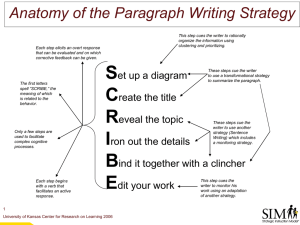
Dot-probe paradigm
advertisement

Introduction to Chapter 3
The experiments in Chapter 2 were designed to investigate whether the presence
of an upright face might capture and hence bias the early allocation of visual attention to
its spatial proximity. In order to determine this, we tested participants on a task that relied
heavily on visual spatial attention, in the peripheral presence/absence of an upright face.
A significant performance decrement on the primary task, in the presence of an upright
face would have supported the claim that faces captured visual-spatial attention (cf. de
Gelder & Rouw, 2001).
However, the consistent finding across all the four experiments showed that the
orientation of a face-distractor had no impact on how well participants performed on the
visual search task. In this chapter, a different visual attention paradigm is used to confirm
whether an upright face’s inability to capture attention is isolated only to that used in
visual search processes, or if it is a general rule. Furthermore, the experiment reported
here addresses certain limitations of the general experimental design that the experiments
in Chapter 2 were based upon.
In the concluding section of Chapter 2, it was suggested that upright faces might
be able to exert exogenous control in the absence of any endogenous control of attention.
The attentional literature contains a dichotomy of exogenous and endogenous attention
(e.g. {Corbetta, 2002 #85}. The former is said to be stimulus-driven and the latter, goaldirected. Having an goal-directed course of action such as a well-defined search-region or
target object-category, could have had an over-riding effect over any exogenous control
that upright faces might have exerted on attention (e.g. {Yantis, 1990 #115}.This could
explain why upright faces failed to capture spatial attention and hence, disrupt the visual
search process in the experiments reported in Chapter 2.
If upright faces capture visual-spatial attention, it might only do so if doing so is
not explicitly antagonistic to an internal goal-directed form of attention (such as with
experiments 1, 2, 3, 4). Supposing this to be true, a task-paradigm is required that will not
encourage the formation of any endogenous control of attention. Firstly, upright faces
will have to appear within the search-space of the primary task, instead of outside it. In an
experiment that also used a letter-detection task, face-learning were found for upright
faces that served as a task-irrelevant background {Jenkins, 2002 #219}. Secondly,
participants should not be primed towards the identification of any particular object
category class, at the detriment of all others. As mentioned in Chapter 2, the active search
strategy for alphabetic letters could have inadvertently suppressed the exogenous
influence of upright faces on attention. The current hypothesis is that upright faces can
act as effective exogenous cues, that orient spatial attention, but only in the absence of
endogenous cues e.g. task-strategy. To test this, an experiment was carried out based on
the dot-probe paradigm, a design that is commonly employed to demonstrate how early
allocation of visual attention can be controlled by exogenous cues.
Dot-probe paradigm
The dot-probe method is usually used to assess preattentive and attentional biases
to information embodied in visual stimuli, particularly in relation to threat and anxiety
(e.g. {Broadbent, 1988 #229;Bradley, 1998 #227;Mogg, 1999 #225;Pishyar, 2004 #223}.
A dot-probe trials simply requires participants to detect a probe that is initially hidden
from view by two juxtaposed images. This format was adapted from paradigms used in
experimental cognitive psychology that showed how spatial attention can be assessed
from the speed of manual responses to visual probes (e.g. {Posner, 1980 #228}. Although
the position of the dot-probe is not usually predictable by the nature of the image that
hides it, certain images can result in faster probe-detection when they do share the same
spatial location as the probe. For example, probes hidden behind threatening images were
quicker to be detected by highly anxious individuals than those preceded by a neutral
image under certain conditions {Mansell, 1999 #224;Bradley, 1999 #226;Bradley, 1998
#227}. Some researchers argue from such evidence that certain images can induce an
attentional bias in the observer, that is based on its informational content {Pishyar, 2004
#223;Mogg, 1999 #225}.
