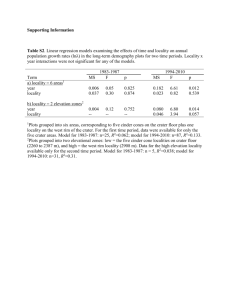
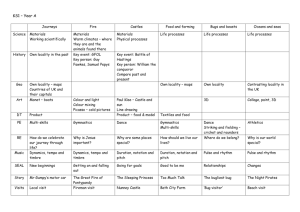
PALYNOLOGICAL CONTRIBUTIONS TO THE
advertisement

PALYNOLOGICAL CONTRIBUTIONS TO THE CHRONOZOGY AND STRATIGRAPHY OF THE HARTFORD BASIN IN CONNECTICUT AND MASSACHUSETTS BRUCE CORNET ALFRED TRAVERSE Department of Geosciences The Pennsylvania State University University Park, Pennsylvania 16802 ABSTRACT Recent discoveries of palynoflorules at numerous localities in the Newark Group basins of the eastern United States provide new evidence for correlation of deposits in these basins. Floras from the Shuttle Meadow and Portland Formations of the Newark Group rocks in the Hartford Basin of Connecticut and Massachusetts indicate that the Triassic-Jurassic boundary is located within the rocks of this basin: a shift of Corollina from more than 90% C. meyeriana to more than 90% C. torosus occurs somewhere between the Shuttle Meadow and Portland Formations and generally indicates a Rhaeto-Liassic age. In the Shuttle Meadow Formation, the overall shape of the palynoflora (particularly the presence of Convolutispora klukiforma), associated fish, paleomagnetic data, and radiometric dates, as well as megafossil evidence of Brachyphyllum scotii and Clathropteris meniscoides, support a basal Liassic age for this formation. Reptilean evidence in older strata of the Hartford Basin suggests that the Triassic-Jurassic boundary may lie just below the Shuttle Meadow Formation. In addition the Portland Formation contains a palynomorph association clearly favoring early to middle Liassic age. Comparisons are made on the basis of studies of recently discovered Newark Group palynoflorules of early Liassic age from Virginia and of Carnian-Norian age from New Jersey. Paleobotanical and geological evidence is summarized, showing that the predominantly Corollina (Hirmerella) palynofloras of the Hartford Basin were associated with a warm, seasonally wet and dry climate, and casting doubt on the idea that Corollina-producing plants composed a swamp association within the original basin. Twenty-seven genera and 42 species are described, as are 1 spore and 1 pollen type of uncertain taxonomic position. Eight new species are described: Camerosporites reductiverrucatus n. sp., Corollina murphyi n. sp., Cycadopites andrewsii n. sp., Cycadopites durhamensis n. sp., Cycadopites westfieldicus n. sp., Dictyophyllidites paramuensteri n. sp., Foveosporites agawamensis n. sp., Verrucosisporites cheneyi n. sp. Six new combinations are proposed: Araucariacites punctatus (Nilsson) comb. nov., Callialasporites segmentatus (Balme) comb. nov., Corollina itunensis (Pocock) comb. nov., Corollina simplex (DanzéCorsin & Laveine) comb. nov., Cycadopites reticulatus (Nilsson) comb. nov., Granulatisporites infirmus (Balme) comb. nov. The genera Circullina and Corollina are formally emended, as are the species Corollina torosus (Reissinger) Klaus and Circulina simplex Malyavkina. INTRODUCTION Recent discoveries of palynomorphs in most of the major Newark Group basins of the eastern United States (Text-fig. 1) provide new evidence for correlating Newark Group strata with European TriassicJurassic type sections. The Hartford Basin TEXT-FIGURE 1. "Newark" basins of eastern North America. Asterisks indicate basins in which palynoflorules have been found. palynofloras of Connecticut and Massachusetts (Text-fig. 2) provide at present the most comprehensive data for upper Newark Group correlation, and form an assemblage different from Triassic assemblages of other Newark Group basins. The Hartford Basin assemblage is largely Early Jurassic in age; it is not restricted to this basin, but is also present in the Newark Basin of New Jersey and the Culpeper Basin of Virginia. The recognition of Jurassic deposits in the Newark Group (Cornet et al., 1973), which until recently has been considered entirely Triassic, is not new. As early as 1855, E. Hitchcock, Jr., presented paleobotanical and geological evidence for a possible Rhaeto-Liassic age range for the Hartford Basin sediments. Several other geologists and paleontologists of the mid-1800s speculated that the deposits of the Hartford Basin extend into the Jurassic, but no convincing evidence was then available. Newberry (1888) described a small megafossil flora from the Connecticut Valley but considered it Rhaetian. Since then reptilean and fish remains have been interpreted as indicating only a Late Triassic age (Reeside et al., 1957). Recently Reeve and Helsley (1972), in attempting to explain the paleomagnetic discrepancy between the upper portion of the Chinle Formation, New Mexico, and the igneous rocks of the upper Newark Group, suggested a post-middle Rhaetian, but pre-Pliensbachian age for much of the upper portion of the Newark Group. The palynofloras from the Shuttle Meadow and Portland Formations of the Hartford Basin largely confirm this suggestion, indicating that the Triassic-Jurassic boundary exists within the strata of this basin. TEXT-FIGURE 2. Hartford and Deerfield basins of Connecticut and Massachusetts. Numbers 1-13 indicate palynoflorule localities discussed in this paper. Middletown, Conn., is at Locality 13. North Guilford is just south of Locality 1. HARTFORD BASIN STRATIGRAPHY The Hartford Basin sediments and igneous rocks are divided into four sedimentary and three volcanic formations or units. The lowest formation, the New Haven Arkose, has been computed to have a maximum stratigraphic thickness of 2600 m near the eastern edge of the basin, decreasing to around 1500 m near the western edge (Krynine, 1950) (Text-fig.3). The Talcott Formation conformably overlies the New Haven Arkose in southern Connecticut, but TEXT-FIGURE 3. Stratigraphic units of the Hartford Basin, showing their relative position and the stratigraphic location of palynoflorule localities 1-13 (cf. Text-fig. 2). This is a restored composite section from multiple sources, representing generalized North (left)-South (right) trends. The double line to the left of Gaillard Graben indicates a fault, but because of restoration, no displacement is shown along the fault. Locality 2 has the same position as Locality 1 and has therefore been omitted. apparently it is missing in northern Connecticut and Massachusetts. In most of southern Connecticut, the Talcott is a single basalt flow of variable thickness (30-75 m). Sanders (1970) has shown that within the Gaillard Graben, which is restricted to the southeastern part of the basin, the Talcott Formation exists as four lava-flow units interbedded with three sedimentary units, totaling about 305 m. The Gaillard Graben has been interpreted as a post-depositional structure (Sanders, et al., 1963, p. 11). However, the existence outside the Graben of only one (the lowest?) lava flow suggests that penecontemporaneous erosion removed much of the once continuous Talcott Formation before deposition of the overlying Shuttle Meadow Formation, and the Talcott was preserved in the graben as a result of downward displacement of sediments in the graben before the erosional cycle began. On the other hand, the absence in northern Connecticut and Massachusetts of basalt units referable to the Talcott might indicate only that the basalts did not extend that far north. However, it seems reasonable that 300 m of superimposed volcanic cones in Mt. Hitchcock in northern Massachusetts (Bain, 1941) may be related to the lava flows of the Gaillard Graben, judging from stratigraphic position and thickness. The Shuttle Meadow Formation, like the Talcott Formation, varies considerably in thickness and in extent over the basin. In the southernmost part of the basin, within the Gaillard Graben, the Shuttle Meadow Formation reaches its maximum thickness of about 270 m (Sanders, 1970). The formation steadily decreases in thickness northward: 170 m in North Guilford, Conn.; 94 m near Middletown, Conn.; 94 m at type section, New Britain, Conn.; 30 m in Farmington, Conn.; 15 m near Simsbury, Conn. (Krynine, 1950; Lehmann, 1959; Schnabel, 1960). The upper part of the formation appears to be the most extensive. North of Simsbury the Talcott basalt disappears, and the Shuttle Meadow Formation, if it continues north, would be recognized as the uppermost part of the New Haven Arkose in northern Connecticut and Massachusetts. The Holyoke lava-flow unit conformably overlies the Shuttle Meadow Formation in Connecticut, and the New Haven Arkose and Hitchcock volcanics in Massachusetts. This unit, consisting of at least two lava flows, ranges in thickness from 120 to 170 m over most of the basin, but it has been recorded under 92 m near the eastern and northern edges of the basin (Bain, 1941). The East Berlin Formation conformably overlies the Holyoke lava-flow unit, and appears to thicken both northward and southward from its minimum thickness near Avon, Conn.: 326-458 m in the Gaillard Graben, Branford, Conn.; 170-180 m near Middletown, Conn.; 140-150 m near Avon, Conn.; 250 m near Westfield, Mass. (Krynine, 1950; Lehmann, 1959; Schnabel, 1960; Colton and Hartshorn, 1966; Sanders, 1970). In the northern part of the basin, the upper part of the East Berlin Formation interfingers with the Granby Tuff and local lava flows (Brophy et al., 1967). These volcanics and pyroclastics contain at least one flow that apparently is continuous with the Hampden lava-flow unit (Bain, 1941). The Hampden lava-flow unit conformably overlies the East Berlin Formation and ranges in thickness from 18 to 60 m. Much of this variation in thickness seems to be the result of differences in the number of internal units or flows, which varies from two to eight (Chapman, 1965). In southeastern Connecticut a 12 m sedimentary unit has been found separating two flows (Sanders, 1970). The Portland Formation, which conformably overlies the Hampden lava-flow unit and interfingers with some of the Granby Tuff (Bain, 1941), has a maximum thickness of more than 2000 m in Massachusetts. The "Chicopee shale" and "Longmeadow sandstone" are here considered as local members, at present recognized only in Massachusetts. In Connecticut, where reported thickness rarely exceeds 920 m, post-depositional faulting and erosion are responsible for removing much of this formation. DESCRIPTIONS OF PALYNOFLORULE LOCALITIES Of the 13 palyniferous localities thus far discovered in the Hartford Basin (Text-fig. 2, 3), 4 have been selected as palynoflorule localities for purposes of formal description in this paper. Samples from the 9 remaining localities have yielded the same species of palynomorphs that are found at the formally described localities. Palynoflorule Locality No. 8 Formation: Upper New Haven Arkose. Stratigraphic position: 116 ± 6 m below Holyoke lava-flow unit. Geographic location: Massachusetts; Hampden County, 2.5 km south of Mt. Tom, just west of Holyoke; roadcut on Southampton Road, 2.65 km east of junction with Country Road. Palyniferous horizon: Tan to dark gray siltstone, .3-m-thick fossil plant bed situated within a 4.7+-m-thick sequence of orange to tan coarse sand stone and conglomerate. Remarks: Plant bed has produced dozens of well-preserved Clathropteris meniscoides Brongniart fronds and Equisetites stems. Palynoflorule Locality No. 1 Formation: Shuttle Meadow Formation. Stratigraphic position: 100 ± 6 m below Holyoke lavaflow unit. Geographic location: Connecticut; New Haven County, North Guilford; stream outcrop on east side of Totoket Mountain, .825 km north of Bluff Head. Palyniferous horizons: Thin layers of gray micaceous silty shale just below and .5 m above l-m-thick lake bed containing abundant fossil fishes; additional layer 4.5 m above lake bed, consisting of variegated greenish gray to red, poorly bedded silty claystone interbedded within a red siltstone and silty shale sequence. Remarks: See Cornet et al. (1973), for more detailed description of section. Palynoflorule Locality No. 5 Formation: Lower Portland Formation. Stratigraphic position: 257± 6 m above Hampden lava-flow unit. Geographic location: Connecticut; Middlesex County, just west of Cromwell; neat top of ravine, overlooking Chestnut Brook and Connecticut Route 9. Palyniferous horizon: 8-cm-thick lens of greenish gray, silty shale, which is part of a micaceous, arkosic sandstone sequence with numerous dinosaur foot-prints. Remarks: Palyniferous horizon apparently thickens to 38 cm about 100 m upstream in Chestnut Brook. Palynoflorule Locality No. 11 Formation: Middle Portland Formation. Stratigraphic position: 1100 ± 6 m above Hampden lava-flow unit, provided no major faults exist. Geographic location: Massachusetts; Hampden County, Agawam; cliff exposure on south bank of Westfield River, next to Bridge Street bridge. Palyniferous horizons: 6 beds of dark gray siltstone and shaly siltstone interbedded with brown-red siltstone and sandstone. Remarks: Cliff sequence measures 16 m thick, with an average interval of 2.3 m between palyniferous horizons. SYSTEMATIC PALEONTOLOGY Samples were prepared using standard HF-maceration and ZnCl2 separation. Glycerin jelly was used as mounting medium. Type specimens are deposited in the palynological collection at the Pennsylvania State University. The assignment of specimens to species previously described in the literature is based on 5 or more specimens. If less than 5 specimens are described, and comparison with an established species indicates affinity, our specimens are compared to that species. (The notation "cf." is used in such cases. However, "cf." also is used in some instances where more specimens are available but doubt as to exact affinity still exists.) A new species is proposed only if 10 or more specimens are described and if the specimens constitute a coherent group distinct from any already in the literature. If less than 10 specimens are described for a new form, it is given a reference number; for example, sp. 1 or Spore A. The median rather than the mean is given for unimodal and relatively non-skewed site ranges. For skewed or very broad size distributions, mean and mode are also given. Abundance of a species at a given locality is indicated by the following terms: rare (less than 1%), scarce (1-3%), common (4-10%), abundant (11-40%), and dominant (41-100%). A question mark indicates that rare, poorly preserved specimens are present, or that at a certain cited locality there is some doubt as to affinity of the specimens concerned. Location of type specimens is given with reference to an "X" scratched on the upper surface of the slide, per Traverse (1958). The terms intrastructure and intrastructural are used in descriptions for internal features of exines because "infrasculpture" confuses structure (internal) and sculpture (external), and "infrastructure'' has been used in other senses. Genus Anapiculatisporites Potonié & Kremp 1954 Type species: Anapiculatisporites isselburgensis Potonié & Kremp 1954 Anapiculatisporites cf. A. dawsonensis Reiser & Williams 1969 Plate 2, figure 9 Description: See Reiser and Williams, 1969, p. 3. Remarks: One specimen measures 44 pm in diameter. Locality: Portland Fm. 11 (rare). Genus Converrucosisporites Potonié & Kremp 1954 Type species: Converrucosisporites triquetrus (Ibrahim 1933) Potonié & Kremp 1954 Basionym: Verrucosisporites triquetrus Ibrahim 1933 Converrucosisporites cameronii (de Jersey 1962) Playford & Dettmann 1965 Plate 2, figures 3-8 Verrucosisporites cameronii de Jersey, 1962, p. 6, P1. 