supplemental material
advertisement

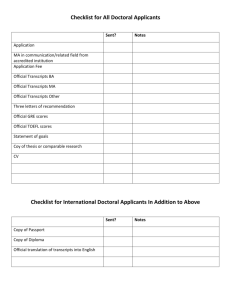
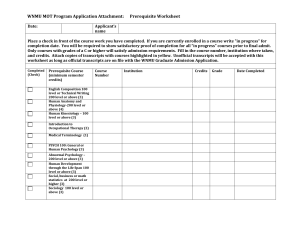
SUPPLEMENTAL DATA Supplemental Text 1 Molecular methods RNA extraction: Around 200 male Drosophila mayaguana flies (stock 15081-1397.03 from the Tucson Stock Center) were dissected for their accessory glands. Dissections were done in PBS with RNA later (Ambion). Accessory glands were pooled together and kept in RNA later at –20°C until RNA extraction. Total RNA was extracted with the RNAeasy extraction kit (QIAGEN) and mRNA was isolated from total RNA using the Oligotex mRNA kit (QIAGEN). cDNA synthesis and library construction: cDNA synthesis was done with the Super SMART cDNA Synthesis Kit (Clontech). This kit uses PCR to increase the amount of initial mRNA. Female cDNA generated by the same process was used as probe for subtraction of housekeeping genes and other non-male specific genes using the PCRSelect cDNA Subtraction Kit (Clontech). A potential drawback of using the Super SMART cDNA synthesis kit is that the protocol involves a restriction reaction, which may lead to the restriction of some of the cDNA, generating a lot of short fragments. To decrease the chance to clone these short fragments, the last PCR product of the subtraction process was size fractionated for fragments larger than ~700bp. Size fractionation was carried out in 2% low-melting agarose gel stained with SYBR. PCR fragments were retrieved from the gel using the Wizard gel purification kit (PROMEGA). After a quick incubation with Taq polymerase for adding overhanging A’s, the retrieved fragments were cloned into a TA plasmid vector (Topo TA cloning kit, Invitrogen). Chemically competent cells (Invitrogen) were then transformed to produce an accessory gland subtracted library. Dot blot: To do a further screening of accessory gland specific genes, a dot-blot procedure was carried out. About 600 white colonies were picked and used for PCR amplification of inserts using standard M13 primers. PCR products were gathered in 96well plates and cleaned using a BIOMEK robot. 5ml of the cleaned PCR product was mixed with 5ml of 0.6N NaOH and 1ml of this mixture was blotted onto positively charged nylon membranes. DNA was immobilized on the membranes by UV- crosslinking. Membranes were hybridized with DIG labeled female cDNA as probe. Hybridization and visualization was done with the DIG starter kit (Roche), according to the manufacturer’s protocol. Sequencing and editing: Inserts were sequenced with the M13 forward primer and a PolyT primer. Sequences were obtained using an ABI3730xl automated sequencer and edited with Sequencher v4.2 (Gene Codes). Vector and adaptor sequences were trimmed and similar sequences were assembled in contigs. Supplemental Text 2 Acp classification Dot-blot data was available for 75 of the unique sequences. Among those, 48 had a clear, negative result (no hybridization with female probes; Table 1). Presence of a signal peptide can be evaluated using three different statistics (S-mean, D-score, and p), as implemented in the program SignalP. Since these statistics can be sometimes discordant, here we chose to report all three of them for easy of comparison with other studies (Table 1). Considering any of the three statistics, 64 transcripts had more than 50% of chance of containing a signal peptide. Among the sequences for which a signal peptide was not detected, many seem to be lacking their 3’ end and therefore the presence of signal peptide in the gene cannot be ruled out (Supplemental data - Table 1, n*). Among the 91 unique transcripts uncovered in the D. mayaguana library, 18 matched at least three of the Acp criteria (Table 1, category 1). Also classified as Acps were 12 transcripts with negative dot-bolt results and a high probability of having a signal peptide as supported by the three statistics obtained with SignalP, for which there were no Acp orthologs in other species or known Acp related conserved domain (Table 1, category 2). For some transcripts (15), the dot blot results were inconclusive, but homology to an Acp or a common