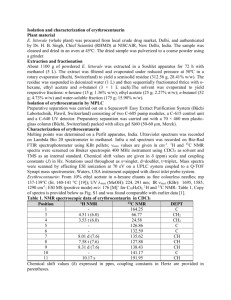
Materials and Methods:
advertisement

Solution NMR structure and dynamics of human apo-S100A1 protein
Michał Nowakowski a, Łukasz Jaremko a,b, Mariusz Jaremko a, Igor Zhukov a,c,d, Agnieszka
Belczyk a, Andrzej Bierzyński a, and Andrzej Ejchart a,*
a
Institute of Biochemistry and Biophysics, Polish Academy of Sciences, Pawińskiego 5A, 02-
106 Warsaw, Poland
b
Faculty of Chemistry, Warsaw University, Pasteura 1, 02-093, Warsaw, Poland
c
Slovenian NMR Centre, National Institute of Chemistry, Hajdrihova 19, P.O.B. 660, SI-
1001 Ljubljana, Slovenia
d
EN-FIST Center of Excellence, Dunajska 156, SI-1000, Ljubljana, Slovenia
* Corresponding author. Phone: +48 22 5922037. Fax: +48 22 5922038.
E-mail addresses: lyam@ibb.waw.pl (M. Nowakowski), jaremko@gmail.com (Ł. Jaremko),
antypater@gmail.com (M. Jaremko), igor@ibb.waw.pl (I. Zhukov), agibel@ibb.waw.pl (A.
Belczyk), ajb@ibb.waw.pl (A. Bierzyński), aejchart@ibb.waw.pl (A. Ejchart).
1
Abstract
S100A1 belongs to the EF-hand superfamily of calcium binding proteins. It is a
representative of the S100 protein family based on amino acid sequence, three-dimensional
structure, and biological function as a calcium signal transmitter. It is a homodimer of
noncovalently bound subunits. S100A1, like most of other members of the S100 protein
family, is a multifunctional, regulatory protein involved in a large variety of biological
processes and closely associated with several human diseases. The three-dimensional
structure of human apo-(i.e. calcium free)-S100A1 protein was determined by NMR
spectroscopy (PDB 2L0P) and its backbone dynamics established by 15N magnetic relaxation.
Comparison of these results with the structure and backbone dynamics previously determined
for bovine apo-S100A1 protein modified by disulfide formation with -mercaptoethanol at
Cys 85 revealed that the secondary structure of both these proteins was almost identical,
whereas the global structure of the latter was much more mobile than that of human apo-S100
protein. Differences between the structures of human and rat apo-S100A1 are also discussed.
Keywords: calcium binding protein, human apo-S100A1, NMR, backbone dynamics, protein
structure
Abbreviations: CPMG, Carr – Purcell – Meiboom – Gill; DSS-d4, 3-trimethylsilyl-2,2,3,3tetradeuteropropionic acid sodium salt; EDTA, ethylenediaminetetraacetic acid; HSQC,
heteronuclear single quantum coherence; HPLC, high performance liquid chromatography;
NMR, nuclear magnetic resonance; NOE, nuclear Overhauser effect; S100A1, homodimeric
S100A1 with noncovalent interactions at the dimer interface; S100A1-ME, mixed disulfide
of S100A1 protein with β-mercaptoethanol; TRIS-d11, perdeuterated 2-amino-2(hydroxymethyl)1,3-propanediol.
2
1. Introduction
S100 proteins play a crucial role in many biological processes regulated by calcium in
the cells of vertebrate species (Santamaria-Kisiel et al., 2006). S100A1 protein can be treated
as an ideal representative of the S100 protein family because of its amino acid sequence,
three-dimensional structure, and biological function as a calcium signal transmitter.
It is a homodimer of noncovalently bound subunits. Each of which contains two EFhand structural motifs formed by two -helices flanking a calcium-binding loop. These motifs
are covalently connected through a short linker and noncovalently by interhelical hydrophobic
interactions and a hydrogen bond(s) between the two Ca2+-binding loops that stabilizes a
short, antiparallel -sheet. Such a structure, determined for rat S100A1 protein using NMR
methods (Rustandi et al., 2002; Wright et al., 2005), closely corresponds to other known
structures of S100 proteins (Marenholtz et al., 2004).
S100A1, like other members of the S100 protein family (with rare exceptions) is a
multifunctional, regulatory protein involved in a large variety of biological processes. It is
closely associated with several human diseases including cardiomyopathy, neurodegenerative
disorders, and cancer (Heizmann et al., 2004).
Recently, we have shown that chemical modifications of the unique SH group within
the protein (that of Cys 85) by the covalent S-S bonding to small thiols, such as glutathione,
cysteine or even -mercaptoethanol, results in an enormous increase (a couple of orders of
magnitude) of the protein affinity for calcium (Goch et al., 2005). Therefore, it is highly
probable that the protein can act as a linker between the redox- and calcium-signal pathways
that, in a number of biological processes, were proven to be coupled with each other (DalleDonne et al., 2008), although very little is known about the molecular mechanism of this
coupling.
3
It seems, therefore, to be interesting to find out how the S100A1 structure is changed
by chemical modifications of its Cys 85 residue. We tried to answer this question by
determining the structure of bovine apo-S100A1 modified by -mercaptoethanol and
comparing it with the only structure of apo-S100A1 determined so far: that of rat protein
(Rustandi et al., 2002). The results of that work were presented and discussed in our previous
paper (Zhukov et al., 2008). Nevertheless, we were aware that the conclusions presented in it
were perhaps arguable. The observed structural differences between the both proteins were
small. They could arise, at least in part, from the difference of their sequences or, even worse,
they simply could be an artifact, because experimental procedures and data analysis methods
used to determine these structures had not been the same.