2, fig. 2, 3. Triletes hungaricus Venkatachala & Góczán, 1964, p. 210, P1. 1, fig. 1~14. Conbaculatisporites densus Mädler, 1964, p. 175, Pl. 1, fig. 9. Converrucosisporites cameronii (de Jersey) Playford & Dettmann, 1965, p. 136, Pl. 12, fig. 11-13. C. luebbenensis Schulz, 1967, P· 561, P1. 2, fig. 15-17; P1. 25, fig. 1. C. minor Pocock, 1970, p. 47, P1. 8, fig. 4, 5, 7. Emended description (more than 500 specimens): Isospores radial, trilete. Amb usually subaiangular with broadly rounded apices and sides slightly concave to slightly convex; amb sometimes subspherical. Laesurae extend almost to, but rarely reach, equaterial margin, and are usually bordered by narrow, slightly elevated lips. Kyrtomes prominently to weakly developed, not always present. Laesurae frequently bordered by arcuate folds concave toward proximal pole. Exine variably sculptured with a mixture of rounded or blunt-topped verrucae, large and small bacula, broad and narrow-based coni, rate clavae, and grana (grana comprising less than 50% of sculpture: compare description of Granulatisporites infirmus (Balme) comb. nov.). Sculpture comprehensive, although usually more scattered and reduced proximally. Distal and equatorial sculpture randomly spaced, distance between larger elements frequently greater than their diameter. Bases of sculptural elements usually joined to form a low, imperfect ridge system or incipient reticulum. When elements are large and closely spaced, many are fused into elongate, irregular tugae. Sculpture usually dominated by 1 type: verrucae 0.8-1.8 um in diameter and height, bacula 0.8-1.4 um high, coni 0.5-1.0 um high, and clavae about 1.0 um high. Exine 0.4-0.8 um thick (exclusive of sculpture), single-layered. Equatorial diameter (60 specimens) ranges from 33 um to 51 um (median 42 um), 90% of grains between 36 um and 49 um· Remarks: Specimens of this study compare very closely with C. cameronii from Australia, as well as with the spore of Clathropteris meniscoides Brongniart figured by Harris (1931, Pl. 18, fig. 3). The largest sculptural elements of the Australian form differ slightly from those of specimens from the Hartford Basin in being taller and conate rather than blunt and verrucate. The list of synonyms contains species that appear to be largely dominated by one type of sculpture: Triletes hungaricus with broadly based coni, Conbaculatisporites densus with closely spaced bacula, Converrucosisporites luebbenensis with verrucae and "truncae," and Converrucosissporites minor with broad verrucae. Numerous specimens from locality 8 closely compare with these species. Rather than divide what appears to be a natural species into 6 morpho-species, all of which would overlap in size and sculptural variation, 2 species are used that encompass the wide variation in sculpture with the least amount of overlap: Granulatisporites infirmus (Balme) comb. nov. and Converrucosisporites cameronii. Affinity: See discussion under affinity of G. infirmus. Localities: Portland Fm., 5, 11 (rare to scarce); Shuttle Meadow Fm. 1, 3, 4, 12 (rare to common); upper New Haven Arkose, 8 (abundant to dominant). Genus Convolutispora Hoffmeister, Staplin & Malloy 1955 Type species: Convolutispora florida Hoffmeister, Staplin & Malloy 1955 Convolutispora klukiforma (Nilsson 1958) Schulz 1967 Plate 1, figures 1-3 Description: Isospores radial, trilete. Amb usually subspherical, occasionally subuiangular. Distal and equatorial sculpture composed of large irregular verrucae, basally joined to form a large reticulum; lumina of reticulum 5-6 um in diameter. Verrucae generally aligned in 2 roughly perpendicular directions, one axial, the other equatorial. Proximal sculpture similar but reduced in height and broader, forming a flat contact area flanking the laesurae. Radius of laesurae 27-51% (median 34%) of equatorial diameter. Exine thickness 1.7-5.l um. Equaterial diameter (53 specimens) 44-75 um (median 59 um), 90% of specimens between 48 um and 71 um. Remarks: Contignisporites dunrobinensis (Couper 1958) Schult 1967 compares somewhat with this species, but axial sculptural lineation weakly expressed, equatorial ribs well developed. C. klukiforma may be conspecific with Striatella jurassica Mädler 1964. Species most prevalent in lake bed at Locality 1. Affinity: Schizaeaceous fern? Localities: Shuttle Meadow Fm., 1, 12 (rare). Genus Deltoidospora Miner 1935 Type species: Deltoidospora hallii Miner 1935, designated by Potonié 1956. Remarks: For generic synonymy, see Pocock, 1970, p. 27. Deltoidospora hallii Miner 1935 Plate 3, figure 6 Deltoidospora hallii Miner, 1935, p. 618, P1. 24, fig. 7, 8. D. cascadensis Miner, 1935, p. 618, Pl. 24, fig. 9-12. Tripartina variabilis var. glabra subvar, gradata Malyavkina, 1949, p. 50, P1. 7, fig. 10, 12, 14. Leiotriletes gradatus (Mal.) Bolkhovitina, 1953, p. 19, PI. 1, fig. 1~12; Pi. 7, fig. 10. Deltoidospora gradata (Mal.) Pocock, 1970, p. 28, P1. 5, fig. 2. Description: Isospores, trilete. Amb subtriangular in polar view, with sides nearly straight to moderately convex. Laesurae extend to, sometimes ending short of, equatorial margins. Laesurae may or may not be bordered by narrow raised lips; kyrtomes absent. Exine single layered, psilate, 0.8-1.0 um thick. Size range (14 specimens): 20-39 um in diameter (median 31 um). Remarks: The site range of our specimens completely overlaps the size ranges of species listed in the synonymy. Miner (1935) separated D. hallii and D. cascadensis on the basis of shape, but our specimens contain gradational types between spores with straight sides and ones with convex sides. The presence or absence of narrow, raised lips is not a significant difference -- lips may be variable in development as they are with the spore types referable to Clathropteris meniscoides Brongniart. Locality: Portland Fm., 11 (rare). Genus Dictyophyllidites Couper 1958 Type species: Dictyophyllidites harrisii Couper 1958 Dictyophyllidites paramuensteri n. sp. Plate 3, figures 4, 5, 8 Holotype: Plate 3, fig. 5, size 63 um; Slide BS2, coordinates 23.2 x 116.2, ref 16.1 x 119.6. Description: Isospores radial, trilete. Amb subtriangular with broadly rounded apices, and sides slightly concave to slightly convex. Laesura extends almost to, rarely reaching, equatorial margin, sometimes bordered by narrow, slightly elevated lips. Proximal side flattened, sometimes appearing slightly concave in lateral compressions; distal side strongly convex. In lateral compressiqns depth of spore equal to or greater than width. Kyrtomes usually weakly developed, sometimes strongly developed in polar compressions, frequently not evident in lateral comptessions. Exine psilate to vaguely punctate, single layered, 0.6-0.8 um thick. Equatorial diameter (252 specimens) ranges from 26 um to 110 um (median 68 um), 90% of grains between 41 um and 101 um. Remarks: The distribution curve is very broad and basically unimodal. Three slight peaks occur at 60 um, 68 um, and 85 um, but are not distinct enough to warrant the creation of more than one organ species. If this species were derived from one source, the extreme variation in size suggests some sort of environmental stress or adaptive significance. The size range completely overlaps the ranges of Dictyophyllidites harrisii Couper 1958 (36-56 um), D. mortonii (de Jersey) Playford & Dettmann 1965 (2845 um), Deltoidospora minor (Couper 1958) Pocock 1967 (26-56 um), D. australis (Couper 1358) Pocock 1970 (58-90 um), and the spores (26-47 um) of the fern Dictyophyllum nilssonii (Brongniart) Goeppert (quoted from Harris, 1944, by Couper, 1958). Dictyophyllum muensteri (Goeppert) Nathorst is the only Rhaeto-Liassic fern known to produce spores of the Dictyophyllidites type of a size similar to D. paramuensteri n. sp. The average size of these spores given by Harris (1931) is 65 um. Harris' illustration of one such spore (Harris, 1931, Pl. 14, fig. 3) measures 76 um at the given magnification. These 2 figures agree with the median of 68 um for D. paramuensteri n. sp.; hence, the implied relationship. Comparison of this species with Deltoidospora Miner 1935 reveals some close similarities in shape. Many of our specimens appear to lack kyrtomes, but cannot be separated by other characteristics from the specimens that possess them. The thin, flexible exine of this species contrasts with the more rigid exine generally found in Deltoidospora (synonym: Cyathidites). This species is most prevalent in the lake bed at Locality 1. Affinity: Probably Dictyophyllum muensteri, in part, but the size range could include some spores of other ferns. Localities: Portland Fm., 11?, 13 (rare); Shuttle Meadow Fm., 1, 12 (rare to scarce); upper New Haven Arkose, 8 (rare). Dictyophyllidites cymbatus Venkatachala & Góczán 1964 Plate 1, figures 11, 12 Remarks: Specimens from the Hartford Basin agree well with this species, particularly in the strong development of kyrtomes. Fifteen specimens range from 31 um-5l um (median 42 um) in equatorial diameter. Exine 0.5-0.9 um thick. Localities: Portland Fm., 5, 11 (rare). Dictyophyllidites harrisii Couper 1958 Plate 1, figures 7, 8 Remarks: Eight specimens from the Hartford Basin range from 37 um to 51 um (median 43 um) in equatorial diameter. This species is present in a mottled gray brown claystone, 5 stratigraphic m above the lake bed at Locality 1, and appears to be separable from D. paramuensteri n. sp at this horizon by its lighter color and more restricted size range. Dictyophyllidites spp. from the lake bed horizon do not show a similar bimodal distribution in size. Localities: Portland Fm., 5, 11 (rare); Shuttle Meadow Fm., 1, 12 (rare); upper New Haven Arkose, 8?. Genus Dictyotriletes Naumova 1939 Type species: Dictyotriletes bireticulatus (Ibrahim 1932) Potonié & Kremp 1955 (designated by Potonié & Kremp, 1955) Basionym: Sporonites bireticulatus Ibrahim 1932 in Potonié, Ibrahim, and Loose, 1932. Dictyotriletes Naumova 1939. Ischyosporites Balme 1957. Klukisporites Couper 1958. Dictyotriletes sp. 1 Plate 1, figures 5, 6 Description: Isospores, trilete. Amb subtriangular in polar view, strongly convex distally. Proximal face psilate; laesurae extend almost to equatorial margins. Distal side sculptured with a large, predominantly complete reticulum, which extends slightly proximal to equatorial margin at apices, generally ending at equatorial margin between apices. Muri of reticulum 2-7 um wide; lumina of reticulum 1.7-5.2 um in overall diameter. Exine doublelayered, 1.7-3.0 um thick, increasing to 4.0 um at apices; endexine thinner than ektexine, but not clearly delimited. Size (2 specimens): 44-48 um in polar-view diameter, one specimen measuring 51 um in depth. Remarks: This species is similar to Ischyosporites marburgensis de Jersey 1963 (recorded from the Liassic of Australia), which differs in having a slightly thicker exine and more uniform distribution and shape of lumina. Dictyotriletes sp. 1 has also been recovered from our Licking Run locality near Midland, Va., where several distinct species of Dictyotriletes are present (Pi. 1, fig. 4). Locality: upper New Haven Arkose, 8 (rare). PLATE 1 1 Convolutispora klukiforma (Nilsson) Schulz. Locality 1, 53 um. 2 C. klukiforma. Locality 1, 56 um. 3 C. klukiforma. Locality 1, 63 um wide. 4 Dictyotriletes sp. Locality on Licking Run, near Midland, Va., 70 um wide. 5 Dictyotriletes sp. 1. Locality 8, 44 umwide. 6 Dictyotriletes sp. 1. Locality 8, 48 umwide. 7 Dictyophyllidites harrisii Couper. Locality 1, 44 um. 8 D. harrisii. Locality 5, 42 um. 9 Murospora sp. 1. Locality 11, 37 um. 10 Murospora sp. 1. Locality 11, 26 um. 11 Dictyophyllidites cymbatus Venkatachala & Góczán 1964. Locality 5, 39 um 12 D. cymbatus. Locality 5, 43 um. 13 Foveosporites agawamensis n. sp (Holotype). Locality 11, 46 um (under oil). 14 F. agawamensis n. sp. Locality 11, 25 um (under oil). 15 F. agawanzensis n. sp. Locality 11, 41 um. 16 Gleicheniidites cf. G. nilssonii Pocock. Locality 5, 41pm. PLATE 2 1 Granulatisporites infirmus ( Balme) comb. nov. Locality 8; part of spore mass found in close association with Clathropteris meniscoides Brongniart; grain on far left, 42 um. 2 G. infirmus.Locality 8; S.E.M., 41 um. 3 Converrucosisporites cameronii (de Jersey) Playford 8 ( Dettmann. Locality 8, 46 um. 4 C. cameronii. Locality 8, 42 um. 5 C. cameronii. Locality 8; S.E.M., 41 um wide. 6 C. cameronii. Locality 8; S.E.M.; part of spore mass probably derived from Clathropteris meniscoides Brongniart; central grain, 37 um wide. 7 C. cameronii. Locality 8; S.E.M., 45 um. 8 C. cameronii. Locality 8, 56 um. 9 Anapiculatisporites cf. A. dawsonensis Reiser & Williams. Locality 11, 46 um. 10 Perotriletes cf. P. pseudoreticulatus Couper. Locality 11, 54 um. 11 Leptolepidites cf. L. major Couper. Locality 11; focus on sculpture to Fig. 12. 