meiosis and crossing over
advertisement

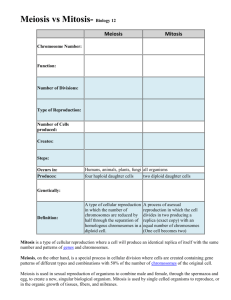
STUDY GUIDE: CHAPTER 8 The Cellular Basis of Reproduction and Inheritance Intro.: An attempt was made to fertilize with pollen the last known wild kuhihewa plant, deep in the Hawaiian rain forest, using pollen from kuhihewa plants grown in a botanical garden. The effort failed, but it introduces the effort to maintain wild species that are on the verge of extinction. Also, it illustrates fertilization of the egg with sperm (from pollen grains). Sea stars have sexual reproduction and can also regenerate lost arms, or in some cases, divide into two organisms, through asexual reproduction. Life cycle: Sequence of life stages leading from adults of one generation to adults of next generation. Two basic phases: 1) Development from fertilized egg to adult, 2) Reproduction, the formation of new fertilized eggs from adults. Sexual reproduction: The reproductive process involving union of egg and sperm. Asexual reproduction: The production of offspring by a single parent without participation of egg and sperm. Each egg and sperm carries a set of the genetic info for that species—the organism’s genome. Offspring of sexual reproduction thus inherit traits from two parents. 8.1 The saying “Like begets like” in its strictest sense applies only to the asexual reproduction: offspring in this case is identical genetically to the one parent. Some organisms reproduce primarily by asexual reproduction. An example: the Amoeba, a protist. Chromosomes are structures that contain most of the organism’s DNA. In sexual repro., offspring resemble the two parents and each other, but show variations from them, as well. 8.2 Cells arise only from preexisting cells “All cells come from cells”: Rudolf Virchow’s principle from 1858. Sexual and asexual reproduction depend on cellular reproduction, commonly referred to as cell division. 8.3 Prokaryotes reproduce by binary fission Binary fission: a type of cell div. used by prokaryotes and diagramed in Fig. 8.3A. The chromosome must be replicated in the process of binary fission. For E. coli, the chromosome is 500 times the length of the cell, so this shows the complexity of duplicating it within the cell. The two daughter chromosomes of the cell going thru binary fission, once formed, begin to separate, probably thru growth of the plasma membrane, to which they appear to be fixed. Binary fission is not complete until the two resulting cells are entire, able to operate independently. 8.4 The large, complex chromosomes of eukaryotes duplicate with each cell division. Chromosomes are only visible as entire structures when the cell is dividing. At other times exist as a mass of thin fibers of DNA and protein, called chromatin. Eukaryotic chromosomes contain many more protein molecules, which help to organize the DNA and control the activity of its genes. No. of chromosomes depends upon the species: humans have 46. Well before the cell actually begins to divide, the chromosomes are duplicated. The result, visible as cell division progresses, are called sister chromatids: the chromosome doubled but remaining as a single structure until separated in the process of cell division. The two sister chromatids are joined together at the centromere, which may be located at various points on different chromosomes. 8.5 The cell cycle multiplies cells Some cells in the human body divide once a day, some less often, others not at all: ex.: mature nerve and muscle cells. Cell division must be controlled to prevent chaos: It goes thru an orderly process called the cell cycle: an orderly sequence of events that extends from the previous cell division to the next one. Most of the time of the cell cycle is spent in interphase: a period when cell growth takes place, chromosomes are duplicated; many other cell parts are also made. Three stages in Interphase: G1 (Gap 1), S (DNA synthesis), and G2. Actual sorting of chromosomes to two nuclei and division of the cell into two daughter cells occurs in the Mitotic phase, which includes mitosis and cytokinesis. Mitosis: The nucleus and the chromosomes are evenly divided to the two daughter nuclei. Cytokinesis: The cytoplasm is divided into two. 8.6 Cell division is a continuum of dynamic changes Five major steps in cell division: 1) Interphase: chromosomes and many other cell structures are replicated, cell growth occurs. The chromosomes remain in the nucleus as a mass of chromatin. 