Therefore, we decided to determine and compare the structures of the same S100A1
molecule, native and modified at Cys 85 in its apo and holo states. Human S100A1 protein
has been chosen as the object of our studies and in this paper its structure and backbone
dynamics in the apo state are presented.
2. Materials and methods
2.1. Sample preparation
15
N-labeled and 13C,15N-double labeled S100A1 protein was obtained as previously
described (Bolewska et al., 1997). The synthetic gene encoding for human S100A1 was
cloned into pET-30a+ plasmid and expressed in E. coli utilizing the T7 expression system.
The bacteria were grown in M9 media containing (15NH4)2SO4 as the sole nitrogen source and
either unlabeled or 13C labeled glucose as the sole carbon source. Expression products were
isolated using the classical method of ammonium sulfate precipitation (Dixon and Webb,
1961; Falconer et al., 1953), purified by reversed-phase HPLC on a semipreparative Vydac
C18 column, and identified by electrospray ionization mass spectrometry using a Macromass
4
Q-Tof spectrometer. Two forms of the protein, one with the sequence strictly corresponding
to its gene sequence and another one, with the additional initiator methionine at the Nterminus, were obtained from HPLC as partly overlapping peaks. NMR measurements
indicated that structural differences between both forms are small and restricted to the close
proximity of the N-terminal Met residue (Baldisseri et al., 1999). Therefore, a mixture of both
forms of proteins was used in all NMR measurements. 650 L samples contained 1 mM
protein solution (monomer concentration) in 90%/10% H2O/D2O, 50 mM TRIS-d11, 1mM
EDTA, 0.1mM NaN3 and 50 mM NaCl with pH adjusted to 6.8 (uncorrected value). In case
of HC detected experiments 100% deuterated buffer was used.
2.2. NMR spectroscopy
All NMR measurements were performed at the temperature carefully adjusted to 37°C
checked by an ethylene glycol reference sample. Time domain data were acquired using the
States-TPPI quadrature detection (Marion et al., 1989) followed by the sensitivity enhanced
detection introduced by Kay et al. (1992a). 1.6 s recycling delay was used, if not stated
otherwise. All chemical shifts in 1H NMR spectra were reported with respect to external DSSd4. Chemical shifts of 13C and 15N signals were referenced indirectly using the 0.251449530
and 0.101329118 frequency ratios for 13C/1H and 15N/1H, respectively (Wishart et al., 1995).
Experimental data were processed using the NMRPipe software package (Delaglio et al.,
1995). Zero filling and 90° shifted squared sine-bell filter were performed prior to the Fourier
transformation. Processed spectra were analyzed with both the CARA (Keller, 2004) and the
SPARKY (Goddard and Kneller) software.
The sequence-specific assignment was performed using a uniformly 13C,15N-double
labeled sample of apo-S100A1. 3D heteronuclear HNCO (Muhandiram and Kay, 1994),
HNCA (Ikura et al., 1990), HN(CO)CA (Ikura et al., 1991), HNCACB (Wittekind and
Mueller, 1993), CBCA(CO)NH (Grzesiek and Bax, 1992) and (HCA)CO(CA)NH (Lohr and
5
Ruterjans, 1995) spectra were used to obtain assignments of the backbone 1H, 13C and 15N
resonances. The assignment was additionally confirmed by analysis of sequential and
medium-range NOEs in the 3D 15N-edited NOESY-HSQC experiment (Zhang et al., 1994).
Aliphatic side chain 1H and 13C resonances were assigned from the analysis of the 1H–13C
HSQC, C(CO)NH (Grzesiek et al., 1993), HBHA(CBCACO)NH (Grzesiek and Bax, 1993),
HCCH-TOCSY (Bax et al., 1990) and 13C-edited NOESY-HSQC spectra (Muhandiram et al.,
1993). Aromatic side chain resonances were fully assigned from the (HB)CBHD, (HB)CBHE
(Yamazaki et al., 1993), 1H–13C HSQC and the 13C-edited NOESY-HSQC spectra recorded
with the offset, spectral widths and 13C–1H coupling constants tuned to aromatic carbons.
Distance constraints were obtained from the 15N-edited NOESY-HSQC spectrum and the 13Cedited NOESY-HSQC spectra separately tuned to aliphatic and aromatic carbons. The
detailed information about all experiments is summarized in Supplementary Table A.1.
Relaxation measurements were performed at three magnetic fields: 9.4, 11.7 and 16.4
T, with 15N-labeled sample. Longitudinal (R1) and transverse (R2) relaxation rates were
measured using the sensitivity enhanced 1H–15N HSQC pulse sequence (Kay et al., 1992a)
with the option of either R1 or R2 measurements of 15N nuclei (Farrow et al., 1994). The R2
relaxation rate measurements were performed with the CPMG pulse train. A refocusing time
of 650 s was used during the evolution delays. The acquisition parameters for R1 and R2
measurements on each spectrometer were identical with the exception of the delay between
(1H) pulses used for the cross-correlation effect suppression (Kay et al., 1992b). 5 and 10 ms
delays were used in the R1 and R2 measurements, respectively. Delays between the scans of
1.8 s were employed in both types of experiments. {1H}–15N NOEs were measured with the
pulse sequence included in the ProteinPACK Varian Inc. (Palo Alto, CA) software. Details
and experimental parameters of relaxation spectra are gathered in Supplementary Table A.2.
6
Resonance intensities were used in calculating relaxation rates and NOE values. All
experiments were repeated twice. Experimental errors of relaxation rates were obtained from
appropriate elements of the variance-covariance matrix. Experimental errors in NOE values
were evaluated based on signal-to-noise ratios obtained for corresponding signals in spectra
with and without NOE (Fushman, 2003). Consistency of measurements performed at different
magnetic field strengths was confirmed as proposed by Morin and Gagne (2009).