Existing dot-probe experiments with face-stimuli have mostly been interested in
the emotional valence of face-stimuli, and how levels of anxiety in observers correlate
with their exhibited levels of attentional bias exhibited towards threatening faces {Pishyar,
2004 #223;Mansell, 1999 #224;Mogg, 1999 #225}. Interestingly, this is an effect that is
not consistently replicable with threatening non-face stimuli e.g words {Mansell, 2002
#230;Pishyar, 2004 #223}. Pishyar and colleagues (2004) recently concluded that the use
of emotional face-stimuli in dot-probe tasks is a more sensitive index of attentional bias
than emotional words.
With dot-probe tasks, attentional-bias towards or away from a particular imagetype is always measured in relation to its accompanying control image. For example,
there exists an attentional bias to threatening faces, but only in relation to a neutrally
expressive face (e.g. {Pishyar, 2004 #223}. The question that will be addressed here is
whether there exists an early attentional bias even to upright face-stimuli, when compared
to inverted faces that differ only in terms of orientation. In the study reported here the
element of emotional valence was removed from the face-stimuli and only faces with
neutral expressions were used. Thus, according to de Gelder and Rouw (2001), upright
faces which are said to capture spatial attention should
Facilitatory and Inhibitory components of orienting
The dot-probe task is essentially based on an experimental approach that has early
precedence in the understanding of attentional capture; namely, the cuing-paradigm. Such
studies explored the facilitatory and inhibitory influences that a brief non-informative cue
could exert in the detection of a target that follows it (e.g. {Eriksen, 1973 #231;Posner,
1980 #228}. Thus, a brief non-informative visual cue presented in the periphery can
speed up the detection response to a target that appears in the same location within
100msecs of the cue, compared to a target that might appear in a different location
{Posner, 1980 #228}. The elegance of the cuing paradigm lied in its ability to prove that
spatial shifts of attention can occur, even in the absence of observable motor behaviour
e.g. eye-movements {Posner, 1980 #228;Posner, 1990 #110}.
Effective orienting could be induced by either by peripheral and central cues. A
peripheral cue oriented attention by drawing it towards itself, whereas a central cue
appeared at fixation and directed attention symbolically e.g. arrow cues {Jonides, 1981
#232;Posner, 1980 #228}. Peripheral cues were said to exert exogenous shifts in attention
whereas central cues appealed to the observers’ endogenous knowledge of its connotated
meanings. The dot-probe paradigm which forms the basis of the study reported here,
investigates exogenous rather than endogenous shifts of attention, since there is a
simultaneous presentation of two images on both sides of fixation that act as competing
peripheral cues. In such an experimental design, the greater efficacy of either image to act
as a cue would indicate there there is something concerning the differences in visual
characteristics across the two images, that allows one image to orient exogenous attention
more effectively than the other. Such a paradigm is thus ideal for the purposes of the
current study as it is the exogenous shifts in spatial attention that upright faces are
claimed to induce {de Gelder, 2001 #33}. Furthermore, it allows for a direct comparison
to be made between the upright face-image and a control image i.e. inverted-face, for the
ability to capture attention. Unlike central cues, peripheral cues are said to require no
interpretation or cognitive effort in order to capture attention. As a result, attentional
shifts induced by peripheral cues are said to be reflexive and impossible to suppress
compared to central cues, even if they are manipulated to be consistently incongruous to
the probe-position {Jonides, 1981 #232}. Again, this mirrors the claim made by de
Gelder and Rouw (2001), in that face-detection is a process that is fast and automatic.
Usually, facilitation for the detection of a target is associated with the occurence
of a peripheral cue that precedes it in the same location {Jonides, 1981 #232;Posner,
1980 #228}. As mentioned already, this is explained by imputing a spatial shift of
attention to the appearance of a peripheral cue that goes on to make a positive
contribution to the detection of items in the corresponding spatial region. However, there
is a caveat to this rule. It has been found that facilitation for target detection by a valid
cue is highly dependent on short cue-target onset asynchronies; that is, the time-interval
between presentations of the cue and target. Specifically, facilitation only occurs with
short cue-target onset asynchronies (~100msec). With longer cue-target onset
asynchronies (~200msecs), a valid cue results in the inhibition of response for a targetdetection instead {Posner, 1984 #233;Klein, 2000 #234}. This late component of
orienting has been termed ‘inhibition of return’, which suggests that early orienting of
attention to a spatial location that does not yield a target is subsequently replaced with
inhibition that serves to prevent attention from re-orienting back to a searched location
{Posner, 1985 #235}.