12 Leptolepidites cf. L. major. Locality 11, 54 um. 13 Todisporites rotundundiformis (Malyavkina) Pocock. Locality 11; 54 um. 14 T. rotundiformis. Locality 11; 46 um. 15 T. rotundiformis. Locality 11; 36 um. Genus Foveosporites Balme 1957 Type species: Foveosporites canalis Balme 1957 Foveosporites agawamensis n. sp. Plate 1, figures 13-15 Holotype: Plate 1, fig. 13, size 46 um; Slide AGA6-1, coordinates 52.8 x 111.2, ref 28.5 x 111.6. Description: Miospores, trilete. Amb subspherical, rarely subtriangular. Laesurae margins slightly thickened, sometimes as narrow conspicuous lips. Radius of laesura 35-46% of overall diameter of spore. Exine 0.6-0.8 um thick, proximally covered with densely to sparsely spaced pits, 0.3-0.4 um in diameter. Pits more closely spaced near laesura, decreasing in number equatorially; distal side generally lacking pits. Size range (11 specimens): 25-52 um (median 41um). Specific epithet refers to Agawam, Mass. Locality: Portland Fm., 11 (rare). Genus Gleicheniidites Ross 1949 Type species: Gleicheniidites senonicus Ross 1943 Gleicheniidites cf G. nilssonii Pocock 1970 Plate 1, figure 16 Description: See Pocock (1970, p. 32). Remarks: The uniqueness of this species necessitates some description: our specimens compare very closely with G. nilssonii, but have a slightly thicker exine (about 2.6 um thick between apices, decreasing to about 2.1 um at apices). A distal triangular area of thickening is delimited by 3 arcuate ridges, intersecting at the apices. Two specimens measure 41 um and 43 um in diameter. The small number of specimens permits only tentative assignment at this time. Locality: Portland Fm., 5 (rare). Genus Granulatisporites Ibrahim 1933 emend. Potonié & Kremp 1954 Type species: Granulatisporites granulatus Ibrahim 1933 Granulatisporites infirmus (Balme 1957) comb. nov. Plate 2, figures 1, 2 Concavisporites infirmus Balme, 1957, p. 21, Pl. 2, fig. 32, 33 (Basionym). Trachysporites asper Nilsson, 1358, p. 39, P1. 2, fig. 3. Granulatisporites cf. G. asper (Nilsson) Playford & Dettman, 1965, p. 135, Pl. 12, fig. 10. Emended description (more than 500 specimens): Isospores radial, trilete. Amb usually subtriangular with broadly rounded apices, and sides slightly concave to slightly convex; amb sometimes subspherical. Laesura extends almost to, rarely reaching, equatorial margin, usually bordered by narrow, slightly elevated lips. Kyrtomes prominently to weakly developed, not always present. Laesurae frequently bordered by arcuate folds concave toward proximal pole. Exine sparsely to densely sculptured with grana and small verrucae, mixed with a few bacula and coni, all of which are frequently joined at their bases by a low, imperfect ridge system or incipient reticulum. Sculpture is comprehensive, but usually proximally scattered and more reduced; sculpture ranges from predominantly small grana and verrucae, 0.1-0.8 um in diameter (always more than 50% of sculpture), to a mixture of grana, verrucae, bacula, and coni; occasionally either low coni or bacula dominate the larger sculptural elements. Exine 0.4-0.8 um thick (exclusive of sculpture), single layered. Equatorial diameter (100 specimens) ranges from 22 um to 62 um (median about 40 um), 30% of specimens between 33 um and 43 um. A few abnormal monolete and dilete (chevron) grains observed. Remarks: Playford and Dettmann (1965) considered Concavisporites infirmus Balme (equatorial diameter 35-46 um, mean 40 um) to be distinct from Trachysporites asper Nilsson (size 38-41 um) and their specimens, since C. infirmus possesses kyrtomes and a finely rugose sculpture. However, Balme's illustrations bear remarkable resemblance to specimens of the present study, most of which have some degree of kyrtome development. The illustrated specimen of T. asper (Nilsson, 1958, P1. 2, fig. 3) also has raised arcuate folds flanking the laesurae. The finely rugose sculpture in the illustrations of C. infirmus is not obvious (the sculpture appears to consist of discrete grana and verrucae); a basal incipient reticulum could be described as finely rugose, because closely spaced sculptural elements would be united to give an apparent lineation pattern (S.E.M. photomicrograph, P1. 2, fig. 2). The lack of obvious distinction between C. infirmus, T. asper, and our specimens favors uniting these forms under the earliest name, C. infirmus. This species conforms to the generic requirements of Granulatisporites ibrahim emend. Potonié & Kremp, which has priority over Concavisporites Pflug and Trachysporites Nilsson. Granulatisporites infirmus grades into Converrucosisporites cameronii (de Jersey) Playford & Dettmann in our samples, but for practical reasons the two forms are separated by gross sculptural differences. Granulatisporites subgranulosus (Couper, 1958) Fisher 1972 differs from G. infirmus by having a much thicker, double-layered exine (l.5-3.0 um thick). Affinity: Granulatisporites infirmus has been recovered by the first author from sporangia of Clathropteris meniscoides Brongniart at Locality 8. Associated spore masses in sporangial preparations contain C. cameronii. Some dispersed spore preparations from the megafossil plant layers are dominated by C. cameronii, while others are dominated by G. infirmus, although both forms are invariably present in the same preparation. Specimens of Equisetites spp. are the only other megafossils present in the plant bed. Probable roots and rhizomes of C. meniscoides and Equisetites are preserved in situ. Only one species of Clathropteris appears to be present, although it exhibits a wide range in frond site. Localities: Portland Fm., 5, 11 (rare to scarce); Shuttle Meadow Fm., 1, 3, 4, 12 (rare to common); upper New Haven Arkose, 8 (abundant to dominant). Genus Harrisispora Pocock 1970 Type species: Harrisispora mollis (Bolkhovitina 1956) Pocock 1970 Basionym: Adiantum mollis Bolkhovitina 1956 Remarks: See Pocock (1370, p. 38, 39) for generic description and generic synonymy. Harrisispora sp. 1 Plate 3, figure 3 Description: Miospores, trilete. Amb subtriangular with slightly convex sides. Radius of laesura about 30% of diameter of spore; laesura occupies for nearly its entire length a narrow zone or groove (1.6-3.0 um wide) of exinal thinning where ektexine and endexine apparently separate. Both endexine and ektexine participate in forming the laesura. Narrow, raised lips present on surface of endexine in groove. Adjacent to the narrow zone of thinning, a band of exine (67 um wide) is inaapunctate and thickened on each side of laesura. Exine double layered, psilate, about 1.4-1.6 um thick (exclusive of proximal area of thickening); ektexine 1.0 um thick, endexine 0.4-0.6 um thick. Size (1 specimen): 80 um in diameter. Affinity: Possibly ferns related distantly to Lygodium (Pocock, 1970). Locality: Portland Fm., 11 (rare). Genus Leptolepidites Couper 1953 Type species: Leptolepidites verrucatus Couper 1953 Leptolepidites cf. L. major Couper 1958 Plate 2, figures 11, 12 Remarks: Two specimens, measuring 46 um and 54 um, agree with L. major in size, and in having verrucae frequently in contact. However, the verrucae of our specimens are much smaller (1.0-2.5 um high; 1.0-3.0 um in diameter), agreeing in size and in number around the edge with Leptolepidites bossus (Couper) Schulz. Our specimens appear to be intermediate between L. major and L. bossus. Locality: Portland Fm., 11 (rare). Genus Murospora Somers 1952 Type species: Murospora kosankei Somers 1952 Murospora sp. 1 Plate 1, figures 9, 10 Description: Miospores, trilete. Amb subcircular to subtriangular in polar view; equatorial outline of cingulum generally parallels that of central body. Laesura extends almost to, sometimes reaching, equatorial margin of central body. Radial apices occasionally protrude beyond general outline of central body, producing a bulge on the cingulum. Laesura usually bordered by narrow, raised lips. Exine double layered, psilate. Ektexine extends over whole surface, including cingulum. Endexine generally restricted to central body, occasionally extending as lobes into, sometimes to outer margin of, cingulum. Exine 0.9-1.0 um thick, ektexine about 0.4 um thick, endexine about 0.6 um thick. Size range (7 specimens): 26-53 um in overall diameter (median 37 um); diameter of central body 22-41 um; width of cingulum 3-7.0 um. Locality: Portland Fm., 11 (rare). Genus Perotriletes Couper 1953 Type species: Perotriletes granulatus Couper 1953 Perotriletes cf. P. pseudoreticulatus Couper 1953 Plate 2, figure 10 Description: Miospores radial, trilete. Amb subtriangular in polar view. Laesura extends almost to equatorial margin, flanked by narrow, raised lips. Exine (exclusive of perispore) about 0.6 um thick; both proximal and distal faces sculptured with irregular, occasionally anastomosing, ridges, 1-2 um high. Perispore closely fitting, densely covered with small grana, 0.1-0.4 um high; perispore attached only at laesura margins. Overall site (2 specimens) 55-56 um, about 52-53 um in diameter, excluding perispore. Remarks: The presence of grana on the perispore might be sufficiently different from the type descrip- tion to warrant the creation of a new species. However, because of the small number of specimens, a tentative assignment is made until further information can be obtained. Our specimens differ significantly from other described species of Perotriletes. Locality: Portland Fm., 11 (rare). Genus Todisporites Couper 1958 Type species: Todisporites major Couper 1958 Todisporites rotundiformis (Malyavkina 1949) Pocock 1970 Plate 2, figures 13-15 Cyclina pseudolimbata var. rotundiformis Malyavkina, 1949, p. 53, P1. 9, fig. 13. Leiotriletes rotundiformis (Mal.) Bolkhovitina, 1956, p. 30, P1. 1, fig. 42b. Todisporites minor Couper, 1958, p. 135, P1. 16, fig. 9, 10. T. rotundiformis (Mal.) Pocock, 1970, p. 30, P1. 5, fig. 15. Description: See Couper (1958, p. 135). Remarks: Our specimens are very similar to T. minor, but have a thinner exine (0.4-0.7 um) and a distribution curve skewed toward smaller size. Measurement of 32 specimens from Locality 11 gives a size range of 29 um to 54 um (median 39 um, mean 40 um, mode 35 um). However, these differences are not significant. Pocock (1970) gives a size range of 38 um to 46 um, Couper (1958) gives 32 um to 50 um (mode 45 um), Malyavkina (1949) gives 30 um to 40 um. Some of our larger specimens are subtriangular in shape, but invariably these have major folds. Affinity: Couper (1958) compared T. minor with the spores of Todites princeps (Presl) Gothan, which are smaller (20-40 um) and have a mean (32 um) close to our mode. Localities: Portland Fm., 10, 11 (rare). Genus Verrucosisporites Ibrahim 1933 emend. Potonié & Kremp 1954 Type species: Verrucosisporites verrucosus Ibrahim 1933 Verrucosisporites cheneyi n. sp. Plate 3, figures 1, 2 Holotype: Pl. 3, fig. 1, size 73 um; Slide BS3, coordinates 40.5 x 122.7, ref. 56.2 x 126.2. Description: Isospores, radial, trilete. Amb circular to subcircular, proximal face less convex than distal side. Exine densely sculptured with verrucae and a few grana, 0.9-3.4 um high, 1.5-2.8 um wide. Verrucae sometimes in contact with one another, occasionally fused into larger elements. Radius of laesurae variable, 26-78% of equatorial diameter. Exine (exclusive of sculpture) approximately 0.8 um thick, single layered. Equatorial diameter (41 specimens) 58-83 um (median 70 um), 90% of grains between 59 um and 79 um. Remarks: This species compares closely with the Permian Verrucosisporites trisectus Balme & Hennelly 1956b, but differs somewhat in size range (V. trisecatus: 65-135 um), and in the degree of suture development (laesurae of V. trisecatus frequently extend onto distal surface of spore). The sculpture of both species is very similar. Species most prevalent in lake bed at Locality 1. Species named after Jane Cheney, director emeritus of the Children's Museum of West Hartford, Conn., in recognition of her efforts to preserve the natural history of the Connecticut Valley. Affinity: Osmundaceous fern? Localities: Portland Fm., 6, 11, 13 (rare); Shuttle Meadow Fm., 1 (rare); upper New Haven Arkose, 8 (rare). Spore A Plate 3, figure 7 Description: Miospore, trilete. Amb triangular in polar view, apices rounded. Radius of laesura about 30% of diameter of spore. Laesura open, bordered by a kyrtome consisting of narrow bands of exinal thickening. Distai side extended, equatorially strongly concave, forming triradiate fold centered at distal pole. Exine double layered; endexine 0.7 um thick; ektexine 1.0 um thick, loosely attached along triradiate fold. Size: 98 um in diameter. Locality: Portland Fm., 11 (rare). Genus Pilasporites Balme & Hennelly 1956a Type species: Pilasporites calculus Balme & Hennelly 1956a Pilasporites allenii Batten 1968 Plate 4, figure 11 Description: See Batten (1868, p. 638). Remarks: Our specimens agree closely in all respects with P. allenii Batten from the Lower Cretaceous of England, except in exine thickness, which is not a significant difference for the creation of a new species. Measurement of 72 specimens from Locality 8 gives a size range of 30 um to 54 um (median 39 um, mean 39 um), indicating a slightly larger average size than that of the type description (mean 36.7 um; Batten, 1968). Exine of our specimens is 0.5-0.8 um thick as compared with 1.25-1.75 um given by Batten. Slightly irregular distribution in the size-range curve of our specimens suggests more than one source plant, as is also indicated by megafossils. Affinity: Equisetites cf. E. sarranii Zeiller (abundant in layers containing P. allenii), as well as possibly two other species of Equisetites, present at Locality 8. Localities: Portland Fm., 5, 2 1 (rare); Shuttle Meadow Fm., 1 (rare in lake bed); upper New Haven Arkose, 8 (rare to scarce). Genus Araucariacites Cookson 1947 Type species: Araucariacites australis Cookson 1947 Remarks: Cookson (1947), following the suggestions of Erdtman (1947), created the "sporotype" names Granulonapites (± = form-generic name) and Araucariacites (± = organ-generic name), as alternative names to be combined with the epithet, australis. The 1972 I.C.B.N. restriction on the use of alternative names is retroactive only to January 1, 1953. Consequently, we consider subsequent