The 1H, 13C and 15N resonance assignments and 15N magnetic relaxation data have
been deposited in the BioMagResBank with the accession code 16360.
2.3. Structure calculation
The NOESY interproton distance constraints were derived from the 3D heteronuclear
15
N- and 13C-edited NOESY-HSQC experiments. The initial structure calculations were
performed with the CYANA 3.0 software (Guentert et al., 1997). The dimeric interface
between the protein subunits linked by a chain of dummy atoms, was defined basing on
structural similarity to the known structure of rat S100A1 protein (PDB entry 1K2H). The
automatic NOESY assignment procedure (Herrmann et al., 2002) provided 754 short, 434
medium and 272 long range intrasubunit distance constraints and 132 intersubunit distance
constrains, defining their mutual orientation. Additionally, 134 restraints for backbone φ and
ψ torsion angles for Ser 2–His 18, Leu 28–Glu 63, and Val 69–Thr 82 were predicted from the
chemical shifts using the PREDITOR server (Berjanskii et al., 2006). For the less defined
protein regions (like Ser 19–Lys 27 and Asn 64–Glu 68 calcium binding loop fragments or the
C-terminus of helix IV starting from Val 83) the dihedral angle constraints were not applied.
Stereospecific assignments for 122 side chain chiral groups in each subunit were generated by
the program GLOMSA (Guentert et al., 1991) that is included in the CYANA software.
Before the final structure calculations, 184 constraints defined as rHN-O = 1.5–2.5 Å and rN-O =
2.5–3.5 Å for 92 hydrogen bonds were added based on geometric criteria. If a given hydrogen
7
bond existed in more than 75% of structures in the ensemble, then it was selected for the final
refinement. The final structure refinement was performed with the XPLOR-NIH 2.2.5
program (Schwieters et al., 2003) using the symmetry equivalence constraints between two
subunits. Evaluation of the obtained human apo-S100A1 structure quality was done with the
PROCHECK-NMR (Laskowski et al., 1996) and the What-If (Vriend, 1990) programs.
2.4. Analysis of 15N relaxation data
Methodology for analyzing 15N relaxation data measured at multiple magnetic fields
has been described in our previous papers (Zhukov et al., 2007; Zhukov et al., 2008). There
are, however, two differences between the method utilized earlier and that used in this paper.
First, the extended model-free approach (Clore et al., 1990) has been applied rather
than the genuine one (Lipari and Szabo, 1982). The original model often fails to represent
experimental data accurately, especially the calculated NOE values tend to be too small. The
extended model includes five parameters; rotational diffusion correlation time R, two order
parameters, Sf and Ss, and two effective correlation times, f and s, associated with fast and
slow internal motions. One can expect that local motions of residues located in calcium
binding loops and a linker will be more adequately described with two internal motions taking
place on significantly different time scales.
Second, the approximation of the axially symmetric overall motion has been replaced
with a fully asymmetric one. It has been checked with Fisher-Snedecor statistics (F test) that
the fully asymmetric model is significantly better at the probability 0.01 (details of the model
selection are given in Supplementary Table A.3). Appropriate spectral density functions can
be derived (Baber et al., 2001, Tjandra et al., 1995) combining the extended model-free
approach with anisotropic overall tumbling (Woessner, 1962).
Three Euler angles relate the directions of the principal axes of the asymmetric
diffusion tensor to the molecule fixed coordinate system. Because of the symmetry
8
requirement one of the principal axes of the diffusion tensor of homodimeric S100A1 protein
is collinear with the protein symmetry axis. This condition reduces the number of Euler
angles from three to one. The coefficients describing orientation of H–N vectors in to the
molecule fixed coordinate system have been calculated using the atomic coordinates of the
NMR-derived structure. The vibrationally averaged N–H distance was assumed to be 0.104
nm (Ottiger and Bax, 1998) and chemical shift anisotropy of axially symmetric 15N chemical
shift tensor was taken to be –170 ppm.
The least-squares procedure used to optimize the model parameters consisted of
minimization through a grid-search of the target function χ given by:
N
N
M
i 1
i 1 j 1
i [( Pij,exp Pij,calc ) 2 / ij2 ]
where the sum was over M relaxation parameters for each of N residues, and Pij,calc were the
appropriate relaxation parameters calculated from the assumed model. The ij values were the
corresponding standard deviations of experimentally derived Pij,exp. The minimization
procedure delivered four global parameters (three diffusion coefficients and one Euler angle)
and N sets of local, residue specific parameters comprising Sf, Ss, f, s, and Rex. The residues
with incomplete experimental data (5 M < 9) were excluded from the procedure at this stage
of calculations. Their local parameters were determined separately for each residue by
minimizing partial target function i and using known global parameters obtained earlier. The
residues of both flexible termini were also excluded from the calculation delivering the
overall diffusion parameters. Those unstructured, flexible terminal segments cannot be
regarded as a part of the rigid rotor and description of their motions in terms of a single
overall correlation time does not seem to be appropriate (Alexandrescu and Shortle, 1994;
Brutscher et al., 1997). Model parameter uncertainties derived in the minimization of target
9
functions and i were obtained as standard deviations from 200 Monte Carlo simulations
(Press et al., 1986).
3. Results
3.1. Sequence-specific resonance assignment
The 2D 1H-15N HSQC spectrum of homodimeric S100A1 protein displayed relatively
good dispersion of amide correlation signals (Fig. 1). Cross peaks for all residues could be
assigned, except that of Asn 87, which was not observed due to line broadening most possibly
caused by an exchange process. The sequence-specific backbone and side-chain assignments
were completed using 2D and 3D NMR experiments. The 13Cβ chemical shifts for the unique
cysteine residue (Cys 85) showed that its thiol group was in the reduced state (Martin et al.,
2010; Sharma and Rajarathnam, 2000). All amide
resonances of
13
15
N resonances except that of Asn 87, all
C nuclei with attached protons and all 1HC and amide 1HN resonances have
been assigned unequivocally. 122 pairs of prochiral groups have been stereoassigned in each
monomer unit.