Acp functional domain, together with strong evidence of a signal peptide, was considered good indicators of an Acp status (Table 1, category 3). Finally, nine other transcripts were classified as Acps even though there was little support for the presence of a signal peptide, if (a) there was evidence of another marker of secretory sequence as detected by the program SecretomeP and/or (b) there was no evidence that the 3’ tip of the mRNA sequence was complete (Table 1, category 4). These sequences also had either criteria 1 (homology) or 4 (function) and had a negative dot blot. These classification steps resulted in a total of 54 likely Acp candidates. Among the transcripts that were not classified as Acps, 18 could possibly be Acps (Supplemental data – Table 1, category 5), since there is no strong evidence against their inclusion as Acps (dot blot results were negative, they did not have domains that point to other non-Acp function, and the presence of a signal peptide could not be ruled out). The remaining 19 transcripts either had a clear, positive hybridization in the dot blot tests or a non-Acp associated function. Supplemental Text 3 Gene families with more than three identified genes in D. mayaguana Cluster 7: Members of Cluster 7 had D. mojavensis Acp16b and Acp24 as their best hits in the D. mojavenis reproductive tract library database. WAGSTAFF and BEGUN (2005b) identified two paralogs of Acp16b in D. mojavensis, named Acp16a and Acp16c (although they did not mention the similarity between Acp16 and Acp24). To have a better understanding of the orthology of genes in the two species, we carried a phylogenetic analysis including the two D. mayaguana transcripts and D. mojavensis Acp16a, Acp16b, Acp16c, and Acp24. Although likelihood and maximum parsimony trees did not agree on all the gene relationships, the two analyses were in accordance in that D. mayaguana transcripts form a clade, while Acp16a, Acp16b, and Acp16c (of D. mojavensis) cluster together in another clade (Supplemental data - Figure 2). The D. mayaguana transcripts had as their best hit in the D. mojavensis genome the same locus as Acp24 when using BLASTn, but were more similar to the Acp16b locus if the program used was tBLASTx. All these loci, including the D. mayaguana similar transcripts, had several hits similar to the beginning of their sequences (30 – 80bp), in a genomic region spanning ~25kb. This is a transposon rich genomic region, but none of the transcripts matched a transposon repeat sequence itself. Also, none of these genes have a conserved protein domain. Cluster 11: The genes included in cluster 11 hit the same D. melanogaster Acp in the BLAST searches, Acp53C14c. Acp53C14c is the most divergent of four paralogous genes (plus Acp53Ea, Acp53C14a, Acp53C14b) present in D. melanogaster (HOLLOWAY and BEGUN 2004). The genomic region is conserved across Drosophila species and may include different numbers of in tandem repeats of these genes. For instance, D. pseudoobscura have seven similar ORFs in this microsyntenic region, but there is no information on whether all these copies are expressed and where (WAGSTAFF and BEGUN 2005a). The four D. mayaguana sequences in cluster 11 matched three D. mojavensis Acps: Acp1, Acp2, and Acp25. These three loci are in adjacent regions of the D. mojavensis genome. In fact, the region encompassing these loci, spans approximately 5.5 kb and has seven ORFs identified by different methods, including Oxford pipeline, NCBI Gnomon, GeneWise (BIRNEY et al. 2004), and Contrast (GROSS et al. 2005). A phylogenetic analysis including all the D. mayaguana paralogs and orthologous sequences in D. mojavensis, D. arizonae, and D. mulleri (Supplemental data – Figure 3b), showed that relationships, within each ortholog clade, are compatible with the phylogenetic hypothesis most accepted for the mulleri cluster (DURANDO et al. 2000). The trees suggest that mayAcp2a is the product of an early duplication with subsequent loss in D. mojavensis or in the lineage leading to this species. Cluster 12: With a total of nine distinct sequences, Cluster 12 is