attempts to validate one of the two generic names (Couper, 1953, Nilsson, 1958) unnecessary, since Cookson's monotypic species was validly published. Her sporotypes and sporomorphs are handled normally in all respects as genera and species, respectively. Our choice of the generic name, Araucariacites, is based on the fact that Cookson does not accept the name Granulonapites in her publication: wherever the generic concept is given a single name, that name is always Araucariacites. Araucariacites punctatus (Nilsson 1958) comb. nov. Plate 6, figures 14, 15 Granulonapites punctatus Nilsson, 1958, p. 70, Pl. 5, fig. 22. Description: Pollen grains inaperturate, spherical to oval, frequently folded. Exine 0.5-0.7 I~m thick, densely sculptured with small grana, 0.2 um or less in diameter. Size range (50 specimens): 28-63 um (median 44 um) in maximum width or diameter, 90% of specimens between 34.0 um and 56.8 um. Remarks: This species agrees in all morphological respects with Araucariacites australis Cookson, but has a smaller mean and smaller site distribution than generally recognized for A. australis. Couper (1953) reported grains of similar size under the name A. australis from Jurassic-lower Oligocene strata of New Zealand. Nilsson (1318) described one specimen (upon which A. punctatus is based) from the middle Keuper of Scania. This species is generally much more common than A. australis in our samples. Localities: Portland Fm., 5, 6, 7, 9, 10, 11, 13 (rare to scarce); Shuttle Meadow Fm., 1, 12 (rare). Araucariacites australis Cookson 1947 Plate 6, figure 13 Remarks: This species is distinguishable from A. punctatus (Nilsson) comb. nov. by its larger size and usually darker color. Range in size for 26 specimens: 54-94 um (median 73 um). Our specimens compare well in shape and in manner of folding to grains of Brachyphyllum mamillare Brongniart as illustrated by Couper (1958, Pl. 27, fig. 1, 2). Species is more frequently encountered in the Portland Fm. than in older strata. Some specimens assigned to this species may be unsplit examples of A. fissus Reiser & Williams. Localities: Portland Fm., 5, 11 (rare); Shuttle Meadow Fm., 1 (rare). Araucariacites fissus Reiser & Williams 1969 Plate 7, figures 6, 7 Description: See Reiser & Williams (1969, p. 17). Remarks: The size distribution (17 specimens) indicates a larger average size than that given by Reiser & Williams (1969), being 58-110 um (median 85 um) rather than 34-95 um (?mean 57 um) for the Australian forms. The indicated Early Jurassic age for A. fissus Reiser & Williams agrees well with the proposed age for the Portland Fm. This species becomes more frequent in younger strata. The largest forms are more common at Locality 5. Discussion of relationship of A. fissus to Triangulopsis: Our specimens (esp. P1. 7, fig. 7) from Locality 11 compare well in overall shape, manner of splitting, and size range to Triangulopsis discoidalis Döring 1961, recorded from the Upper Jurassic of Germany and western Canada. However, T. discoidalis apparently has a much thicker, double-layered exine, separable into an outer envelope and an internal body (Pocock, 1970, p. 74). Döring considered T. discoidalis to be microplanktonic, while Pocock's study of a large population of this species indicated morphological affinity with vascular plants. Döring considered Zonalapollenites to be synonymous with Triangulopsis, suggesting a similar basic structure. It is possible that A. fissus is related to Triangulopsis discoidalis, lacking only the development of a distinct endexine. Localities: Portland Fm., 5, 10, and 11 (rare). Genus Perinopollenites Couper 1958 Type species: Perinopollenites elatoides Couper 1958 Perinopollenites elatoides Couper 1958 Plate 4, figures 12, 13 Description: See Couper (1958, p. 152). Remarks: One specimen from~locality 11 clearly shows a pore (Pl. 4, fig. 12). Other specimens possess a highly folded "perine," which may simulate a crude reticulum (Pl. 4, fig. 13). Specimens from Locality 5 generally have a slightly smaller body, giving the appearance of a relatively larger "perine." Six specimens range in maximum diameter from 34 um to 52 um; diameter of central body 28 um-43 um. Localities: Portland Fm., 5, 11 (rare). Genus Callialasporites Sukh Dev 1961 Type species: Callialasporites trilobatus (Balme 1957) Sukh Dev 1961 (designated by Sukh Dev, 1961) Basionym: Zonalapollenites trilobatus Balme 1957 Zonalapollenites (in part) Pflug in Thomson and Pflug, 1953, p. 66, 67. Callialasporites Sukh Dev, 1961. Applanopsis Döring, 1961. Triangulopsis (in part) Doring, 1961. Pflugipollenites Pocock, 1962. Applanopsipollenites Levet-Carette, 1964. Description: See Sukh Dev, 1961, p. 48; Balme, 1957, p. 32. Remarks: Sukh Dev (1961) and Pocock (1970) discuss the synonymy of Zonalapollenites and Tsugaepollenites, and differences between these two genera and species now placed in Callialasporites. Callialasporites cf. C. dampieri (Balme 1957) Sukh Dev 1961 Plate 4, figure 10 Description: See Balme (1957, p. 32). Remarks: Three specimens range in overall diameter from 60 um to 77 um; body 56 um-59 um in diameter. Sculpture of saccus is finely granulate. Endexine of body is about 1.3 um thick. A slightly darkened area is present on the central part of the body. One specimen has saccus folded back on body, resembling a similar condition found in C. segmentatus (Balme) comb. nov. Affinity: Gamerro (1965) found grains of C. dampieri and C. trilobatus (Balme) Sukh Dev in male cones in organic connection with leaves of the gymnosperm Apterocladus lanceolatus Archangelsky. Locality: Portland Fm., 11 (rare). Callialasporites segmentatus (Balme 1957) comb. nov. Basionym: Zonalapollenites segmentatus Balme 1957, p. 33, Pl. 9, fig. 93, 94. Remarks: A formal transfer is made of this species here, although no specimens definitely referable to it were found. Callialasporites trilobatus (Balme 1357) Sukh Dev 1961 Plate 4, figure 4 Description: See Balme (1957, p. 33). Remarks: This species is larger than our specimens of Callialasporites dampieri, 7 specimens measuring 78 um to 104 um in overall diameter; diameter of body 46 um-84 um. Exine of body is 0.8-1.1 um thick. Saccus is densely covered with small grana, 0.2-0.8 um in diameter. The shape of the body ranges from oval to subtriangular, with 3 lobes well developed on forms with subtriangular bodies. Forms with oval bodies tend to possess sacci without indentations. Sukh Dev (1961) described a new species, Callialasporites monoalasporus, that closely resembles our forms with oval bodies. More specimens are needed to determine if a significant distinction exists between our forms resembling C. monoalasporus Sukh Dev, and C. trilobatus. In addition, C. monoalasporus may be conspecific with C. turbatus (Balme) Schulz 1967. Affnity: Gamerro (1965) illustrates grains of C. trilobatus and C. dampieri, as well as forms resembling C. monoalasporus, which he found associated with Apterocladus lanceolatus Archangelsky. Reiser & Williams (1963) found in their study forms of Callialasporites gradational between different species of this genus, suggesting that many of them are derived from one source plant. Locality: Portland Fm., 11 (rare). Genus Alisporites Daugherty 1941 emend. Nilsson 1958 Type species: Alisporites opii Daugherty 1941 Alisporites thomasii (Couper 1958) Nilsson 1958 Plate 4, figure 5-7 Pteruchipollenites thomasii Couper, 1958, p. 150, P1. 26, fig. 10, 11. Alisporites thomasii (Couper) Nilsson, 1958, p. 83, P1. 8, fig. 1. Description: See Couper (1958, p. 150). Remarks: Our specimens compare closely with A. thomasii in overall size and shape, as well as exine structure and sculpture. Twenty-one specimens range from 48 um to 77 um in overall breadth (median 69 um); breadth of body 20 um-46 um (median 36 um); length of saccus 36 um-57 um (median 40 um). Some specimens, not included in the above measurements, have bodies compressed more or less obliquely with pendantly attached sacci, producing forms suggestive of pollen of extant Pinus. More study is needed to determine whether these unique forms should be included in A. thomasii. Localities: Portland Fm., 5, 11 (rate). Genus Pityopollenites Reissinger 1950 emend. Jansonius 1962 Type species: Pityopollenites pallidus Reissinger 1950 Vitreisporites Leschik 1956 (imprint date, 1955). Caytonipollenites Couper 1958. Vitreisporites Leschik emend. Jansonius 1962. Remarks: Pityopollenites was validly published by Reissinger in 1950, and was monotypic at the time of publication. Vitreisporites, as originally described by Leschik in 1956, required a faint laesura and a size of the central body under 20 um. Jansonius (1962) removed these characteristics from the generic description, considering them either not convincingly demonstrated or too restrictive. For further discussion, see Dunay (1972). Pityopollenites pallidus Reissinger 1950 emend. Nilsson 1958 Plate 4, figures 2, 3 Pityopollenites pallidus Reissinger, 1950, p. 105), P1. 15, fig. 1-5. Vitreisporites signatus Leschik, 1956 (imprint date, 1955), p. 53, P1. 8, fig. 10. Caytonipollenites pallidus (Reissinger) Nilsson, 1958, p. 78, P1. 7, fig. 12-14. Description: See Nilsson (1958, p. 54). Remarks: Twenty-one specimens of this study range fiom 22 um to 33 um in overall breadth (median 27 um); breadth of body 7 um-15 um (median 27 um); length of saccus 14 um-22 um (median 17 um). Specimens from Locality 5 are generally smaller than those from Locality 11. Locatities: Portland Fm., 5, 11 (rare). Genus Platysaccus Naumova 1939 emend. Potonié & Klaus 1954 Type species: Platysaccus papilionensis Potonié & Klaus 1954 Remarks: Platysaccus was validly published by Naumova in 1939 (p. 355, fig. 1), although no type species was designated, which is not required by the I.C.B.N. before 1358. Potonié and Klaus (1954) designated their new species, P. papilionensis, as type, and emended the genus. Platysaccus cf. P. lopsiensis (Malyavkina 1961) Pocock 1970 Plate 4, figure 1 Podocarpus lopsiensis Malyavkina in Samoilovich et al., 1961, p. 130, Pl. 35, fig. 9a,b. Platysaccsus lopsiensis (Mal.) Pocock, 1970, p. 85, Pl. 18, fig. 6, 7. Description: See Pocock (1370, p. 85). Remarks: Two specimens compare closely with P. lopsiensis, measuring 56 um to 59 um in overall breadth (grains not completely expanded); breadth of body 26 um-27 um ; length of saccus 26 um-46 um. Locality: Portland Fm., 11 (rare). Genus Podocarpidites Cookson 1947 Type species: Podocarpidites elliptica Cookson 1947 (designated by Potonie', 1958) Remarks: This generic name was validly published by Cookson (1947) under circumstances similar to those described above under Araucariacites Cookson. Podocarpidites sp. 1 Plate 4, figure 14 Description: Pollen grains, bisaccate. Body of grain generally longer than broad; sacci strongly distally pendant, rarely expanded, usually highly folded. Distal surface traversed by an elongate sulcus, about 20 um long, flanked by basal attachment of sacci. When expanded, saccus measures about 36 um in length, 50 um in breadth. Proximal surface of body covered by a closely adhering cappa, 1.4-2.6 um thick, tightly folded to form a verrucose and rugose surface. Exine of body about 0.9 um thick. Sacci possess fine reticulum with lumina 0.2-0.3 um in diameter. Size of body (5 specimens): 40-46 um long; 31-34 um broad. Sacci generally about 30 um in diameter (folded). Overall breadth approximately 50-60 um, but highly dependent on shape of sacci. Remarks: Bisaccates are gehetally rare in the Shuttle Meadow Fm. However, they increase slightly in abundance in the lake bed and underclay at Locality 1. A similar florule from Licking Run near Midland, Va. (see Discussion), contains as much as 15% bisaccates in certain lacustrine horizons. Most bisaccates from floras dominated by Corollina meyeriana compare best with Podocarpidites and Pityosporites, although examples of Alisporites have been found. Generally poor preservation of most of the Shuttle Meadow bisaccates prevents description of several other types present. Localities: Shuttle Meadow Fm., 1, 12 (rare); upper New Haven Arkose, 8? Circulina-Corollina-Classopollis Nomenclatural problem: Circulina and Corollina were validly published as generic names by Malyavkina (1949), even though her generic and specific descriptions have been regarded as vague and inadequate for validation (Pocock and Jansonius, 1961; Reyre, 1969; Traverse, Ames and Spackman [CCFSP, 1974, in press: 38-103, 38-128]). Classopollis is generally regarded as the acceptable generic name, but was originally described as tricolpate by Pflug (1953). Subsequent emendation of Classopollis by Pocock and Jansonius (1961) and Couper (1958) largely corrected the description of this pollen type, but Chaloner (1962) and Pettitt and Chaloner (1964) provided the most accurate description, clarifying the structure of the exine with electron micrographs of well-preserved grains. Klaus (1960) emended the description of Circulina and Corollina, providing Hilfstypus -- better called neotype, which is admissible under 1372 I.C.B.N., Guide for Determination of Types, p. 76, for establishing taxonomic validity. However, Klaus chose to distinguish Circulina from Corollina, making Classopollis a junior synonym of Corollina. S. Samoilovich (Institute of Petroleum, Leningrad, USSR; pers. comm. to A. Traverse, 1973) has indicated that Malyavkina's type material was not preserved. She provided photographs (Pl. 4, fig. 9; Pl. 5, fig. 10) of "neotypes" (chosen by Zauer, supplied to Klaus, and chosen by him as Hilfstypus) for species of Circulina and Corollina, derived fiom the same Lower Jurassic stratigraphic horizon as studied by Malyavkina (from the same locality, a borehole 80 km to the west of the settlement of Sagiz). Even though the assignment of these "neotypes" by Zauer is subjective, it is perhaps the only way to obtain stability of nomenclature and to avoid personal preference for the name Classopollis. It is interesting to note that the "neotype" of Corollina compacta Malyavkina (Pl. 5, fig. 10) compares well with Classopollis in exinal structure, even though a well-developed distal pseudopore is not present, and equatorial striations are questionably present. Thus Klaus was apparently correct in making Classopollis synonymous with Corollina rather than with Circulina, It is further significant that the "neotype" of Circulina simplex Mal. closely resembles Malyavkina's illustrations, lacks a well-developed ring tenuitas (rimula) and distal pseudopore, and resembles specimens of Spheripollenites Couper and endoexinal bodies of Classopollis more than it does Circulina meyeriana (Mal.) Klaus, raising the question of the correctness of Klaus's emendation of Circulina. Reyre (1969) considers Circulina Mal. emend. Klaus as "an exceptional case within the genus Classopollis, corresponding to the coincidence of the limiting cases of the two characters, equatorial thickening and ornamentation of the intrastructure." He describes two species, Classopollis quezeli and C. kieseri, with a typical Classopollis intrastructure, but without equatorial striations or with only vaguely defined striations (pseudostriations). These two species closely resemble grains that have been erroneously assigned to Corollina meyeriana (Klaus) Venkatachala & Goçzán (Schulz, 1967, P1. 