3.2. Three dimensional structure of human apo-S100A1
The three-dimensional structure of the protein was calculated based on NMR data
(1460 intrasubunit distance constraints and 63 dihedral restraints for each subunit together
with 132 intersubunit NOE-based distance constraints). 98.8 % of all residues are located in
the most favored (86.9%) or additionally allowed regions (11.9%) of the Ramachandran plot
giving an estimated precision of the structure comparable to the resolution of 1.7 Å of X-raybased structure. The statistics for the ensemble of 20 most favorable structures is given in
Table 1. A ribbon diagram for the lowest energy structure is shown in Fig. 2. The atomic
coordinates for all 20 structures of human apo-S100A1 have been deposited in the Protein
Data Bank with the accession code 2L0P.
10
Each subunit contains four α-helices formed by residues Glu 3–His 18, Lys 30–
Leu 41, Val 51–Asn 64 and Phe 71–Asn 86, two short 310-helical segments: Ser 42–Ala 47
and Asn 87–Phe 89 and one short antiparallel β-sheet formed by Leu 28, Ser 29, Glu 68, and
Val 69. The existence of helices, especially of the second short 310-helix at the C-terminus
was additionally confirmed by short and medium range NOEs from the 15N-edited NOESYHSQC spectrum.
For residues Glu 5–Cys 85 within one subunit, the RMSD for the backbone atoms (N,
C, C') equals 0.46 ± 0.07 Å and for all heavy atoms 0.97 ± 0.09 Å, while for both subunits in
the dimer, the corresponding values are 0.53 ± 0.08 Å and 1.01 ± 0.10 Å, respectively. Almost
identical RMSD values obtained for the single subunit and the homodimer prove that the
interface between subunits is unequivocally determined and the resulting ensemble of
structures represents a well defined homodimeric protein. The dimer interface is located
mainly between helices I and IV' and helices IV and I'. On average 4 – 5 long-range distance
restraints are observed for residues involved in the dimer formation.
3.3. Protein dynamics
The lowest energy S100A1 conformer was used to calculate the molecular inertia
moment tensor and the geometric coefficients Ak. The ratios of the inertia tensor principal
values were equal to Ix : Iy : Iz = 1.13 : 1.00 : 1.54 with the z axis collinear with the molecular
symmetry axis. Full relaxation data were available for 60 out of 92 backbone amides and they
were used for the simultaneous determination of 4 global parameters and 60 sets of local
parameters. Incomplete relaxation data for 4 additional residues (Leu 4, Ser 19, Leu 28,
Gln 48) were sufficient to obtain the local parameters. The remaining 28 residues, mostly
located at both termini and in helices I and IV, displayed partially superposed 1H/15N
correlations in the 2D spectra (see Figure 1). Therefore, their precise quantitative analysis was
not possible.
11
The principal values of the overall diffusion tensor are equal to:
Dx = (1.94±0.02)·107 s-1, Dy = (2.02±0.02)·107 s-1, and Dz = (1.83±0.01)·107 s-1. The angle
between the directions of Ix and Dx axes was found to be equal to 26 ± 7 deg. The averaged
isotropic rotational correlation time, R = (2Dx + 2Dy + 2Dz)-1 = 8.63±0.08 ns is typical for
globular proteins of about 20 kDa size and corresponds well to the results obtained for other
proteins from the S100 family (Bertini et al., 2009; Dutta et al., 2008; Inmam et al., 2001;
Zhukov et al., 2008).
Out of the five residue specific parameters describing the local mobility of backbone
amide N–H vectors within the frame of the extended model-free approach, S2=Sf2Ss2 and Rex
are of special importance. The former describes restrictions imposed on motions much faster
than the overall protein diffusion (the ps time scale) and the latter allows the detection of
much slower motions, but fast enough to average chemical shifts of the exchanging sites
(usually on the s – ms time scale). The S2 values of the residues located in both calcium
binding loops and in the linker indicate that the mobility of these protein segments is higher
than those situated in the helices. On the other hand, residues involved in chemical exchange
processes are predominantly located in the N-terminal calcium-binding loop (Fig. 3).
The obtained model parameters reproduce well input experimental data. The plot of
calculated vs experimental R2/R1 ratios is given in Supplementary Fig. S.1.
Since the local motion analysis was not possible for all backbone NH amide groups,
the S2 values were also predicted from the chemical shifts using TALOS+ software (Shen et
al., 2009). As can be seen in the upper part of Fig. 3 the predicted S2 values reasonably well
reproduce the experimentally determined ones.
Differences in mobility of the protein structural elements become much more evident
when the weighted means of S2 and Rex values determined for the residues building them are
compared. The data presented in Fig. 4 indicate that the outer helices I and IV are more rigid
12
than the inner ones: II and III. It seems to be a common feature of apo-S100 protein structures
as shown by relaxation studies of S100B (Inmam et al., 2001), S100A4 (Dutta et al., 2008),
and S100A5 (Bertini et al., 2009) proteins. It is also visible that small but significant Rex
values appear in the C binding loop. Summing up, comparison of the dynamics of the
structural elements of the protein reveals that, relative to helices, the binding loops display
increased mobility on both, ps and s – ms time scales, whereas the linker is more mobile
only within the range the ps time scale.