2) Prophase, divided into two steps: Early prophase, in which the chromatin fibers become densely compacted, discrete chromosomes become visible under light microscope, each chromosome visible as two sister chromatids joined at the centromere. Mitotic spindle begins to form. Late prophase: the nuclear envelope breaks up, chromosomes are attached to the mitotic spindle at the centromeres. Kinetochore: the point of the centromere region attached to the mitotic spindle (one on each sister chromatid). Metaphase: the chromosomes, still as paired sister chromatids, are lined up in a plane approximately equidistant from the two poles of the cell: the metaphase plate, all attached to the mitotic spindle. Anaphase: the sister chromatids have separated, begin to move apart towards the poles, as cell elongates. Telophase and cytokinesis: the chromosomes have moved to opposite ends, nucleus reforms, cell divides into 2 daughter cells. Chromosomes uncoil into chromatin fibers, nucleoli reappear. 8.7 Cytokinesis differs for plant and animal cells By a process called cleavage, the cleavage furrow marks the division of animal cells: cell appears to “pinched” into two by a constriction. In plant cells, the formation of a cell plate takes place, which allows for maintaining the integrity of the cell walls. The cell place grows outward from the center, as vesicles containing membrane and material for the cell walls fuse with the growing cell plate. 8.8 Anchorage, cell density, and chemical growth factors affect cell division. Controlling the timing of cell division is important for an organism throughout its life. Ex.: cells lining the stomach must be replaced daily, but other cells duplicate less often. Anchorage dependence: cells from a cell culture placed in a petri dish do not start dividing until allowed to anchor on the surface of the dish. Cells, once they start dividing, usually stop when they touch one another: densitydependent inhibition. Clearing a space in a cell culture is similar to cutting yourself: when you cut yourself, cells immediately begin dividing, stopping when the space is filled (cells contact each other), the wound “healed”. A growth factor is a protein secreted by certain cells that stimulates cells nearby to start dividing. Most cells require growth factor to start dividing, and they stop when they run out of it. 8.9 Growth factors signal the cell cycle control system The cell cycle control system is a cyclically operating set of proteins in the cell that both triggers and coordinates major events in the cell cycle. This system has three key checkpoints: the G1, G2, and M (mitotic) positions (Fig. 8.9A). At these checkpoints, progress to the next stage is stopped until everything necessary for the subsequent stage is ready. For many cells the G1 checkpoint is the most critical: if they don’t get the signal at this point, they become non-dividing cells, which many of the body’s cells are. We discussed signal transduction in section 5.13. As shown in Fig. 8.9B, signal transduction may be involved in the satisfaction of the G1 checkpoint. 8.10 Growing out of control, cancer cells produce malignant tumors. Cancer is a disease of the cell cycle. Cancer cells don’t have a properly operating cell cycle control system. Excessive growth results in a mass of cells called a tumor. A benign tumor is a mass of essentially normal cells: they do cause serious problems, but normally can be removed safely simply by surgery. In contrast, a malignant tumor is cancerous, a benign is not. A malignant tumor is a mass of cancerous cells capable of spreading to other parts of the body. Metastasis: The spread of cancer cells beyond their original site. Four categories of cancers, based on the tissues originating from: Carcinomas: of the external or internal covering of the body (skin, etc.) Sarcomas: supportive tissue cancers—bone, muscle for ex. Leukemias and lymphomas: cancers of the blood-forming tissues—bone marrow, spleen, & lymph nodes. Cancer cells are not usually affected by density-dependent inhibition—will pile up on one another, some breaking off from the mass and possibly entering the lymph system. Treatments for cancer are directed at stopping cells from dividing. Normal cells don’t divide as rapidly (some not at all) as do cancer cells: so they are not necessarily affected. However, cells of the testes and ovaries may be affected, with the result of sterilization. Other bad side effects can also occur. Chemotherapy and radiation therapy generally work to inhibit cell division. 