22, fig. 6) or Circulina meyeriana Klaus (Geiger and Hopping, 1968, P1. 4, fig. 32). In fact this confusion suggests that there is really no clear generic distinction between grains with a massive intrastructure and no equatorial thickening or striations, and grains with a structured exine, equatorial thickening, and striations. Consequently the only valid genetic characteristics are those that Circulina emend. Klaus and Classopollis have in common: frequent occurrence as tetrads, spherical shape, size range (18-60 um), proximal trilete mark, distal pseudopore, and subequatorial furrow or rimula. Since the intrastructure of the exine ranges from massive through various types (Reyre, 1969), equatorial striations vary in number and completeness, particularly in some species, for example, Classopollis torosus (Reissinger) Balme and C. kieseri Reyre, and because the amount of equatorial thickening is variable, even in a single species, these characteristics are best reserved for specific distinction. It is therefore clear that Circulina (as emended by Klaus) and Corollina should be combined into a single genus, with Classopollis as a junior synonym. Circulina Malyavkina may not be identical with Circulina Mal. emend. Klaus (the status of which is also confused). Corollina was monotypic when published, whereas Circulina was not. Thus, Corollina does not present problems of generic typification and is therefore chosen to embody all of the common generic characters. Genus Corollina Malyavkina, 1949 emend. Corollina Malyavkina 1949. Circulina Malyavkina (in part), 1949. PLATE 3 1 2 3 4 5 6 7 8 Verrucosisporites cheneyi n. sp. (Holotype). Locality 1, 73 um. V. cheneyi n. sp. Locality 1, 85 um wide. Harrisispora sp. 1. Locality 11, 80 um. Dictyophyllidites paramuensteri n. sp. Locality 1, 65 um wide. D. paramuensteri n. sp. (Holotype). Locality 1, 63 um. Deltoidospora hallii Miner. Locality 11, 27 um. Spore A. Locality 11, 98 um. Dicryophyllidites paramuensteri n. sp. Locality 1, 53 um. PLATE 4 1 Platysaccus ccf. P. lopsiensis (Malyavkina) Pocock. Locality 11, 58 um. 2 Pityopollenites pallidus Reissinger. Locality 5, 22 um. 3 P. pallidus. Locality 5, 26 um. 4 Callialasporites trilobatus ( Balme) Sukh Dev. Locality 11, 90 um. 5 Alisporites thomasii (Couper) Nilsson. Locality 5, 58 um. 6 A. thomasii. Locality 11, 63 um. 7 A. thomasii, Locality 11, 73 um. 8 Circulina simplex Malyavkina. Locality I, 25 um. 9 C. simplex Malyavkina (Neotype). Locality indicated under description, about 22 um. 10 Callialasporites cf. C. dampieri ( Balme) Sukh Dev. Locality 11, 75 um. 11 Pilasporites allenii Batten. Locality 8, 44 um. 12 Perinopollenites elatoides Couper. Locality 11, 41 um. 13 P. elatoides. Locality 11, 44 um. 14 Podocarpidites sp. 1. Locality 1, 58 um. Classopollis Pflug 1953. Classopollis Pflug emend. Couper 1958. Corollina Mal. emend. Klaus 1960. Circulina Mel. emend. Klaus 1960. Classopollis Pflug emend. Pocock & Jansonius 1962. Monilapollis Chang 1963. Pagiophyllumpollenites Chang 1963. Classopollis Pflug emend. Reyre 1969. [Also the species listed below referred to "Pollenites" by Thiergart and by Reissinger.] Generic type: Corollina compacta Malyavkina, 1949, p. 124, Pl. 46, fig. 10. (Liassic). Holotype: Malyavkina, 1949, P1. 46, fig. 10, 11. Neotype: Plate 5, fig. 10. Designated by Zauer, photograph supplied by Samoilovich, 1973. Stratigraphic position and locality of neotype: Borehole Kozhegali K36, 466.8 m-471.9 m, Lower Jurassic, 80 km west of Sagiz near Caspian Sea. Species assigned to genus: Circulina funifera Malyavkina, 1949, p. 123, P1. 46, fig. 6; BennettiteaePollenites reclusus Thiergart, 1949, p. 11, P1. 2, fig. 14, 15, P1. 3, fig. 6; Pollenites torosus Reissinger, 1950, p. 115, P1. 14, fig. 20. Emended generic description: Pollen grains distally pseudoporate, proximally trilete, spherical to ovoid, or compressed, acorn shaped; occur as tetrads or isolated grains; exine two layered, with distinct endexine and ektexine. Endexine comprises a spherical, internally separate body, which frequently may be poorly developed and difficult to distinguish, or well developed, sometimes occurring as separate bodies from rupnued grains. Ektexine (modified from Reyre, 1969) composition variable on different parts of the grain; composed of inner structured layer, comprising inuastructure, and outer tegillum, which is continuous over entire grain and covered with outer sculpture uniformly distributed over whole surface. Intrastructure massive, alveolate, punctate, vermiculate or pseudovermiculate, reticulate or pseudoreticulate, but can be heterogeneously organized: reduced, or thickened, in parts of the grain; reduced at distal pole (pseudopore) or only along circular line surrounding it, reduced along a subequatorial furrow (ring tenuitas or rimula) on distal side of equator, and sometimes reduced at proximal pole (triangular area). Intrastructure generally thidcened in equatorial zone on proximal side of furrow or rimula, but never thinner than ektexine proximai to equatorial zone or belt; intrastructurai elements within this equatorial zone may or may not be organized into more or less continuous circumequatorial striations; they are absent if intrastructure of this zone is massive. Remarks: Endexinal body may or may not possess a poorly developed trilete mark; it infrequently possesses a distal area of thinning or tenuitas. The neotype of Circulina simplex Malyavkina (our P1. 4, fig. 9) and illustrations by Malyavkina (Pl. 46, fig. 7-3) strongly resemble separated endexinal bodies of Corollina, and therefore do not possess sufficient characteristics to place C. simplex in Corollina. Circulina funifera Mal. (Pl. 46, fig. 6), on the other hand, appears to possess a subequatorial furrow (border sharply differentiated from the body, narrow, double), and exinal intrastructure (border covered with fine, radially directed, straight lines), which possibly place it in Corollina. Corollina torosus (Reissinger 1950) Klaus 1960 emend. Plate 5, figures 2-3, 14 Pollenites torosus Reissinger, 1950, p. 115, Pl. 14, fig. 20. Classopollis classoides Pflug, 1953, Fig. 4, J-M, P1. 16, fig. 20-25, 229-37; emend. Pocock & Jansonius, 1961, p. 443, P1. 1, fig. 1-3. Classopollis cf. C. torosus (Reissinger) Balme, 1957, p. 37, Pl. 11, fig. 114-119. C. torosus (Reissinger) Couper, 1958, p. 156, Pl. 28, fig. 2-7; Nilsson, 1958, p. 74, Pl. 7, fig. 68. C. torosus (Reissinger) Balme; Chaloner, 1962, p. 13, fig. 2, Pl. 11, fig. 1, 2; emend. Burger, 1965, p. 65, P1. 1, fig. 1, 2. Monilapollis hsui Chang, 1963, p. 435, 439, P1. 1, fig. la-lc, 2a-2c. "Classopollis torosus" Pettitt & Chaloner, 1964, P1. 1, fig. 1-5. Emended description: Isolated pollen grains or occurring as atrads; spherical to ovoid, or acorn-shaped; diameter in equatorial view (100 grains): 18-41 um (median 32 um), 90% of grains between 24 and 37 um (Couper, 1958, notes range 24-46 um, mode 32 um). Diameter in polar view 24-37 um (diameter of subequatorial furrow 83-88% of overall diameter). Distal pseudopore variable in size, even for grains of same size: 4.3-8.5 um in diameter, frequently distorted and widened in equatorial view. Proximal triangular area of thinning, which sometimes contains trilete mark, 6.8-13.6 um high. Exine two layered; ektexine divisible into outer tegillum, continuous over whole surface of grain, and inner structured layer, which is thickened in equatorial region to form a band or belt. Endexine variably distinct to indistinct, spherical, frequently scabrate, occasionally with a small proximal trilete mark. Structure of equatorial band ranges from pseudoreticulate, to vaguely striate with aligned intrastructural columellae (pseudostriations), to distinctly striate, with columellae fused to form more or less parallel bands, the number of striations frequently varying from one part of band to another. Sometimes combinations of grains with all degrees of striation development occur in same tetrad. When striations are well developed, maximum number per grain ranges from 6 to 10. Width of equatorial band, which frequently ranges from 5 um to 13 um at its maximum, usually decreases in one area to 4-10 um, and striations decrease in number to 4 or 5 through anastomosis or disintegration of more proximal striations into discrete intrastructural elements. Exine thickness varies with grain size, and presumably also with preservation; it is thickest in equatorial band: 1.3-2.4 um (Couper, 1958, records 3.0 um), decreasing gradually toward proximal pole: for example, 2.l um down to 0.9 um; thickness on distal side of equaterial furrow (range 0.7-1.7um) generally slightly less than that proximal to equatorial band. Intrastructure of ektexine mostly pseudoreticulate, with positive elements largest on proximal side of equatorial furrow, particularly if equatorial band is pseudoreticulate; pseudoreticulum on distal side of grain usually finer in pattern, occasionally becoming punctate. Sculpture of tegillum variable (perhaps owing to preservation); light microscope indicates roughened surface; S.E.M. indicates microscabrate sculpture, mixed with microverrucae and microconi (Pl. 5, fig. 14; see also Pettitt and Chaloner, 1964, p. 614); sculpture distinctly less well developed than in species figured by Reyre (1969), perhaps less well developed than in forms described under Classopollis torosus (Reissinger) Couper emend. Burger 1965. Remarks: The emended description of Classopollis classoides Pflug by Pocock and Jansonius (1961) is not adequate and conflicts with our present knowledge of the exine structure of this genus. We find no distinction between C. classoides and C. torosus, these species agreeing in exinal structure and variation in striation development (Pocock and Jansonius, 1961, Pl. 1, fig. 1-9). The size ranges overlap widely, but the mean equatorial width for C. classoides (29 um) is somewhat smaller than that of C. torosus (32 um). However, Harris (1957) noted a still smaller mean in his samples (25 um) for grains placed by Pettitt and Chaloner (1964) in C. torosus. These differences may result in part from preservational states and preparation techniques. The mean diameter for C. torosus from Locality 11 is 32 um, while it is 25 um from Locality 5. Table 1 gives data for relative abundance of C. torosus that suggest the existence of two separate "Circumpolles" populations, intermixing at Locality 5. The morphology of these two populations does not permit their distinction in a mixed sample, as the ranges of variation widely overlap. Therefore, only one form-species is described, although two different source plants may be indicated. Affinity: Hirmerella muensteri (Schenk) Jung 1968, possibly Pagiophyllum connivens Kendall 1948. Localities: Portland Fm. 5, 6, 7, 9, 10, 11 (dominant); Shuttle Meadow Fm. 1, 3, 4, 12 (rare to scarce); upper New Haven Arkose, 8 (scarce). Corollina itunensis (Pocock 1962) comb. nov. Plate 5, figure 1 Classopollis itunensis Pocock, 1962, p. 71, Pl. 11, fig. 176, 177; P1. 12, fig. 178 (Basionym). Description: See Pocock (1970, p. 104). Table 1. Approximate percentages of major Corollina species in Hartford Basin (Based on Corollina counts only. Most of palynoflorules counted were at least 90% Corollina.) Localities arranged in stratigraphic order, youngest at right: N.H.A. = New Haven Arkose; S.M.F. = Shuttle Meadow Formation; P.F. = Portland Formation; + = present, but less than 1%. STRATIGRAPHIC POSITION INTRASTRUCTURAL TYPE Corollina SPECIES N.H.A. 95% 2.5% 2.5% S.M.F. 97% 1% + No. 12 97% + 3% No. 5 2% 4% 58% No. 11 1% + 23% 36% 76% C. meyeriana C. murphyi n. sp. C. torosus C. torosus massive to micropunctate vermiculate (non-striate) pseudoreticulate to punctate (non-striate) pseudoreticulate to punctate (pseudostriatestriate) Remarks: Our specimens compare very closely in structure to C. torosus, but differ significantly in size and color or transparency. They are very light in color, pale yellow. Equatorial-view diameter (16 specimens) from 44 um-57 um (median 48 um); depth from pole to pole 36-53 um. These specimens appear distinct from C. torosus, because they have been found as tetrads and in a fragment of a pollen mass that contained only grains of the above description. The lighter color is not the result of thinning of the exine: endexine 1.3 um thick; ektexine 0.3-0.4 um proximally, 1.3 um equatorially, and 0.9 um distally. This species is common to the Canadian Jurassic, first occurring in zone J1 3 of Pocock (1970), which is equivalent to the upper part of the Lower Jurassic. Locality: Portland Fm., 11 (rare). Corollina meyeriana (Klaus 1960) Venkatachala & Goçzán 1964 Plate 6, figures 3-9 Circulina meyeriana Klaus, 1960, p, 165, P1. 