It is worth to mention here that the Rex values determined by us for 15N nuclei are by
no means correlated with their chemical shift changes induced in S100A1 proteins by calcium
binding as calculated from the data published for apo (Rustandi et al., 2002; BMRB accession
no. 4285) and holo (Wright et al., 2005; BMRB accession no. 6583) states of rat S100A1 and
fully confirmed by our preliminary studies of human holo protein (to be published). The most
striking examples corroborating this statement are as follows. The largest shift of 15N signal
induced by calcium binding is observed for Asp 63: as much as 10.8 ppm. Yet, its Rex value is
very low: 0.16 s-1. On the other hand, the highest Rex (3.7 s-1) has been found for Glu 22 amide
nitrogen (see Fig. 3) which signal position does not change considerably upon Ca2+
coordination ( = -0.9 ppm).
4. Discussion
The complete resonance assignment of aromatic residues allowed us to determine the
precise positions and orientations of aromatic side chain groups, especially that of Trp 90,
which displayed two unequivocal NOE contacts to Tyr 26' located in the N loop of
neighboring subunit: H1(W90)/H1(Y26') and HN(W90)/H1(Y26'). The three 1H resonances
involved in these contacts are well separated from others. Together with another unequivocal
NOE contact, H(N87)/H1(Y26'), they are decisive for the determination of the structure
13
terminating helix IV in the both protein subunits as a short 310-helix comprising Asn 87,
Phe 88 and Phe 89 residues. Such a tightening of the final -helix turn is not unusual (Barlow
and Thornton, 1988). This structure is less precisely defined than the -helical ones. The
lower number of NOEs per residue for the Asn 87–Phe 89 region (10.8 NOE/res. in
comparison with 34.3 NOE/res. for Phe 71–Asn 86 helix IV) is accompanied by the lack of
Asn 87 NH cross peak in the 2D 1H-15N HSQC spectrum suggesting the involvement of this
region in an exchange process.
Up till now there are three available structures of apo-S100A1 proteins: rat (1K2H)
(Rustandi et al., 2002), human (2L0P) [this paper], and bovine modified at Cys 85 by mercaptoethanol (2JPT) (Zhukov et al., 2008). All of them have been determined by NMR
methods. The positions of secondary structure elements as well as mutual orientations of
helices (Table 2), are very similar in all of the structures. Nevertheless, there are a few small,
but important, structural differences between these proteins.
Differences between the interhelical angles listed in Table 2 considerably exceed the
estimated error limits. They can be explained in two ways: either the method commonly used
to determine the -helix orientation and/or error estimation in such a structure analysis is
untrustworthy, or even very small changes in the protein sequence or its slight chemical
modification can result in noticeable structural changes. Both explanations should be taken
into account. Nevertheless, a dramatic increase of Ca+2-binding affinity of bovine S100A1
protein induced by its Cys 85 chemical modification strongly suggests that at least the latter is
correct.
From our point of view (see below) the most important structural fact is the absence,
in the structure of rat apo-S100A1 protein (Rustandi et al., 2002) of the helical turn formed by
residues 87-89 found in both human and modified bovine proteins as can be seen in Fig. 5.
All these structures are based on solid evidence provided by NOE spectra.
14
H1(W90)/H1(Y26') and HN(W90)/H1(Y26') cross peaks identified in the human protein
spectrum, decisive for determination of the 87-89 helix, were not observed in rat S100A1. On
the other hand, H(W90)/H(D46) and H(W90)/H(K49) signals, incompatible with the 8789 helical structure, are present only in the rat protein spectrum but not in that of human apoS100A1.
Nevertheless, the conformational differences between the C-termini of both proteins
can be much less dramatic than it looks at first glance. First of all, medium range NOE
signals: H(N87)/HN(W90) and HN(F88)/HN(W90), characteristic for a helical arrangement or
a turn of the involved residues have been observed by us in human apo-S100A1 but also by
Rustandi et al., (2002) in the rat protein. In fact, a hydrogen bonded turn comprising residues
N87 – W90 may be identified in this structure (Fig. 5a). Besides, the predicted S2 values (Fig.
3) clearly show that 87-89 helical structure in human apo-S100A1 is quite labile, so that this
protein segment can easily unwind and take some other, non-helical conformation. Therefore,
it seems that we deal here with an equilibrium between the helical and non-helical states
which are differently populated in both proteins.
This point is discussed here at length because of its importance for our studies whose
main goal is to find out why chemical modification of the Cys 85 SH group affects, in a
dramatic way, the calcium binding ability of S100A1 protein (Goch et al., 2005). We tried to
solve this problem by comparing the structure of bovine apo-S100A1 modified at Cys 85 by
-mercaptoethanol (Zhukov et al., 2008) with the only corresponding structure of S100A1
published at the time – that of the rat protein (Rustandi et al., 2002). This comparison led us
to the conclusion that the primary effect of Cys 85 modification by thiol molecules is the
elongation of helix IV (Figure 5c). The results presented in this paper do not confirm this
conclusion: in unmodified human S100A1 protein amino acid residues 87–89 participate in
the -helical conformation. Even if Cys 85 thionylation stabilizes this structure, such an effect
15
cannot explain the dramatic increase of the protein affinity for calcium ions. Further,
meticulous studies (see Introduction) are needed to solve this problem.
Nevertheless, comparison of modified bovine and native human apo-S100A1 can be
very instructive. Their structures were determined using very similar, though not identical,
experimental procedures and data analysis methods. Their sequences are much more similar
to each other than those of rat and human proteins: only two mutations: (A51→V and
D64→N) convert the sequence of bovine protein studied by us previously into that of human
S100A1 presented in this paper, whereas as many as six mutations are needed in the case of
rat protein (D35→E, S43→G, V47→A, A51→V, I57→V, and F74→Y). Moreover, the
dynamics of modified bovine and native human apo-S100A1 proteins have been measured
and analyzed using the same theoretical model.