composed of transcripts that had Acp5a or Acp23 as best hits in the D. mojavensis Acp database. This cluster is composed mostly of short transcripts (163 - 356bp), although one of them is 925bp long. Most of these transcripts were found in two or more clones and seven of these sequences had a signal peptide as detected by the program SignalP 3.0, indicating that they are likely to represent secretory genes. These transcripts did not have significant hits in other Drosophila species. An alignment of the 10 D. mayaguana sequences and the three homologs of D. mojavensis showed that although there are many large and short indels and a lot of substitutions, there is one long (100bp) plus a few short blocks that are conserved. No conserved protein domain was detected in the CD-search, however. Repetitive elements were searched in these genes using RepeatMasker 3.0 (SMIT and GREEN 1996-2004), but no significant association was found. Nevertheless, they match transposon sequences located in a transposon rich region in the genome of D. mojavensis. A phylogenetic analysis of the sequences suggest species specific duplications. A third copy of Acp5 was also found in the D. mojavensis genome (WAGSTAFF and BEGUN 2007). Cluster 13: The largest cluster (Cluster 13 in Table 4) included 10 transcripts. These sequences were not similar to any D. mojavensis Acps or other ESTs found in the reproductive tract of males of this species. They have similarity, (1e-11 > E > 1e-33) to five adjacent loci in the D. mojavensis genome, although only three of these loci were best hits for each of the D. mayaguana transcripts in this cluster. All D. mayaguana transcripts in this cluster were relatively short (220 – 450bp), with a peptide signal and a conserved Kazal-type serine protease inhibitor domain (cd00104). These genes do not seem to have orthologs in any Drosophila species (Supplemental data – Table 2). The dot blot results were variable, with some being apparently specific to accessory glands, while others showed strong hybridization to female probes. When searched against each other, every sequence hit all the other family members, but E-values varied considerably (1e-11 > E > 1e-169), showing that there are distinct degrees of similarity among them. A phylogenetic analysis of the family showed that its members fall into three main clades and all clades have members that met the Acp criteria (Supplemental data – Figure 5). D. simulans - 31 D. sechellia - 30 D. melanogaster - 30 D. yakuba - 33 D. erecta - 31 D. ananassae - 31 D. pseudoobscura - 36 36 D. persimilis - 36 D. willistoni - 36 D. mojavensis - 87 D. virilis - 39 D. grimshawi - 34 Supplemental Figure 1. Number of D. mayaguana accessory gland transcripts with significant BLAST hits (E < 1e-05) in each Drosophila species with a complete genome sequence. Phylogeny taken from FlyBase. Supplemental Figure 2. Maximum likelihood tree illustrating relationships among members of Cluster 7. Prefix of gene names refer to the species they belong to: D. mojavensis (moj), D. arizonae (ari), and D. mayaguana (may). Supplemental Figure 3a. Maximum likelihood tree depicting relationships among members of Cluster 11 identified in D. mojavensis and D. mayaguana through cDNA libraries (sequences with prefixes mojAcp and mayAcp, respectively) and D. mojavensis predicted ORF’s in the correspondent genomic region. Gene predictions were obtained by Oxford pipeline (TRdmoj prefix), GeneWise (GI_EISE; BIRNEY et al. 2004), Gnomon (GI_NCBI; http://www.ncbi.nlm.nih.gov/genome/guide/gnomon.html), and Contrast (GI_BATZ; GROSS et al. 2005). Supplemental Figure 3b. Maximum likelihood tree depicting relationships among members of Cluster 11. Notice the long branches between paralogs as opposed to shorter ortholog branches’ lengths. Prefix of gene names refer to the species they belong to: D. mojavensis (moj), D. arizonae (ari), and D. mayaguana (may). Supplemental Figure 4. Maximum likelihood tree depicting relationships among members of Cluster 12. Supplemental Figure 5. Maximum likelihood tree depicting relationships among members of Cluster 13.