G.A. Goodfriend, and K.W. Flessa 1998. High-resolution estimates of temporal mixing within shell beds; the evils and virtues of time-averaging. Paleobiology 24: 287-304. Langer, M. R. and J. H. Lipps. 2003. Foraminiferal distribution and diversity, Madang Reef and Lagoon, Papua New Guinea. Coral Reefs, 22: 143-154. Lazo, D.G., 2004. Bivalve taphonmy: testing the effect of life habits on the shell condition of the Littleneck Clam Protothaca (Protothaca) staminea (Mollusca: Bivalvia). Palaios 19: 451-459. Lescinsky, H. L., E. N. Edinger, and M. J. Risk. 2002. Mollusc Shell Encrustation and Bioerosion Rates in a Modern Epeiric Sea: Taphonomy Experiments in the Java Sea, Indonesia. Palaios, 17: 171-191. Meldahl, K.H., K.W. Flessa, and A.H. Cutler, 1997. Time-averageing and post-mortem skeletal survival in benthic fossil assemblages: quantitative comparisons among Holocene environments. Paleobiology 23: 207-229. Pandolfi, J. M., 1996. Limited membership in Pleistocene reef coral assemblages from the Huon Peninsula, Papua New Guinea: constancy during global change. Paleobiology, 22: 152-176. Pandolfi, J. M. 2001. Ch. 3, 3.3.3 Ancient Reefs in: Briggs, D. E. G. and P. R. Crowther, eds. 2001. Palaeobiology: II. Blackwell Scientific Publications, UK, pp 307-309. Pandolfi, J. M., and B. J. Greenstein. 1997a. Taphonomic alteration of reef corals: effects of reef environment and coral growth form. I: The Great Barrier Reef. Palaios, 12: 27-42. Pandolfi, J. M., and B. J. Greenstein. 1997b. Preservation of community structure in death assemblages of deep-water Caribbean reef corals. Limnology and Oceanography, 42: 1505-1516. Best et al., Experimental taphonomy in Madang Lagoon, for PALAIOS 16 Pandolfi, J. M., and P.R. Minchin. 1995. A comparison of taxonomic composition and diversity between reef coral life and death assemblages in Madang Lagoon, Papua New Guinea. Palaeogeography, Palaeoclimatology, Palaeoecology, 119: 321-341. Perry, C.T. 1998. Macroborers within coral framework at Discovery Bay, north Jamaica: species distribution and abundance, and effects on coral preservation. Coral Reefs, 17: 277-287. Perry, C.T. and S.G. Smithers, 2006. Taphonomic signatures of turbid-zone reef development: examples from Paluma Shoals and Lugger Shoal, inshore central Great Barrier Reef, Australia. Palaeogeography, Palaeoclimatology, Palaeoecology, 242: 1-20. Perry, C.T., and L.J. Hepburn, 2008. Syn-depositional alteration of coral reef framework through bioerosion, encrustation and cementation: taphonomic signatures of reef accretion and reef depositional events. Earth Science Reviews 86: 106-144. Scoffin, T.P., 1992. Taphonomy of coral reefs: a review. Coral Reefs 11: 57-77. Tapanila, L.M., P. Copper, and E.N. Edinger, 2004. Environmental and substrate control on Paleozoic bioerosion in corals and stromatoporoids, Anticosti Island, eastern Canada. Palaios 19(3): 292-306. Zuchsin, M., J. Hohenegger, and F. F. Steininger. 2000. A comparison of living and dead molluscs on coral reef associated hard substrata in the northern Red Sea—implications for the fossil record. Palaeogeography, Palaeoclimatology, Palaeoecology 159:167–190.. 17 Best et al., Experimental taphonomy in Madang Lagoon, for PALAIOS FIGURE CAPTIONS. Figure 1: Madang Lagoon and location of field sites along onshore-offshore gradient. Nagada Harbour, Middle site: Gozem Island, Offshore site, Paddoz Reef. Figure 2: Pre-experimental photographs of the experimental taxa. Inshore site: A: Acropora pulchra; B: Goniastrea retiformis; C: Fungia sp., (top) ;; D: Fungia sp. (bottom) E: i) Gaffrarium tumidum exterior, ii) Gaffrarium tumidum interior; F: i) Geloina coaxans exterior, ii) Geloina coaxans interior. Scale bars in figs A-D: 5 cm. Scale bars in E-F: 2 cm. Figure 4. Alteration of coral and bivalve skeletons. A: Acropora, buried, Nagada Harbour. B: Acropora, sediment/water interface, Nagada Harbour, showing loss of surface detail. C: Fungia, initially buried, Paddoz Reef; encrusted by serpulid worm and therefore presumably exhumed during the deployment period. D: Fungia, sediment/water interface, Paddoz Reef; encrusted by bryozoan (b), calcareous red algae (c) and miliolid foram (m). Linear features running diagonally though photo are coral septae. E: Goniastrea, buried, Paddoz Reef; encrusted by red foraminiferan Homotrema rubrum and therefore presumably exhumed during the deployment period . F: Goniastrea, sediment/water interface, Paddoz Reef; bored by Lithophaga sp. bivalve. G: Gaffrarium tumidum, buried, Gozem Island; H: G. tumidum, sediment/water interface, Nagada Harbour, with encrusting sponge (left valve) and extensive clionid sponge boring (right valve). I: Geloina coaxans, buried, Nagada Harbour; J: G. coaxans, sediment/water interface, Nagada Harbour, with extensive encrustation and discolouration. Figure 5. Degree of surface alteration of corals and mollusk valves. A: sediment-water interface; B: buried. Not discerable refers to cases where extensive encrustation prevented assessment of alteration of the original surface. Figure 3. Percent weight change observed in corals (Acropora, Goniastrea, Fungia) and bivalves (Gaffrarium, Geloina) at the three sites, and under the two experimental treatments. Points without error bars indicate cases with less than 3 replicates recovered. High variance in results for Gaffrarium at Paddoz Reef also reflects loss of replicates. Weight gain reflects results of encrustation by skeletal epibionts. Figure 6. Total encrustation of buried and sediment/water interface corals and bivalves in each site. Percent cover of encrusters differed significantly between sediment/water interface and buried treatments, and among reefs for buried only. Figure 7. Average species richness and frequency of occurrence of encrustor taxa on sediment/water interface and buried corals and mollusk valves. Frequency of occurrence calculated as the average occurrence of each encrustor on corals or mollusk valves in each replicate; e.g. 1 of 4 Goniastrea Best et al., Experimental taphonomy in Madang Lagoon, for PALAIOS replicates among the buried corals at Paddoz reef were encrusted by corals. each replicate indicated under each bar. 18 N of corals or valves in Figure 8. Total bioerosion in corals and bivalves, measured as the total number of boreholes per substrate. Mean 95% c.i. Figure 9. Average species composition of bioeroding organisms on sediment/water interface and buried corals and bivalves. Figure 10A. 2-D MDS ordination of epibiont and endobiont species composition at the presence-absence level, comparing treatment and reefs. The dominant effect on species composition was exposure vs. burial, while there was little difference in encrustor species composition between sites.. Figure 10B. 2-D MDS ordination of epibiont and endobiont species composition at the presence-absence level, comparing treatment and experimental taxa. Again, the dominant effect on epibiont and endobiont species composition was exposure vs. burial while experimental taxon had little effect. TABLE CAPTIONS. Table 1. Experimental site locations and characteristics along onshore-offshore gradient. Inshore site: Nagada Harbour, Middle site: Gozem Island, Offshore site, Paddoz Reef. Table 2. specimen recovery, fragmentation, disarticulation summary table. Best et al., Experimental taphonomy in Madang Lagoon, for PALAIOS 19 Table 3. Total Encrustation and macroboring 2-way ANOVA results. Table 4. Analysis of similarity (ANOSIM) results comparing species composition of epibionts and endobionts among experimental treatments, reefs, and taxa,: Bold indicates significant at α=95%. All from Bray-Curtis similarity calculated at the presence-absence level.