Within error, the secondary structure of both these proteins is almost identical. This
statement, based on structural studies, is strongly supported by comparison of S2 parameters
(see Fig. 4) characterizing fast movements within a protein, which, in turn, are mainly
dependent on its secondary structure. Nevertheless, the tertiary/quaternary structure is
different. The Rex values for helices II and IV, the linker and the C-terminal loop are
considerably greater in the modified bovine protein that indicates that its global structure is
much more mobile than that of human apo-S100A1 protein. The results of dynamics
measurements are strongly supported by chemical shift differences of backbone nuclei signals
of both proteins calculated according to Ayed et al. (Ayed et al., 2001) and presented in
Figure 6. These observations lead to the conclusion that Cys 85 modification in S100A1
protein does not provoke its structural transition to any different, well defined, rigid structure,
but rather it has a general destabilizing effect.
Acknowledgments
16
This work was supported by a grant from the Ministry of Science and Higher
Education (N301 031234) and the EU Social Fund project (no. POKL.04.01.01-00-072/09 (to
Ł.J.). Authors thank dr Patric Groves for the help during preparation of manuscript.
Appendix A. Supplementary data
Supplementary data associated with this article can be found in the online version.
17
References
Alexandrescu, A.T., Shortle, D., 1994. Backbone dynamics of a highly disordered 131 residue
fragment of staphylococcal nuclease. J. Mol. Biol. 242, 527-546.
Ayed, A., Mulder, F.A.A., Yi, G.S., Lu, Y., Kay. L.E., Arrowsmith, C.H., 2001. Latent and
active p53 are identical in conformation. Nature Struct.Biol. 8, 756-760.
Baber, J.L., Szabo, A., Tjandra, N., 2001. Analysis of slow interdomain motion of
macromolecules using NMR relaxation data. J. Am. Chem. Soc. 123, 3953-3959.
Baldisseri, D.M., Rustandi, R.R., Zhang, Z., Tang, C., Bair, C., Landar, L., Zimmer, D.B.,
Weber, D.J., 1999. 1H, 13C, and 15N NMR sequence specific resonance assignments for
rat apo-S100A1(). J. Biomol. NMR 14, 91-92.
Barlow, D.J., Thornton, J.M., 1988. Helix geometry in proteins, J. Mol. Biol. 201, 601-619.
Bax, A., Clore, G.M., Gronenborn, A.M., 1990. 1H-1H correlation via isotropic mixing of 13C
magnetization: a new three-dimensional approach for assigning 1H and 13C spectra of
13
C-enriched proteins. J. Magn. Reson. 88, 425-431.
Berjanskii, M.V., Neal, S., Wishart, D.S., 2006. PREDITOR: a web server for predicting
protein torsion angle restraints. Nucl. Acids Res. 34, W63-W69.
Bertini, I., Gupta, S.D., Hu, X., Karavelas, T., Luchinat, C., Parigi, G., Yuan, J., 2009.
Solution structure and dynamics of S100A5 in the apo and Ca2+-bound states. J. Biol.
Inorg. Chem. 14, 1097-1107.
Bolewska, K., Kozłowska, H., Goch, G., Mikołajek, B., Bierzyński, A., 1997. Molecular
cloning and expression in Escherichia coli of a gene coding for bovine S100A1 protein
and its Glu32 Gln and Glu73 Gln mutants. Acta Biochim. Polon. 44, 275-284.
Brutscher, B., Brüschweiler, R., Ernst, R.R., 1997. Backbone dynamics and structural
characterization of the partially folded A state of ubiquitin by 1H, 13C, and 15N nuclear
magnetic resonance spectroscopy. Biochemistry 36, 13043-13053.
18
Clore, G.M., Szabo, A., Bax, A., Kay, L.E., Driscoll, P.C., Gronenborn, A.M., 1990.
Deviations from the simple two-parameter model-free approach to the interpretation of
nitrogen-15 nuclear magnetic relaxation of proteins. J. Am. Chem. Soc. 112, 4989-4991.
Dalle-Donne, I., Rossi, R., Colombo, G., Giustarini, D., Milzani, A., 2008. Protein Sglutathionylation: a regulatory device from bacteria to humans. Trends Biochem. Sci. 34,
85-96.
Delaglio, F., Grzesiek, S., Vuister, G.W., Zhu, G., Pfeifer, J., Bax, A., 1995. A
multidimensional spectral processing system based on UNIX pipes. J. Biomol. NMR 6,
277-293.
Dixon, M., Webb, E.C., 1961. Enzyme fractionation by salting-out: a theoretical note. Advan.
Protein Chem. 16, 197-219.
Dutta, K., Cox, C.J., Basavappa, R., Pascal, S.M., 2008. 15N relaxation of apo-Mts1: a
dynamic S100 protein. Biochemistry 47, 7637-7647.
Falconer, J.S., Jenden, D.J., Taylor, D.B., 1953. The application of solubility measurements to
the study of complex protein solutions and to the isolation of individual proteins.
Discuss. Faraday Soc. 13, 40-50.
Farrow, N.A. Muhamdiram, R., Singer, A. U., Pascal, S.M., Kay, C.M., Gish, G., Shoelton,
S.E., Pawson, T., Forman-Kay, J.D., Kay, L.E., 1994. Backbone dynamics of a free and a
phosphopeptide-complexed Src homology 2 domain studied by 15N NMR relaxation.
Biochemistry 33, 5984-6003.
Fushman, D., 2003. Determination of protein dynamics using 15N relaxation measurements,
in: Zerbe, O. (Ed.), BioNMR in Drug Research. Wiley-VCH, Weinheim, Germany, pp.
283-308.
Goch, G., Vdovenko, S., Kozłowska, H., Bierzyński, A., 2005. Affinity of S100A1 protein for
calcium increases dramatically upon glutathionylation. FEBS J. 272, 2557-2565.
19
Goddard, T.D., Kneller, D.G., SPARKY3, University of California, San Francisco.