36, fig. 57-60. Corollina meyeriana (Klaus) Venkatachala & Goçzán, 1964, p. 217, Pl. 3, fig. 1-15. Gliscopollis meyeriana (Klaus) Venkatachala, 1966, p. 93. Description: Isolated pollen grains or occurring as tetrads; spherical to ovoid, or acorn shaped, occasionally subtriangular in polar view; diameter in polar view (50 specimens): 20-31 um (median 26 um) (Klaus recorded "33 um (35 um) 40 um"); diameter in equatorial view (12 specimens): 22-28 um (median 26 um); distal pseudopore diameter 7-12 um; subequatorial furrow present; height of proximal triangular area of thinning 7-12 um; trilete radii each about 1.7 um long. Exine usually single layered, although occasionally an endexine body is present; exine 0.4-1.6 um thick, sometimes thickened in equatorial region (1.0-1.6 um thick), gradually thinning toward proximal pole; intrasaucture massive to micropunctate, frequently punctate if equatorial region thickened; sculpture roughened to microscabrate. Remarks: The size range given by Klaus is much larger than and does not overlap with, the grains from the Hartford Basin. However, de Jersey (1971) noted a size range for this species in the Ripley Road Sandstone and Helidon Sandstone of Australia of "21 um to 39 um in equatorial diameter." Occasionally proximal or distal parts are found separately, suggesting separation along the subequatorial furrow, but this does not necessarily indicate the operculate condition proposed by Venkatachala and Goçzán (1966). Kendall (1943) gives size range for pollen associated with Brachyphyllum scotii Kendall as 30-40 um in diameter. Affinity: Brachyphyllum scotii Kendall 1949. See Barnard, 1968, p. 175, P1. 1, fig. 3, 6, for further information on pollen associated with B. scotii. Localities: Portland Fm., 5, 6, 7, 9, 10, 11 (rare to common); Shuttle Meadow Fm., 1, 3, 4, 12 (dominant); upper New Haven Arkose, 8 (dominant). Corollina murphyi n. sp. Plate 5, figure 11-13, 1T, 16 Holotype: Plate 5, fig. 11, size 30 um; Slide PL9, coordinates 37.7 x 109.6, ref. 52.4 x 111.3. Description: Pollen grains, spherical to ovoid, occasionally subtriangular in polar view; diameter in polar view (30 specimens) 28-42 um (median 34 um); diameter in equatorial view (6 specimens): 30-37 um; distal pseudopore diameter 6.5-9.5 um; subequatorial furrow present. Width of proximal area of thinning 5.2-10.6 um: area not always observable, rarely triangular, frequently irregular in shape. Exine two layered, 0.9-3.4 um thick; endexine distinct to indistinct, spherical, psilate, thin (0.3 um thick), frequently folded or missing in ruptured grains; ektexine 1.8-3.4 um thick in equatorial region, without striations to possessing 1 or 2 irregularly widened, imperfect circumequatorial bands forming a belt about 4.5 um wide; intrastructure vermiculate to imperfectly pseudoreticulate, formed by broad irregular islands of structural thickening, which are separated by narrow anastomosing lines of exinal thinning; sculpture indistinct to microscabrate. Remarks: Intrastructural elements broader and mote massive than in C. torosus. Species named after Sister Mary Ellen Murphy, Department of Chemistry, Saint Joseph College, West Hartford, Conn., who is contributing important organic geochemical evidence for sediments of the Hartford Basin. Localities: Portland Fm., 5, 11 (scarce to common); Shuttle Meadow Fm., 1, 12 (scarce); upper New Haven Arkose, 8 (scarce). Corollina simplex (Danzé-Corsin & Laveine 1963) comb. nov. Plate 6, figure 10 Basionym: Classopollenites simplex Danzé-Corsin & Laveine 1963 Classopollenites simplex Danzé -Corsin & Laveine, 1963, p. 106, P1. 11, fig. 7, 8. Classopollis simplex de Jersey & Paten, 1964, p. 12, P1. 7, fig. 4-6, non Danzé-Corsin & Laveine 1963. C. simplex (Danzé -Corsin & Laveine) Reiser & Williams, 1969, p. 16, P1. 6, fig. 15. Excluded (homonymous) species: Classopollis simplex Reyre, 1969, p. 312, P1. 54, fig. 3-5. Description: See Reiser and Williams( 1969, p. 16). Remarks: Specimens from the Hartford Basin range in polar view diameter from 17 um-28 um, which agrees well with the range given by de Jersey and Paten (1964). However, Danzé-Corsin and Laveine (1963) reported a range of 20-40 um. The generic name Classopollenites Danzé-Corsin & Laveine 1363 is a superfluous substitute for Classopollis Pflug 1953, and is, hence, illegitimate. Furthermore, Classopollis simplex de Jersey & Paten 1964, although validly published, is a homonym, and is also conspecific with C. simplex Danzé-Corsin & Laveine. Classopollis simplex Reyre is distinctly different and is therefore an illegitimate homonym. It should be noted that Corollina simplex is probably not a natural species, since it has been found as anomalous grains in tetrads of both C. meyeriana and C. torosus. C. simplex grades into forms of C. meyeriana with a slightly thickened equatorial band, as well as into small compact forms of C. torosus. However, this does not preclude use of the concept of C. simplex. Localities: Portland Fm., 5, 7, 9, 11 (rare); Shuttle Meadow Fm., 1, 3, 12 (rare to scarce); upper New Haven Arkose, 8 (rare). Corollinoid? pollen Plate 6, figures 11, 12 Description: Two unusual palynomorphs resembling Corollina possess a circumequatorial saccus or pseudosaccus. Both grains have p'roximal triangular area of exinal thinning, equatorial furrow of exinal thinning, and distal pseudopore (although not obvious on specimen in P1. 6, fig. 12). One specimen (Pl. 6, fig. 11) has massive intrastructure and narrow flange or saccus, the other (Pl. 6, fig. 12) an intrapunctate structure and expanded, well-developed saccus. These differences suggest that they may not be conspecific. Saccus or pseudosaccus on each grain appears to be attached at equatorial furrow. Exine equatorially thickened (about 1.7 um thick), appearing massive in optical section. Size (2 specimens): 42 um in overall diameter; body 31-34 um in diameter; diameter of pseudopore 9-11 um; height of triangular area of thinning about 14 um; saccus or pseudosaccus extends 4-6 um beyond perimeter of body. Remarks: These grains appear to have a close affinity with Corollina.They show different degrees in development of the saccus or pseudosaccus, suggesting a developmental trend. The presence of this circumequatorial feature is striking if these grains were derived from the "Circumpolles." More specimens of these forms in these or other sediments are needed to permit a more complete study. Locality: Portland Fm., 11 (rare). Genus Circulina Malyavkina 1943 emend. Type species: Circulina simplex Malyavkina 1949 Circulina Malyavkina (in part), 1949. Species assigned to genus: Circulina simplex Malyavkina, 1949, p. 224, P1. 46, fig. 7-9. Excluded generic concept: Circulina Malyavkina emend. Klaus, 1960. Excluded species: Circulina funifera Malyavkina, 1949, p. 123, 124, P1. 46, fig. 6. Emended description: Pollen grains inaperturate, oval to circular, scabrate, punctate, or smooth. Exine single layered, lacking internal structure of Corollina. Remarks: This genus is intended for small inapertutate grains that do not have the germinal structures observed in Spheripollenites Couper 1958 or Exesipollenites Balme 1957, and that may be isolated endexinal bodies of Corollina Malyavkina 1943 emend. Circulina simplex Malyavkina 1943 emend. Plate 4, figures 8, 9 Holotype: Malyavkina (1949, P1. 46, fig. 7-9). Neotype: Plate 4, fig. 3, in this paper, a specimen earlier designated by Zauer, photograph supplied by S. Samoilovich, 1973 (for geographic and stratigraphic location, see Corollina compacta neotype, in earlier discussion of "Circulina-Corollina-Classopollis Nomenclatural Problem"). Emended description: Pollen grains inaperturate, oval to circular, punctate to smooth, occasionally scabrate. Exine single layered, sometimes folded to give the impression of an eccentric double wall. Exine about 0.7 um thick. Diameter of grains about 30 um. Localities: Portland Fm., 5, 7, 9, 10, 11, 13 (rare); Shuttle Meadow Fm., 1, 3, 12 (rare); upper New Haven Arkose, 8 (rare). Genus Camerosporites Leschik 1956 emend. Scheuring 1970 Type species: Camerosporites secatus Leschik 1956 Camerosporites reductiverrucatus n. sp. Plate 7, figures 1-5 Holotype: Plate 7, fig. 1, size 54 um; Slide AGA1-22, coordinates 12.6 x 111.4, ref. 56.7 x 110.8. Description: Pollen grains, spherical to ovoid, or acorn shaped, occasionally present as tetrads. Triradiate fold sometimes present at proximal pole. Equatorial furrow or ring tenuitas occasionally well developed; proximal part of grain often found without distal part. No distal tenuitas observed. Grains frequently folded. Exine of proximal part of grain slightly thicker than distal part;·proximal part generally sculptured with low, broad verrucae, more closely spaced around equator, forming equaterial band 4-12 um wide (mode 8.5 um). Distal part of grain usually psilate, but occasionally possesses a few scattered low vetrucae arranged around distal pole. Sculpture of proximal part of grain varies considerably from an abundance of low verrucae, simulating an equatorial band, to almost no sculpture, with a few verrucae restricted to equatorial region. Exine single-layered?; exine of distal part of grain 0.8-0.9 um thick, occasionally intrascabrate; exine of proximal part of grain 0.8-1.2 um thick. Verrucae 2.5-4.5 um wide, irregular in shape, about 0.4 um in height. Size range (50 specimens): 39-70 um in overall diameter (median 56 um); 90% of grains between 48 um and 65 um. Remarks: This species is larger than other species of this genus reported from the Keuper. Our highly sculptured forms resemble Camerosporites secatus Leschik, but lack the thick verrucae of this species. Other sparsely sculptured forms show a tendency of our species to lose the characteristic sculpture of the genus, and these would probably not be placed in Camerosporites if they did not intergrade with highly sculptured forms. This species suggests that at least one source plant for Camerosporites may have persisted into the Jurassic. Locality: Portland Fm., 11 (rare). Genus Cycadopites Wodehouse 1933 ex Wilson & Webster 1946 Type species: Cycadopites follicularis Wilson & Webster 1946 Cycadopites Wodehouse 1933. Entylissa Naumova 1933. Cycadopites Wodehouse ex Wilson & Webster 1946. Monosulcites Cookson 1947. Ginkgocycadophytus Samoilovich 1953. Ginkgoretectina Malyavkina 1953. Monosulcites Cookson ex Couper 1953. Entylissa Naumova ex Potonié & Kremp 1954. Cycadopollenites Danzé-Corsin & Laveine 1963. Ginkoretectinapollenites Danzé-Corsin & Laveine 1963. Remarks: For further synonymy, see Jansonius (1962, p. 80). Potonié (1958) separated many of the genera in the proposed synonymy by the shape of the sulcus or by the degree of overlap of sulcus margins. Subsequent authors have placed many of these genera in synonymy, but they have continued to separate Monosulcites and Cycadopites on the basis of sulcus shape. However, variation in sulcus shape and margin overlap probably reflect differences in orientation and compression during fossilization, as well as variation in the original shape of the grains. Features of the sulcus should therefore be used only as an aid in distinguishing species, but caution should be exercised in assigning grains to different genera on this basis alone. Cycadopites andrewsii n, sp. Text-figure 4; Plate 8, figures 17-21 Holotype: Plate 8, fig. 20; size 37 um long, 25 um wide; Slide CLRZ-1, coordinates 34.4 x 120.9, ref. 26.1 x 121.5. Description: Fusiform to ovoid pollen grains, monosulcate; longitudinal ends rounded to bluntly pointed; sulcus narrowest at distai pole, usually widening toward longitudinal extremities. Exine distally smooth, proximally sculptured with closely spaced verrucae, which form a very distinct pseudoreticulum. Verrucae 0.3 um x 0,3 um to 1.7 um x 2.6um wide, broadest at proximal pole, decreasing in diameter, to a lesser degree in height, toward extremities and equatorial margin. "Lumina" of pseudoreticulum decrease in size away from proximal pole, usually TEXT-FIGURE 4. Diagram of polar cross-section of pollen grains of Cycadopites andrewsii n. sp., showing relationship of ektexine and endexine and distribution of sculpture. disappearing at, or slightly distal to, equatorial margin. Exine double layered, ektexine (exclusive of sculpture) about 0.3-0.4 um thick, sculpture 0.1-0.4 um high, endexine about 0.2 um thick. Endexine apparently loosely attached except at sulcus margins, sometimes separating, in part or completely, as a distinct inner envelope. Lips of sulcus conform to area of apparent ektexinal and endexinal fusion; lips widest at distal pole (1.7-2.6 um wide), tapering to extinction at longitudinal extremities. Size range (30 specimens): length 34-46 um (median 38 um); width 20-30 um (median 22 um). Ratio of width to length 1:1.2-1:2.4 (median 1:1.7). Remarks: Several monosulcate species described in the literature compare with C. andrewsii n. sp., but apparently have an intrareticulum rather than an external pseudoreticulum; for example, Monosulcites rhaetoliassicus Madler 1964 (Rhaeto-Liassic), Cycadopites reticulatus (Nilsson 1958) comb. nov. (Lower Jurassic), Ginkgoretectina couperi (Pocock 1962) Pocock 1970 (Upper Jurassic), and G. ferrei Pocock 1970 (Upper Jurassic). Species named after Professor Henry N. Andrews, paleobotanist, University of Connecticut, Storrs. Locality: upper New Haven Arkose, 8 (rare to scarce). Cycadopites deterius (Balme 1957) Pocock 1970 Plate 8, figures 10-12 Entylissa deterius Balme, 1957, P· 29, Pl. 6, fig. 75-77. Cycadopites gracilis Sah & Jain, 1965, p. 282, P1. 