8.11 Review of the functions of mitosis: growth, cell replacement, and asexual reproduction New cells produced through mitosis have exactly the same no. & types of chromosomes as the parents. Mitosis makes possible growth, replacement of tissues, and budding of asexually produced offspring (as in the Hydra) having the same genes as the parent. MEIOSIS AND CROSSING OVER 8.12Chromosomes are matched in homologous pairs All body cells except gametes (eggs and sperm) are referred to as somatic cells, and in humans have 46 chromosomes. Each chromosome has a twin with centromere in same position, same length, same staining pattern (with partial exception of the sex chromosomes, a single pair). These are the homologous chromosomes, and the 23 pairs can be identified as unique pairs in metaphase of mitosis. The individuals of the pair carry genes for the same characteristics, and these are located in essentially the same locations on both members of the pair. The one pair of chromosomes which may not be identical in appearance are the sex chromosomes: the chromosomes which determine sex, the X and Y chromosomes. Women have two X chromosomes, men an X and a Y. Although the X and Y chromosomes carry a few homologous genes, they are dissimilar in also carrying the genes necessary for sexual reproduction for their respective sex. However, they do act as a homologous pair in meiosis. The remaining 22 pairs are the autosomes. For all homologous pairs, we inherit one individual from the mother, the other from the father. How this comes about, the inheritance of each homologous pr. from the different parents, begins with the process called meiosis. 8.13Gametes have a single set of chromosomes All species which reproduce sexually, including humans, inherit one chromosome of each pair from each parent, as said above. Diploid cells: those which contain two homologous sets of chromosomes, the diploid number, or 2n in humans is 46. The only non-diploid cells are the gametes (eggs and sperm): each of these has one complete set, 23 chromosomes, and said to be haploid (1n). Sexual intercourse allows the haploid sperm and egg to unite, producing the diploid zygote, or fertilized egg cell. This process is called fertilization. Through meiosis, a special type of cell division that only produces sex cells (gametes), these haploid cells is brought about, and of course meiosis occurs only in the reproductive organs. Remember that mitosis produces only 2n cells: the daughter cells have the same no. of chromosomes as the cell they originate from. 8.14Meiosis reduces the chromosome number from diploid to haploid. Meiosis comes about through two cell divisions, starting with the replicated chromosomes (sister chromatids). Divided into two stages: Meiosis I, in which the sister chromatids remain together but homologous pairs separate, and Meiosis II, in which the sister chromatids separate. At the end of Meiosis I the two daughter cells are said to be haploid, since they no longer have homologous pairs. The result of meiosis is four daughter cells, each of which contains the haploid number of chromosomes. 1) Interphase: like in mitosis, chromosomes are replicated, producing sister chromatids. 2) Prophase I: The homologous pairs form tetrads: the pairs actually intertwine, and sometimes identical parts of homologous chromosomes are exchanged in a process called crossing over, explained more completely later in this chapter. 3) Metaphase I: the tetrads are gathered on the metaphase plate, using the spindle fibers connected to the kinetochores of each tetrad. 4) Anaphase I: The homologous pairs separate, being moved to opposite poles by the spindle fibers. 5) Telophase I and Cytokinesis: the haploid cells, still with sister chromatids attached, divide, producing two haploid daughter cells. After cytokinesis, in some organisms the nuclear envelope re-forms and chromosomes uncoil, followed by a short interphase. In other organisms, Telophase I and cytokinesis are immediately followed by Prophase II. 6) Meiosis II is basically similar to mitosis, but with only half the number of chromosomes. It starts with haploid cells. 8.15. Review: A comparison of mitosis and meiosis Mitosis involves one cell division, producing two diploid cells. Meiosis entails two nuclear and cytoplasmic divisions, resulting in 4 haploid cells. All events unique to meiosis occur in Meiosis I: the duplicated homologous chromosomes pairing to produce tetrads, these tetrads moving to the Metaphase I plate, and being separated with the sister chromatids remaining together. 