Grzesiek, S., Bax, A., 1992. Correlating backbone amide and side chain resonances in larger
proteins by multiple relayed triple resonance NMR. J. Am. Chem. Soc. 114, 6291-6293.
Grzesiek, S., Bax, A., 1993. Amino acid type determination in the sequential assignment
procedure of uniformly 13C/15N-enriched proteins. J. Biomol. NMR 3, 185-204.
Grzesiek, S., Anglister, J., and Bax, A., 1993. Correlation of backbone amide and aliphatic
side-chain resonances in 13C/15N-enriched proteins by isotropic mixing of 13C
magnetization. J Magn. Reson. B101, 114-119.
Guentert, P., Qian, Y.Q., Otting, G., Muller, M., Gehring, W., Wuethrich, K., 1991. Structure
determination of the Antp (C39S) homeodomain from nuclear magnetic resonance data
in solution using a novel strategy for the structure calculation with the programs DIANA,
CALIBA, HABAS and GLOMSA. J. Mol. Biol. 217, 531-540.
Guentert, P., Mumenthaler, C., Wuethrich, K., 1997. Torsion angle dynamics for NMR
structure calculation with the new program Dyana. J. Mol. Biol. 273, 283-298.
Heizmann, C.W., Fritz, G., Schafer, B.W., 2004. S100 proteins: structure, functions and
pathology. Frontiers in Bioscience 7, d1356-d1368.
Herrmann, T., Guentert, P., Wuethrich, K., 2002. Protein NMR structure determination with
automated NOE-identification in the NOESY spectra using the new software ATNOS. J.
Biomol. NMR 24, 171-189.
Ikura, M., Kay, L.E., Bax, A., 1990. A novel approach for sequential assignments of 1H, 13C
and 15N spectra of larger proteins: heteronuclear triple-resonance three-dimensional
NMR spectroscopy. Application to calmodulin. Biochemistry 29, 2659-4667.
Ikura, M., and Bax, A., 1991. An efficient 3D NMR technique for correlating the proton and
15
N backbone amide resonances with the -carbon of the preceding residue in uniformly
15
N/13C enriched proteins. J. Biomol. NMR 1, 99-104.
20
Inmam, K.G., Baldisseri, D.M., Miller, K.E, Weber, D.J., 2001. Backbone dynamics of the
calcium-signaling protein apo-S100B as determined by 15N NMR relaxation.
Biochemistry 40, 3439-3448.
Kay, L.E., Keifer, P., Saarinen, T., 1992a. Pure absorption gradient enhanced heteronuclear
single quantum correlation spectroscopy with improved sensitivity. J. Am. Chem. Soc.
114, 10663-10665.
Kay, L.E., Nicholson, L.K., Delaglio, F., Bax, A., Torchia, D.A., 1992b. Pulse schemes for
removal of the effects of cross correlation between dipolar and chemical-shift anisotropy
relaxation mechanisms on the measurement of hteronuclear T1 and T2 values in proteins.
J. Magn. Reson. 97, 359-375.
Keller, R.L.J., 2004. The computer aided resonance assignment tutorial, Cantina Verlag,
Goldau, Switzerland. Downloaded from: www.cara.nmr.ch
Koradi, R., Billeter, M., Wüthrich, K., 1996. MOLMOL: a program for display and analysis
of macromolecular structures. J. Mol. Graph. 14, 51-55.
Laskowski, R.A., Rullmann, J.A.C., MacArthur, M.W., Kaptein, R., Thornton, J.M., 1996.
AQUA and PROCHECK-NMR: programs for checking the quality of protein structures
solved by NMR. J. Biomol. NMR 8, 477-486.
Lipari, G., Szabo, A., 1982. Model-free approach to the interpretation of nuclear magnetic
resonance relaxation in macromolecules. J. Am. Chem. Soc. 104, 4546-4570.
Lohr, F., Ruterjans, H., 1995. A new triple-resonance experiment for the sequential
assignment of backbone resonances in proteins. J. Biomol. NMR 6, 189-197.
Marenholtz, I., Heizmann, C.W., Fritz G., 2004. S100 proteins in mouse and man: from
evolution to function and pathology (including an update of the nomenclature). Biochem.
Biophys. Res. Commun. 322, 1111-1122.
21
Marion, D., Ikura, M., Tschudin, R., Bax, A., 1989. Rapid recording of 2D NMR spectra
without phase cycling. Application to the study of hydrogen exchange in proteins. J.
Magn. Reson. 85, 393-399.
Martin, O.A., Villegas, M.E., Vila, J.A., Scheraga, H.A., 2010. Analysis of
13
C and
13
C
chemical shifts of cysteine and cystine residues in proteins: a quantum chemical
approach. J Biomol NMR 46, 217-25.
Morin, S., Gagne, S.M., 2009. Simple tests for the validation of multiple field spin relaxation
data. J. Biomol. NMR 45, 361–372.
Muhandiram, D. R., Farrow, N. A., Xu, G.-Y., Smallcombe, S. H., Kay, L. E., 1993. A
gradient 13C NOESY-HSQC experiment for recording NOESY spectra of 13C-labeled
proteins dissolved in H2O. J. Magn. Reson. B102, 317-321.
Muhandiram, D.R., Kay, L.E., 1994. Gradient-enhanced triple-resonance three-dimensional
NMR experiments with improved sensitivity. J. Magn. Reson. B103, 203-216.
Ottinger, M., Bax, A., 1998. Determination of relative N-HN, N-C', C-C', and C-H
effective bond lengths in a protein by NMR in a dilute liquid crystalline phase. J. Am.
Chem. Soc. 120, 12334-12341.
Press, W.H., Flannery, B.P., Teukolsky, S.A., Vetterling, W.T., 1986. Numerical recipes. The
art of scientific computing. Cambridge University Press, Cambridge, chapter 14.