Understanding these two components of orienting – namely, the early component
of faciliation and the late component of inhibition – allows for a further prediction to be
made from using upright faces as cues in the dot-probe paradigm. If upright faces truly
exhibit exogenous control over attention, performance in a dot-probe task ought to be
facilitated or inhibited depending on the time-interval between the upright face cue and
probe. Hence, if the spatial position of the upright face cue correspond to the probe’s,
performance should not only be speeded up at short intervals, but it should also be slowed
down for longer onset asynchronies between cue and probe.
Experiment 5: Dot probe design
In the final experiment, we investigate if upright faces are able to act as valid
exogenous cues in visual-spatial attention. This experiment was based on the dot-probe
task paradigm described in previous sections. On each trial, a probe appeared either on
the left or right of fixation and participants were faced with a two alternative forcedchoice task that required them to make an appropriate keypress that indicated the probe’s
position. Presentation of the probe was always preceded by 2 images that were presented
on the both sides of the fixation cross that the probe could appear in. The images used
were either upright or inverted face stimuli. In addition, each pair of images was either a
mixed pairing of an upright and inverted face each or consisted of both upright or
inverted face stimuli.
Given that two cues were always provided on both sides of the fixation cross in
each trial, there was no incentive for the participant to show an orienting bias to either
cue. This was especially so, when the trials were balanced to ensure that the probe was
just as likely to follow an upright face as it was, an inverted one.
Despite this, the claim that upright faces captures attention predicted a spatial
shift of attention towards the side of fixation that corresponded with the upright face cue,
when presented in conjunction with a neutral cue i.e. inverted face. This was expected to
result in faster probe-detection for when the upright face was a valid cue compared to
when the probe followed the inverted face cue instead. However, this prediction is only
valid for short cue-probe intervals less than 100 msec. Lengthening the cue-probe
intervals to greater than 200 msec was expected to induce inhibition in the spatial regions
that benefit from early facilitation. Thus, detection for probes cued by upright faces was
expected to be slower than those cued by inverted faces instead, for long cue-probe
intervals of about 500 msec.
However, the above predictions could be only made for when the upright face cue
is presented alongside an inverted face counterpart. As a control condition, same-cue
pairings were introduced and probe detection that followed from these pairings were not
expected to exhibited any facilitatory effect at all. Even if upright faces can serve as valid
peripheral cues, Posner and colleagues (1980; 1984) have previously argued that selective
attention cannot be simultaneously divided across 2 objects. They arrived at this
conclusion when simultaneous cuing for both the peripheral positions that a probe could
have appeared in failed to speed its detection over non-cued probes, at short cue-probe
asynchronies {Posner, 1980 #228;Posner, 1984 #233}. Nonetheless, inhibition for this
double-cuing procedure was maintained at both cued positions for long onset
asynchronies of cue-probe despite this lack of an initial facilitatory effect. It will be
interesting to note whether upright faces if they do capture attention, can demonstrate the
same pattern of performance. In comparison to the results presented by Posner and
colleagues (1980; 1984), a double-cuing procedure with upright faces ought to show no
facilitatory effect for probe detection at short cue-probe onset asynchronies. However, it
could still bring about an inhibitory aftereffect at long cue-probe onset asynchronies. This
could be compared to trials of the same double-cuing design but with the use of inverted
faces that are not thought to capture attention, and thus expected to demonstrate neither
facilitatory nor inhibitory effect on the probes cued.
Method
Design
This experiment followed a 2 (valid-cue type: upright vs. inverted) x 2 (pairing
type: same vs. different) x 3 (cue-probe timings: see following for details) repeatedmeasures design. Altogether, there were 3 experimental blocks of 96 trials each. At the
start of each trial, a pair of images was presented, each on either side of the fixation cross.