7, fig. 146-149. Description: Pollen grains monosulcate, elliptical. Sulcus frequently widest at extremities, narrower at distal pole. Sulcus margins sometimes parallel one another for much of grain length, occasionally overlap to a lesser or greater degree. Lips generally absent. Exine single layered, 0.5-0.9 um thick, frequently slightly thicker at longitudinal extremities. Sculpture usually psilate distally, scabrate to psilate proximally, with larger grana frequently concentrated on proximal side at extremities. Grain color ranges from dark to light, depending on state of preservation. Size range (23 specimens): 30-60 um long (median 50 um); 14-35 um wide (median 24 um). Ratio of width to length 1:1.5-1:3.0 (median 1:2.2). Remarks: The size range of our specimens is slightly larger than that given by Balme (1957) for Entylissa deterius (41-60 um long), but the average size is the same. The sculpture on E. deterius is psilate, ranging to scabrate on some of our specimens. Cycadopites gracilis is included in the synonymy because it contains a combination of features found in our specimens: scabrate sculpture, overlapping sulcus margins, and a similar, but slightly smaller, size range (length 32-44 um). Together, both E. deterius and C. gracilis cover the range of variations found in our specimens, but cannot be distinguished in out samples as separate organ species. Cycadopites carpentieri (Delcourt & Sprumont) Singh 1964, per the description in Couper (1958), compares very well to this species in overall shape, exine thickness, and features of the sulcus. Although the size range of C. carpentieri overlaps that of C. deterius, the median size of C. carpentieri is about 10 um larger, suggesting that Couper's specimens are a distinct form. The grains illustrated by Delcourt & Sprumont (1955) show that the specimens upon which C. carpentieri was based may have had a significantly thicker exine than our specimens. Specimens from the Shuttle Meadow Fm. Are slightly more fusiform (median ratio of width to length 1:2.6) than those from the Portland Fm. and fall into the lower part of the size range of specimens from the Portland: length 31-46 um (median 40 um); width 14-19 um (median 17.0 um). The sulcus of Shuttle Meadow specimens trends to be more linear in shape, sometimes gaping only at one end, or rarely at the distal pole. Localities: Portland Fm., 5, 7, 10, 11, 13 (rare to scarce); Shuttle Meadow Fm., 1 (rare). Cycadopites durhamensis n. sp. Plate 8, figures 13-15 Holotype: Plate 8, fig. 13, size 26 um long, 12 um wide; Slide LTG-1, coordinates 40.0 x 124.0, ref. 50.7 x 125.7. Description: Pollen grains monosulcate, fusiform or elliptical. In polar comptessions longitudinal extremities generally bluntly pointed to rounded; in lateral compressions ends sometimes pointed, nipple like. Sulcus generally straight, margins frequently overlapping for much of the length of grain. Occasionally one end of sulcus expanded and open; rarely both ends of sulcus expanded on same grain. Infrequently grains ate compressed, with sulcus gaping widely at distal pole: such grains frequently show rents. Lips generally not developed, but occasionally sulcus margins at distal pole thickened. Exine single layered, 0.5-0.7 um thick, possibly slightly intrascabrate. Sculpture psilate. Size range (21 specimens): 22-31 um long (median 27 um); 9-16 um wide (median 13 um). Ratio of width to length 1:1.8-1:3.0 (median 1:2.1). Remarks: This species ranks among the smallest of monosulcates reported from the Mesozoic. It differs from Cycadopites minimus (Cookson) Pocock 1970 per description of Couper (1958, p. 157, Pl. 26, fig. 23-25), in being slightly smaller (smaller average size), and in having a more fusiform outline (ratio of width to length 1:2.1 rather than 1:1.4 as in Couper's specimens). The tendency of the sulcus margins to overlap in our specimens also serves to distinguish C. durhamensis from C. minimus. Our specimens compare well in shape and size with the Rhaeto-Liassic Ginkoretectinapollenites punctata (Malyavkina) Danzé-Corsin & Laveine (1963, p. 109, Pl. 11, fig. 17) but apparently differ significantly in exine thickness, Species named after the geographic locality, Durham, Conn. Affinity: Perhaps Otozamites latior Saporta, which is the most abundant species and only genus of cycadophyte found in the black, microlaminated micritic limestone at Locality 1 (recovered by first author) and Locality 1 (Newberry, 1888, p. 90). C. durhamensis n. sp. is by far the most abundant monosulcate pollen grain at Locality 1. Localities: Shuttle Meadow Fm., 1, 12 (rare). Cycadopites cf. C. jansonii Pocock 1370 Plate 8, figures 7-9 Monosulcites "sp. K" Jansonius, 1962, p. 79, P1. 16, fig. 28-30. Cycadopites jansonii Pocock, 1970, p. 109, P1. 26, fig. 10, 15. C. paruvus Bolkhovitina 1953, p. 63, P1. 10, fig. 5, 6 (in part) ex Pocock, 1970, p. 109, P1. 26, fig. 11. Description: Pollen grains monosulcate, elliptical or fusiform. Sulcus frequently widest at extremities, narrower at distal pole. Sulcus bordered by a pair of longitudinal lips extending full length of the grain. Lips widest (2.5-7.8 um wide) at distal pole. One lip of a pair frequently wider than the other. Exine double layered, 0.5-0.7 um thick; ektexine about 0.4 um thick; endexine about 0.2 um thick, occasionally separating in part as an internal envelope. Length (6 specimens) 37-48 um; width 27-29 um. Ratio of width to length 1:1.6-1:2.7. Remarks: Our specimens agree with the type description in grain size (length 33-54 um; width 23-36 um, Pocock, 1970) and in the form of the lips. Our specimens are referred to C. jansonii with "cf." because of difference in exine thickness (the exine of C. jansonii is about 1.5 um thick). Ginkgo parva Bolkhovitina 1953, part of which species is an apparently identical form, was not validly published because of the listing of an alternative name. Pocock (1970), intending to make a new combination, Cycadopites parvus, for part of Bolkhovitina's species, actually validated the name C. parvus. Locality: Portland Fm., 5 (rare). Cycadopites reticulatus (Nilsson 1958) comb. nov. Plate 8, figures 1-3 Basionym: Entylissa reticulata Nilsson, 1958, p. 62, Pl. 5, fig. 14. Description: Pollen grains monosulcate, elliptical. Sulcus may or may not be expanded at longitudinal extremities, but never at distal pole. Frequently grains collapse, with one margin ofsulcus overlapping the other. Sulcus sometimes bordered by very narrow lips. Longitudinal ends of grain generally rounded, sometimes pointed. Exine structured with an intrareticulum that has its greatest development proximally, decreasing in size equatorially, distally, and toward the extremities, Proximai surface of grain frequently has an irregular surface, reflecting thickening of the exine in the reticulum. Lumina of reticulum 0.2-2.2 um in diameter. Exine 0.6-l.0 um thick, thicker proximally. Sculpture psilate distally, generally irregular proximally because of reticulum. Size range (6 specimens): 47-60 um long, 22-32 um wide. Ratio of width to length 1:1.7-1:2.3. Localities: Portland Fm., 5, 11, (rare). Cycadopites westfieldicus n. sp. Plate 8, figures 4-6 Holotype: Plate 8, fig. 4, size 60 um long, 29 um wide; Slide AGA1-23, coordinates 48.7 x 124.2, ref. 56.8 x 126.4. Description: Pollen grains monosulcate, elliptical. Sulcus frequently widest at extremities, narrow at distal pole. Sulcus margins occasionally overlap at distal pole. Longitudinal ends of grain rounded, rarely pointed. Lips well developed, thin, occasionally difficult to observe on corroded grains. Lips widest at distal pole (5-11 um wide), tapering toward longitudinal extremities. Lips of a pair frequently unequal in width. Exine double layered, about 1.3 um thick; ektexine 0.8-0.9 um thick; endexine 0.4-0.5 um thick. Sculpture psilate. Size range (21 specimens): 46-63 um long (median 56 um); 22-41 um wide (median 29 um). Ratio of width to length 1:1.4-1:2.4 (median 1:1.8). Remarks: This species is larger than Cycadopites jansonii Pocock, but has an exine of similar thickness. Species named after the Westfield River, site of Locality 11. Locality: Portland Fm., 11 (rare). Cycadopites sp. 1 Plate 8, figure 16 Description: Pollen grains monosulcate, elliptical. Sulcus open at longitudinal extremities of grain, margins narrower but basically straight between extremities. Lips absent. Exine (exclusive of sculpture) about 2.6 um at equatorial margin, single layered. Sculpture consists of scattered large verrucae and coni, with some bacula, primarily concentrated around equatorial sides of grain and, to a very minor extent, on proximal and distal faces. Sculptural elements 0.7-1.2 um high, 0.8-2.6 um wide. Size (2 specimens): 45-46 um long, 26-29 um wide. Ratio of width to length 1:1.6-1:1.7. Locality: upper New Haven Arkose, 8 (rare). PL ATE 5 1 2 3 4 5 6 Corollina itunensis (Pocock) comb. nov. Locality 11, 53 um. Corollina torosus (Reissinger) Klaus, emend. Locality 11, 34 um-36 um. C. torosus (Reissinger) Klaus, emend. Locality 11, 32 um-38 um. C. torosus (Reissinger> Klaus, emend. Locality 11, 31um-34 um. C. torosus (Reissinger) Klaus, emend. Locality 5, 28 um-30 um. C. torosus (Reissinger) Klaus, emend. Locality 11, 22 um-25 um. 7 8 9 10 11 12 13 14 15 16 C. torosus (Reissinger) Klaus, emend. Locality 11, 30 um. C. torosus (Reissinger) Klaus, emend. Locality 11, 37 um. C. torosus (Reissinger) Klaus, emend. Locality 11, endexinal body, 34 um (under oil). Corollina compacta Malyavkina (Neotype). Locality indicated under description, about 33 um. Corollina murphyi n. sp, (Holotype). Locality 5, 30 um. C. murphyi n. sp. Locality 8, 34 um. C. murphyi n. sp. Locality 8, 30 um. Corollina torosus (Reissinger) Klaus, emend. Locality 11; S.E.M., 28 um-33 um. Corollina murphyi n. sp. Locality 8, 32 um. C. murphyi n. sp. Locality 11; polar view with endexinal body, 42 um. PLATE 6 1 Brachyphyllum Icotii Kendall. Locality on Licking Run, near Midland, Va.; cuticle of leaf showing stomata; field about 250~Lm across. 2 B. stotii Kendall. Locality on Licking Run, near Midland, Va.; cuticle of leaf; field about 850CLn aCfOSS. 3 Corollina meyeriana (Klaus) Venkatachala & Gdcza'n. Locality 8, 26cLm. 4 C. nzeyeriana. Locality 8, 29CLm. 5· C, meyeriana. Locality 8; S.E.M., 23CLm. 6 3 10 11 12 13 14 15 C. meyeriana. Locality on Licking Run, near Midland, Va.; each grain about 54CLm. 7 C. meyeriana. Locality 1, 26~Lm. 8 C. meyeriana. Locality 1, 24~rrr. C. meyeriana. Locality 1; S.E.M., I~um. Corollz`na st'nrplex (Danz~-Corsin & Laveine) comb. nov. Locality 11, 24CLm. Corollinoid! pollen. Locality 11, 42~Lm. Corollinoid! pollen. Locality 11, 42c~m. Ar-aucariaL-ires australiJ Cookson. Locality 11, 71Clm. At·alccc?ri*cites prrtzc-tatrrs (Nilsson) comb. nov. Locality 11, S~CLm· A. punctattls (Nilsson) comb. nov. Locality 5, 48CLm· DISCUSSION Stratigraphic Correlatio~n Text-figure 5 summarites the stratigraphy of the Hartford Basin as correlated, by this study and other information, with the upper part of the Newark Basin sediments. PalynoAoras dominated by Corollina (exceeding 70% of assemblage) have been reported from the Rhaeto-Liassic and Upper Jurassic (Kendall, 1343; Harris, 1357; Couper, 1918; Lantz, 1358a,b; Pocock and Jansonius, 1961; Chaloner, 1962; Venkatachala and Gdna'n, 1964; Wall, 1965; Burger, 1965; Reyre, 1969; Volkheimer, 1371). Floras composed of more than 30% Corollina meyeriana have thus far been reported only from the basal Liassic of Larachbeg, Morvern, Scotland (Kendali, 1943). However, floras composed of a complex of Corollina species, totaling more than 300Jo, have been reported from the Rhaetian Ktissen facies, Hungary (Venkatachala and Gdcza'n, 1964). This complex contains both equaterially striate and non-striate forms, C. classoides and C. meyeria·t;ta. Palynofloras composed of 700/0 or more Corollina torosus are common in the Liassic (Harris, 1957; Couper, 1958; Wall, 1965; Volkheimer, 1371), but have also been recorded in undifferentiated Rhaeto-Liassic strata (Chaloner, 1362; Jung, 15)68). In general, non-striate to pseudostriate species are most common in the Rhaetian and early Liassic, while equatorially striate species predominate in the Liassic (Geiger and Hopping, 1968; Reyre, 1363). The palynofloras of the Hartford Basin show an upward change in Corollina from more than 30% Corollina meyeriana in the Shuttle Meadow Formation to more than 30% Corollina torostls in the Portland Formation. The age of this shift would lie somewhere between the oldest reported record of a flora dominated by Corollina torosus and the youngest record of a flora dominated by Corollina meyeriana. Although there is admittedly limited evidence, probably the shift occurred between the limits of Rhaetian and basal Liassic. Paleomagnetic data and isotopic dates (171-136 m.y.) for the Hartford Basin lava-flow units agree with a RhaetoLiassic age for this shift (Reesman et al., 1373; Reeve and Helsley, 1372). The saatigraphic position of the Triassic-Jurassic boundary in the Hartford Basin might naturally be placed in that format~on where the greatest change in Corollina species occurs, thus relegating strata containing the flora dominated by Corollina meyeriana to the Triassic, and strata dominated by Cnrollina torosus to the jurassic. It might further be argued that the Corollina-dominated K~issen facies supports a Rhaetian age for the Shuttle Meadow Formation. However, evidence of a basal Liassic Age for the formation comes from the composition of the nonCorollina portion of the assemblage. The Shuttle TEXT-FIGURE 5· Suggested time-stratigraphic correlation of Hardord Basin and upper sediments of the Newark Basin. A = Grllidlas~orites trilobat~s (Balme) Sukh Dev; D = Callialasporites cf. dampieri; H = Hirmerella muenlteri (Schenk)Jung; K = Convolzltis~ora RluRiforma (Nilsson) Schuiz; M = flora dominated by Corolli~na meyeriana (Klaus) Venkatachala & Gciczain; S = Brachyphyllzlm scottii Kendall; T = Aora dominated by Corollina torosus (Reissinger) Klaus. RedFeldius is a subholostean fish with a restricted zone in the Newark Group. Numbers shown for the Newark and Hardord basins refer to our palyniferous localities in these basins. The letters defined above indicate presence of the indicared form. M and T generally indicate dominance. Parentheses indicate forms present in variable but minor amounts. Meadow Formation contains Conuoluti~pma &IzlRiforma, which is characteristic of the European Liassic, rarely being found in the Rhaetian, and absent from the K~issen facies. Conversely, the K~ssen facies contains three species of Oval~olliJ in relatively large numbers (5-16%). This pollen genus is frequently found in the Rhaetian, but is very rare to absent in Liassic paiynofloras. In lacking Oual~olliJ, the Shuttle Meadow flora is similar to basal Liassic floras. Kendall (2949) provided only a brief description of the Larachbeg, Scotland, palyno~ora. However, she did provide a very detailed description of cuticle from Brachyphyllum scotii