Rustandi, R.R., Baldisseri, D.M., Inmam, K.G., Nizner, P., Hamilton, S.M., Landar, A.,
Landar, A., Zimmer, D.M., Weber, D.J., 2002. Three-dimensional solution structure of
the calcium-signalling protein apo-S100A1 as determined by NMR. Biochemistry 41,
788-796.
Santamaria-Kisiel, L., Rintala-Dempsey, A.C., Shaw, G.S., 2006. Calcium-dependent and independent interactions of the S100 protein family. Biochem. J. 396, 201-214.
22
Schwieters, C.D, Kuszewski, J.J., Tjandra, N., Clore, G.M., 2003. The XPLOR-NIH NMR
molecular structure determination package. J. Magn. Reson. 160, 66-74.
Sharma, D., Rajarathnam, K., 2000.
13
C NMR chemical shifts can predict disulfide bond
formation. J Biomol NMR 18, 165-171.
Shen, Y., Delaglio, F., Cornilescu, G., Bax, A., 2009. TALOS+: a hybrid method for
predicting protein backbone torsion angles from NMR chemical shifts. J. Biomol. NMR
44, 213-223.
Tjandra, N., Feller, S.E., Pastor, R.W., Bax, A., 1995. Rotational diffusion anisotropy of
human ubiquitin from 15N NMR relaxation. J. Am. Chem. Soc. 117, 12562-12566.
Vriend, G., 1990. WHAT IF: a molecular modeling and drug design program. J. Mol. Graph.
8, 52–56.
Wishart, D.S., Bigam, C.G., Yao, C.G., Abildgaad, F., Dyson, H.J., Oldfield, E., Markley,
J.L., Sykes, B.D., 1995. 1H, 13C and 15N chemical shift referencing in biomolecular
NMR. J. Biomol. NMR 6, 145-140.
Wittekind, M., Mueller, L., 1993. HNCACB, a high-sensivity 3D NMR experiment to
correlate amide proton and nitrogen resonances with the alpha- and beta-carbon
resonances in proteins. J. Magn. Reson. B101, 201-205.
Woessner, D.E., 1962. Nuclear spin relaxation in ellipsoid undergoing rotational Brownian
motion. J. Chem. Phys. 37, 647-654.
Wright, N.T., Varney, K.M., Ellis, K.C., Markowitz, J., Gitti, R.K., Zimmer, D.M., Weber,
D.J., 2005. The three-dimensional solution structure of Ca(2+)-bound S100A1 as
determined by NMR spectroscopy. J. Mol. Biol. 353, 410-426.
Yamazaki, T., Forman-Kay, J. D., Kay, L. E., 1993. Two-dimensional NMR experiments for
correlating 13C and 1H/ chemical shifts of aromatic residues in 13C-labeled proteins
via scalar couplings. J. Am. Chem. Soc. 115, 11054-11055.
23
Zhang, O., Kay, L.E., Olivier, J.P., and Forman-Kay J.D., 1994. Backbone 1H and 15N
resonance assignments of the N-terminal SH3 domain of drk in folded and unfolded
states using enhanced-sensitivity pulsed field gradient NMR techniques. J. Biomol. NMR
4, 845-858.
Zhukov, I., Bayer, P., Schoellermann, B., Ejchart, A., 2007. 15N magnetic relaxation of
backbone dynamics of the ribosome-associated cold shock response protein Yfia of
Escherichia coli. Acta Biochim. Polon. 54, 769-775.
Zhukov, I., Ejchart, A., Bierzyński, A., 2008. Structural and motional changes induced in
apo-S100A1 protein by the disulfide formation between its Cys 85 residue and mercaptoethanol. Biochemistry 47, 640-650.
24
Figure captions
Figure 1. 1H–15N HSQC spectrum of human apo-S100A1 protein. The terminal NH2 groups
of Asn and Gln residues are not labeled.
Figure 2. Ribbon diagram of the lowest energy structure of human apo-S100A1 protein.
(Color reproduction on the Web and in print)
Figure 3. Experimental data (vertical bars) and predicted values from TALOS+ (Shen et al.,
2009) (full circles) for order parameter values S2 (upper part) and exchange terms Rex at 9.4 T
(lower part) with corresponding error bars for amino acid residues of human apo-S100A1
protein.
Figure 4. Weighted means of order parameter values <S2> (upper part) and exchange terms
<Rex> at 9.4 T (lower part) with corresponding error bars for amino acid residues in various
structural elements of human apo-S100A1 and bovine apo-S100A1-ME proteins. The
experimental 15N relaxation data for bovine apo-S100A1-ME protein (Zhukov et al., 2008)
were reprocessed as described in the Analysis of 15N relaxation data section, i.e. assuming
fully anisotropic molecular reorientation and extended model-free approach.
Figure 5. Ribbon representations of helices IV and C-terminal segments (residues 70 – 93) in
three structures of apo-S100A1 proteins: (a) rat, (b) human, (c) bovine modified with mercaptoethanol at Cys 85. A hydrogen bonded turn may be recognized for residues N87 –
W90 in the rat protein.
(Color reproduction on the Web and in print)
25
Figure 6. The differences of the chemical shift combination, tot, for backbone nuclei
between S100A1 protein (BMRB accession code: 16360) and its thionylated derivative
(BMRB accession code: 4982). The formula proposed by Ayed et al. (Ayed et al., 2001) is
given as: tot = [(HN)2 + (0.154·N)2 + (0.341·CO)2 + (0.276·C)2]1/2. The horizontal
line corresponds to the mean value of tot.
Supplementary Figure S.1. The plot of calculated vs experimental R2/R1 ratios. Correlation
coefficient for the displayed data r2 > 0.99.
26