One of the two images would share the same position as the probe and termed the valid-
cue. This was either be an upright or inverted face. The image accompanying the valid
cue was either an upright or inverted face; that is, same or different to the cue and thus
determined the independent variable of pairing-type. Finally, the experimental blocks
were each defined by the 3 varying time intervals of cue-probe and presentation length of
cue. Specifically, these could be described as: i) cue (presentation time: 60 msec) – 0
msec cue-probe interval; ii) cue (presentation time: 60 msec) – 440 msec cue-probe
interval; iii) cue (presentation time: 500 msec) – 0 msec cue-probe interval. Condition i)
represented short cue-probe asynchronies whereas conditions ii) and iii) were variations
of long cue-probe asynchronies. The trials within each block were equally distributed
across the 4 trial conditions and counterbalanced for cue position; that is, left/right of
fixation cross. In addition, the blocks are counter-balanced for serial order of presentation
across participants.
Participants
18 (age range: 18-19 years) undergraduate students of the Department of
Psychology, University of Manchester, participated in this experiment as part of their
course requirements. All participants reported normal or corrected-to-normal vision.
Materials and apparatus
The experimental set-up was similar to the experiments reported in Chapter 2.
The experiment was controlled by an experimental generator software (Psyscope v.1.1.2),
on a Macintosh G4. A viewing distance of 96cm was maintained throughout the
experiment by use of a fixed chinrest.
The same set of 24 photorealistic images of different faces, used in the
experiments reported in Chapter 2, rendered a total of 48 upright and inverted faceimages that were used as valid cues. For the condition of pairing-type, each valid cue was
randomly paired with an image of a different face, that was either of the same or different
orientation to itself. Hence, there were a total of 96 unique pairings, each of which was
used only once in each experimental block. Fig. 3.1. shows an example of the four
different type of pairings.
Figure 3.1: Examples of possible pairings between a valid cue and accompanying non-valid cue,
represented by the left and right image respectively. Valid cue-type: upright face (a, b), inverted face
(c,d); Pair-type: same (a,d), different (b,c).
Procedure
Each participant had to perform a 2 alternative forced-choice task in this
experiment. On each trial a fixation cross was first presented for 1000 msec, followed by
a pair of images (valid cue and accompanying image), each on either side of the fixation
point. Participants were informed, prior to experimentation, that the these images were
irrelevant to the task and held no information concerning probe location. Following this,
the pair of images was replaced by a single probe that appeared on the side of the fixation
cross, formerly occupied by the valid cue. Figure 3.1 illustrates the design of this
experiment as well as the respective timings depending on the experimental block it
occurred in. Participants were required to make fast and accurate keypress responses that
indicated where they believed the probe to have appeared, relative to fixation.
Nonetheless, the probe remained on the screen for as long as was necessary for
participants to respond.
The entire experiment was made up of 3 blocks of 96 experimental trials each and
participants were allowed to take a break in between the experimental blocks. Each block
of trials was defined by its unique combination of length of presentation for the cuepairing and cue-probe onset asynchrony (see Fig 3.2.).
keypress response
(b) i) 0 msec
ii) 440 msec
iii) 0 msec
(a) i) 60 msec
ii) 60 msec
iii) 500 msec
Fig. 3.2: Diagram illustrating the design of experiment 5. (i), (ii) and (iii) represent the collective timing
conditions for the presentation time cue (a); and timing interval between cue-offset and probe-onset (b),
that defined each of the experimental blocks. Inter-trial interval of 500 msec.
At a fixed distance, the valid cue and accompanying non-cue subtend a maximum
visual area of 1.5º x 2.1º each. Each cue is centred 3.1º on either side of the fixation cross
which is itself, 0.42º x 0.42º.
Results
The dependent variable taken was mean detection latency, measured from probeonset. Collected timings were subjected to a 3 (Timing conditions) x 2 (valid cue type) x
2 (non-valid cue type) repeated measures ANOVA. There was only 1 main effect of that
was statistically significant, timing conditions (F(2,34)=11.7, p<0.001). Neither the
variable of valid cue-type (F(1,17)=0.02, p=0.89) nor pairing-type (F(1,17)=0.02, p=0.90)
brought about significant diferences in detection latency. Table 1 presents a summary of
the mean reaction times across the 3 independent variables.