8.16. Independent orientation of chromosomes in meiosis and random fertilization lead to varied offspring. As discussed earlier, offspring of sexual reproduction are different genetically from one another, as well as from their parents. This arises thru several mechanisms, one being independent movement of maternal and paternal chromosomes of each homologous pair: each offspring winds up with a complete set of chromosomes from each parent, but these are randomly collected from the maternal and paternal chromosomes of each parent. See diagram in Fig. 8.16. This results in a multitude of possibilities for the genetic makeup of each child. Each egg and sperm are likely to be different, and random event through which sperm will fertilize an egg adds to the possibilities. For humans, the 23 pr. of chromosomes gives a possible 223 combinations for each gamete, which is about 8 million possible combinations. The possible combinations of chromosomes for the zygote of a union of one man and one woman becomes 8 million X 8 million or 64 trillion possibilities! In spite of the above randomness of chromosome inheritance, we will find that there are additional possibilities for added variation in the genetics of offspring. 8.17 Homologous chromosomes carry different versions of genes. The importance of the sorting of various chromosomes to each gamete becomes clearer in that each chromosome of a homologous pr. may carry different versions of a particular gene. These versions are alleles, which we will talk about more in Ch. 9. The ex. of eye color and coat color of mice is used here: each is determined by one gene, each of which is known to have two or more alleles. Eye color can be black (B allele) or pink (b allele). Hair color can be brown (C allele) or white (c allele). The obvious result is that chromosomes with different alleles can yield two different types of chromosomes. 8.18 Crossing over further increases genetic variability. Crossing over: the exchange of corresponding segments of two homologous chromosomes. This occurs in Prophase I of meiosis. The location of the crossing over is called a chiasma., and the crossing over occurs between homologous but non-sister chromosomes. It occurs in synapsis—see Fig. 8.14—at which time chromosomes are tightly in contact in a precise gene-by-gene alignment. The crossing over adds to the genetic variability in the offspring. With crossing over, a particular chromosome you receive from your mother may have genes from both your maternal grandfather and maternal grandmother. Fig. 18 a and b. We now have discussed 3 sources of genetic variability in sexually reproducing organisms: 1) Crossing over 2) Independent orientation of chromosomes at metaphase I 3) Random fertilization Mutations are ultimately responsible for genetic variability, since this is how the different alleles came about. Importance of the variation will become more apparent once we study evolution. 8.19 A karyotype is a photographic inventory of an individual’s chromosomes. Errors in meiosis can result in abnormal numbers of chromosomes or in chromosomes with abnormal structures. These can be detected through karotyping the person’s chromosomes. To do a karotype, the cells must be caught at the metaphase of mitosis, when they are most condensed with sister chromatids present. Frequently this is done by taking a blood sample, and stimulating the white blood cells called lymphocytes to divide. Once they have been multiplying for a short period, the cells are treated with a chemical which freezes the lymphocytes at metaphase. Next step is concentrating the white blood cells, then staining and mounting them on a slide. Photographs are taken of cells with the most visible metaphase spreads. Then, individual chromosomes can be cut from the photographs, identified to which pair each belongs, and a complete photographic set of a person’s chromosomes is assembled on paper. 8.20 An extra copy of chromosome 21 causes Down syndrome. Down syndrome, the karotype shown in Fig. 8.20A, is caused by trisomy 21,the presence of an extra chromosome 21. This is the most common chromosome abnormality. It is likely that embryos with other autosomal chromosome abnormalities do not survive to birth. 8.21Accidents during meiosis can alter chromosome number. Nondisjunction occurs when members of a chromosome pair fail to separate. The result can be a chromosomal abnormality such as Downs syndrome. End at Sec. 8.21.