Upright-face
Inverted-face valid cue
Different
431
(18.9)
Same
436.0
(20.3)
Different
442
(19.7)
Same
431.0
(16.0)
60(cue present.)+440 (blank interval)
417
(15.3)
431.6
(22.9)
418
(17.2)
420.5
(15.9)
500(cue present.)
365
(17.2)
370.1
(19.1)
381
(23.1)
361.8
(16.1)
Timing conditions
60 (cue present.)
Table 1: Summary of mean RTs in milliseconds (standard error is bracketed) across timing conditions for
when i) the valid cue is an upright-face vs. inverted-face; and when the pairings were same or different.
Pairwise comparisons indicate that long presentation times, i.e. 500ms, of cues
resulted in significantly shorter mean RTs. When the presentation times of cues are short,
i.e. 60ms, the addition of a 440ms blank interval did not bring about any significant
difference in response times to probe. Finally, There were no significant interactions to
report from this study’s results. Details are reported in Appendix A.
Discussion
In chapter 2, upright faces were shown to be unable to capture attention, at the
expense of goal-directed behaviour. This study was carried out with the intention of
determining whether upright faces could, nonetheless, orient attention when it was not
contrary to goal-directed behaviour. Particularly, face-stimuli fell within the task
designated area in Experiment 5. Furthermore, the target was a single item and detection
was not reliant on object-identification at all. This is in contrast to the experiments
previously reported, whereby target detection performance was reliant upon the
participants’ ability to discriminate the target letter from an array of non-target items on
the basis of the target’s identity.
Despite these modifications, upright faces continued to have no effect in orienting
spatial shifts of attention towards their locality. Having an upright face as a valid cue type
did not influence probe responses in the same way as valid peripheral cues are generally
reported to. Earlier, it was predicted that at shorter cue-probe onset asynchronies (less
that 100msec), a valid upright face cue should facilitate probe-detection if it resembled a
valid peripheral cue (cf{Posner, 1984 #233;Posner, 1980 #228}. Thus, faster detection
ought to have been associated to probes cued by an upright faces, compared to inverted
faces. This was not the case in Experiment 5. Furthermore, longer cue-probe onset
asynchronies (~500 msec) ought to have resulted in an inhibition of attending to the
spatial region formerly occupied by valid upright face cues, such as to induce longer
detection latencies. Again, this prediction was not borne out.
Inhibition was also thought to occur for long cue-probe onset asynchronies, when
both cues were upright faces, without the facilitatory effect because early orienting of
attention is not expected to be dedicated to more than one peripheral cue (cf. {Posner,
1984 #233;Posner, 1980 #228}. Specifically, this would have been expressed by a
significant interaction across all the independent variables of timing conditions, valid
cue-type and pairing-type. The facilitative effect to probe-detection for trials with valid
upright-face cues would only be exhibited with when it was paired with an inverted face,
and at short cue-probe onset asynchronies. Furthermore, long cue-probe onset
asynchronies would have had an inhibitory effect on the detection of probes cued by
upright faces, regardless of the face-image it was paired with. However, this critical
interaction was not proven to be significant.
Thus, the significant effects predicted by the original hypothesis that upright faces
exert an exogenous control over spatial attention was not demonstrated in Experiment 5,
even when attempts were made to remove any form of endogenous control.
Instead, the significant main effect highlighted in this experiment was that of
timing conditions. Regardless of the type of face-image that cued the probe, a longer
interval between the cue and probe presentation resulted in quicker detection of the probe.
A possible explanation for this could that participants were simply better prepared to
respond when more time was given for the particpants to make a decision after the
presentation of the paired cues .
The findings reported here indicate that the image of an upright-face does not
function as a valid peripheral cue any more so than an inverted-face image. This runs
counter to the notion of an automatic face-detection system that automatically orients
spatial shifts of attention to the visual presence of faces {de Gelder, 2001 #33}. If this
claim was true, the results of Experiment 5 should have indicated early facilitation, and
late inhibition, effects on detection towards probes cued by upright faces. Instead, the
results did not reflect such a trend.
These results mirror those reported in Chapter 2 and suggests that the upright face
configuration might not orient attention any more than an inverted face configuration,
whether or not endogenous controls of attention are in place.