Kendall, associated with abundant pollen (more than 90%) supposedly derived from that plant. At a locality 2.2 km north of Midland, Virginia (Culpeper Basin), reported by Baer and Martin (1349), a similar association of CorolLina and well-preserved cuticle, including whole leaves, of B. scotii was recently discovered in a gray brown shale-siltstone sequence exposed along a brook, Licking Run. Many palyniferous layers also occur in the Licking Run section. Fish-bearing black calcareous shale outcrops suatigraphically about 2.2 m below the megafossil plant bed~ The fish fauna from this locality is remarkably similar to the one from the Shuttle Meadow Formation at Locality 1 (Comet, et al., 137~; Schaeffer, 1967; P. E. Olsen, pers. comm.). A more detailed study of the Midland flora is in progress. The Midland flora is essentially identical to that from the Shuttle Meadow Formation, including the presence of Conuolzltispora RI~R~forma. The abundant pollen, including tetrads and pollen masses, associated with the B. scotii cuticle at Midland, is identical with Corollina meyeriana (Klaus) Venkatachala and Gdcza'n. Several distinct species of Dictyot:·'!etes (a genus characteristic of the Jurassic) are also present. One of these species (Dictyotriletes sp. 1) is present at Locality 8 in the upper New Haven Arkose of Massachusetts: (Locality 8 is considered contemporaneous with part of the Shuttle Meadow Fm.; Text-fig. 3). Moderately corri~ded but identifiable cuticle, including whole leaves, of B. scotii has also been recovered from Shuctle Meadow Locality 1 of the Hartford Basin and from lake beds above the lowest lava-flow unit (First Watchung Mt.), just east of Round Top Mountain, near Oldwick, New Jersey (Locality 4, Newark Basin, Text-fig. 5). The Shuttle Meadow, Midland, and Larachbeg palynofloras appear to have much in common. The identification of B. scotii merely indicates that a basal Liassic age is possible. Only one age horizon is known for this conifer – thus it is of little timestratigraphic value, especially if it were the only source for C. meyeriana, which is doubtful. However, the presence of B. scotii, C. klukiforma, Dictyotriletes sp. 1, the dominance of C. meyeriana, and the absence of any palynomorph known to be characteristic of only the Triassic, favors an early Liassic age for the Shuttle Meadow and Midland floras. Todites pinceps (Presl) Gothan was discovered by the first author in the youngest sediments of the Culpeper Basin, near Leesburg, Va. This fern has been found mainly in late Rhaetian to Middle Jurassic age strata (Harris, 1948, p. 182, 183j. ClathropteriJ meniscoides Brongniart has been found at Localities 2 and 8 in the Hardord Basin, and Locality 4 in the Newark Basin. This fern may range from Carnian through the Liassic, butis particularly common in the Rhaetian and lower Liassic (Harris, 1931)· Both of these ferns were found in association with a flora dominated by C. meyeriana, and this strongly indicates a RhaetoLiassic age for this type of association. In order to examine palynofloras that are slightly older than those of the Shuttle Meadow Fm. of Gonnecticut, one must ref~r to strata in the Newark Basin of New Jersey, because palynomorphs have not been found in the older Talcott Fm. of Connecticut, nor in strata of the New Haven Arkose known to be older than the Talcott (Text-fig. ~). Correlation of footprint and fish faunas (P. E. Olsen, pers. comm.), and of palynofforules and lava flows for the Hartford and Newark basins (Text-fig. 5), indicates that some strata in New Jersey containing a typical Shuttle Meadow palynoflora may be older than the Shuttle Meadow Formation of Connecticut. The seventeen fish localities so far investigated by Olsen from sediments between the First (lowest) and Third (highest) Watchung lava-flow units lack the subhoiostean, Redfi~ldizls spp., which is frequently found in younger strata. Therefore it is apparent that the flora dominated by C. meyeriana (occurring with B. scotii) has considerable snatigtaphic range, calculated in the Newark Basin to be greater than 600 m (not including thickness oflava-flow units). The presence of Upper Triassic reptiles, such as StegomuJ, Hypsognathus, and the phytosaur, cf. Rutiodon (P. E. Olsen and J. H. Osuom, pers. comm.), in the middle part of the New Haven Arkose (Text-tig. 5) strongly favors placing the TriassicJurassic boundary just above the Talcott Formation (which at present would not conflict with paleomagnetic data). The boundary might correlate with the erosional unconformity in the Hartford Basin. Recently a well-preserved Carno-Norian palynoflorule (Locality 1, Newark Basin, Text-fig. 5) was obtained from gray shales of the lower Brunswick Formation at Milford, New Jersey (gray members L-M of McLaughlin, 1946). The florule is dominated by bisaccates: AliJporites, Kla~sipolleniteJ, Pityosporites, Protodiploxypinzls, Platysacczls, Triadispora, Chordasporites?, and PodosporiteJ? There are also moderate percentages of Camerosporites verrucosus M~dler, Patinasporites densz~s Leschik, VaLlasporites ignacii Leschik, and tetrads of spherical psilate grains ("Genus A") only before described by Dunay (1972) from the Upper Triassic Dodtum Group of northwest Texas. The stratigraphic position of this middle Keuper florule from New Jersey is about 920 m above "gray member B" of McLaughlin (1344), which is the top of the underlying Lockatong Formation. However, the relationship of the florule to the overlying First Watchung Mountain basalt is less certain, as most of the Brunswick Formation, including the lava flows, is not preserved around Milford, N.J., and very few "unfaulted" sections of the entire lower Brunswick have been measured. Kummel (1836, p. 58) measured one such section "between the mouth of Lawrence Brook, east of New Brunswidc, and the base of the first trap ridge back of Bound Brook" as being 3050 m thid(. Because of the relative proximity of this measured section to Milford, N.J. (in relationship to the rest of the basin), this figure can be safely used to calculate the approximate distance of the~Milford florule beneath the First Watchung Mountain lava-flow unit, a distance of critical importance in the following argument (6. Text-fig. 5). Van Houten (1363) estimated the rate of sedimentation fbr the Brunswick Formation at 30~ +mml1000 yr. The Milford florule lies approximately 2100 m below the First Watchung Mountain basalt (= 3050 m-320 m). The Milford Carno-Norian florule would then be separated in time from the oldest known occurrence of a florule dominated by Corollilza meyerzana (occurring with Brachyphyd Illm SGOtii) in the Newark Basin by at least 6.3 m.y. This amount of time would accommodate much of the Norian and Rhaetian stages and agrees well with the paleomagnetic normal polarity of the lava flows, which can readily be explained if most of the Rhaetian occurred before extrusion of the lavaflow units (Creer, 1371; Reeve and Helsiey, 1972). For comparison, the maximum thickness of Upper Triassic strata in England and Germany appears to be 1400 m, but it is usually less than 500 m (Geiger and Hopping, 1968; Wills, 1970). A probable upper Norian palynoflorule (samples provided by Donald Baird, Princeton Univ.; coll. by N. K. Resch) occurs in a gray siltstone with reptilean footprints, exposed in the construction of U.S. Rt. 280 behind the old Second Precinct Police Station at Newark, N.J. (Newark Basin Locality 11, Textfig. 5). The florule is significant in that it possesses equatorially striate grains of Corollina torosus (Reissinger) Klaus, as well as C. meyeriana (Klaus) Venkatachala 8r Gdcza'n, Patinasporites densuJ Leschik, possibly Enzonalasporites vigens Leschik and TriadiJpora sp. C. torosus, generally not reported below the Rhaetian, and Triadispora sp., rarely reported above the middle Norian, are seldom found in the same assemblage. In addition the presence of P. densus suggests an age no younger than Norian because, while this species occurs as high as the lower Rhaetian, it is rare above the Norian. Corollina spp. represent about ~O% of the assemblage. Bisaccates of the Pityosporites and Alisporites types are common, and Converrucosisporites cameronii (de Jersey) Playford 8r Dettmann and Granulatisporites infirmus (Balme) comb. nov. are much more common than in older floruies of the Newark Group. This palynoflorule is calculated to be about 1100 m below the First Watchung Mountain basalt. The rather low stratigraphic position of middle and upper Keuper florules in the Brunswick Formation supports the paleomagnetic evidence that at least part of the Brunswick below the First Watchung Mountain basalt is Rhaetian, but does not rule out the possibility that strata containing the generally Liassic C. ~neyeriana-B. Jcotii flora may extend downward into the upper Rhaetian. For this reason this floral assemblage is here designated Rhaeto-Liassic wherever it occurs. The Portland Formation florules of the Hartford Basin are generally dominated by C. toroszls (Table 1) and its presumed megafossil sources, Hirmerella sp. (Locality 7, Comet et al., 1373) and Hirmerella nzzlenJteri (Schenk) Jung 1368 (Localities 10, 11, and 13). Lower Portland flotules (Localities S, 6, 7, and 13) appear to vary laterally and to fluctuate vertically in Corollirza spp. percentages, with C. meyeraana sometimes comprising much more than 2~0, but rarely being dominant. poor preservation of most of these fforules makes specific identification difficult, but cuticle fragments show that the ratio of Brachy~hylltlm scotaz to Hirmerella spp. varies in proportion to the apparent ratio of C. meyeriana to C. toroJus. The minor constituents of the Portland Formation assemblage include several good Liassic or Jurassic indicators, such as Carollina itzlnensir, Callialasporites 6. C. dampieri, CaLLzalasporites trilobatt~s, Araz~cariacites fissz~s, Cycadopites reticulatzls, Perotrilete~ 6. P. pselldoreticlllatzls, LRptole~iditeJ cf. L. major, and Gleicheniidites cf. G. ·nilssonii. The overall composition of the flora compares well with that of the Liassic (Balme, 1857; Couper, 1358; Nilsson, 1958; Wall, 1365; Schulz, 1967; Reiser and 'Williams, 1363; Pocock, 1370; Volkheimer, 1971; Volkheimer, 1972; Antonescu, 1373), particularly in the dominance of Corollina torosus and the presence of Corollina itzlnensiJ, C. meyeriana, Araucariacites australiJ, A,Fssus, Cycadopites deterizlJ, C. reticsclattcJ,'C. janJonii, AliJporites thomasii, Pityopollenites pallidus, Platysaccus 6. P. lopJiensis, PerinopoLleszites elatoides, Callialasporites spp., GranulatiJporiteJ infirmus, ConveP·PucosiJporiteJ cameronii, Todisporites rotundiformis, Gleicheniidites cf. G. rrzilssonii, LRptolepidktes sp., and HarrisiJpo~a sp. Even though many of these palynomorphs are long ranging and not good index fossils, together they make up a flora unlike any flora reported from the Triassic. In contrast to those of the Upper Triassic, palynofloral assemblages of the Liassic are relatively uniform, both geographically and chronologically. Although new species appear throughout the Liassic, floral change is gradual, with a. number of plant types forming dominant associations over wide geographic areas. Minor floral constituents seem to be the most variable. Several good index spores and pollen have been recognized, such as certain species of Dictyotriletes, ContzgniJporiteJ, and Callialasporites. When present in significant amounts (generally greater than 1%), these forms can be used to distinguish the upper part of the Liassic from the lower and middle parts. Antonescu (1973) reported the presence in very small amounts of several characteristic upper Liassic palynomorphs in the middle Liassic of Rumania: Callialasporites and Leptolepidites. The rare occurrence at our Locality 11 of CallialaJporiteJ 6. C. dampieri, CallialasporiteJ trilobatzls, Leptolepidites PL ATE 7 1 2 3 4 5 6 7 Carmerosporites reductiverrucatus n. sp. (Holotype). Locality II, fj4Clm. C. reductiverrucatus n. sp. Locality 11; composite photograph of two focal levels, 46Ccm-561~m. C. reductiverrucatus n. sp. Locality 1 1, 5 ~um. C. reductiverrucatus n. sp. Locality 11, 58tcm. C. reductiverrucatus n. sp. Locality 11, uiradiate fold present; distal part of grain missing, 33CLal· Araucariacites fissus Reiser & Williams. Locality 11, 30CLm· A, fissus. Locality 11, 76tGm. PLATE 8 1 2 Cycadopites reticukr~us (Nilsson~ comb. nov. Locality C. retict~latzls~ (Nilsson) comb. nov. Locality 11, 48~m (under oil). C. retictilatzis (Nilsson) comb. nov. Locality 5, 32CLm wide (under oil). 4 C~L-crdopites z~~e.rtfEelclic/rs n. sp. (Holotype). Locality 11, GOE~Lm. 5. u~esif~elrlicus n. sp. Localit)r 11, ~O~m (under oil). 6 C. rceJrf;rldicNs n. sp. Localit)r 1 1, (,0 CLm(under oil). 7 Cycadopites 6. C. janJonii Pocock. Locality ~, 36~m. 8 C~~cadopitss 6. C.iansonii. Localir)· ~, 35~Lm. 9 Cycado~iles 6. C.iansonii. Locality ~, 46CLm. 10 C3,cadopites deterius (Balme) Pocuck. Locality 5, 11 12 13 14 15 16 17 18 19 20 21 48~m. C. deteri~s. Locality 11, 4 I~m. C. dett·ri~s. Locality ~, Tl~m~ Cycadopites durhamensis n. sp. (Holotype). Locality 1, 26~Lm. C. dllrhamenJis n. sp. Locality 1, 28~m. C. dtirhamensis n. sp. Locality 1, 29Ccm. C~lcadopites sp. 1. Locality 8, 4SjCLm· Cycado~ites andrewsii n. sp. Locality 8, S.E.M.; 34CLm. C. annrewsii n. sp..locality 8, 37ctm. C. andrewsii n. sp. Locality 8, 341~Lm. C. andrewsii n. sp. (Holotype). Locality 8, ~7cLm. C. andrewsii n. sp. Locality 8, 42CLm. cf. L. niirlor, and C~rOllind zt~nensis, generally not reported below the upper Liassic, suggests that the middle part of the Portland Formation is perhaps Pliensbachian or youn~er in aae. Dictyotriletes spp. (Pi. 1, fig. 4-6) and Convolz~tispora ~luRiforma are present in the Hartford and Culpeper basins in strata presumed to be older than the Portland Formation, Such an occurrence strongly favors an early to middle Liassic age for much of the Portland Formation (Text-fig. 5). The upper part of the Portland is as yet unsampled, but based on stratigraphic thickness, this part of the formation could well be upper Liassic. Galton (1971) compared the prosauropods Anchisaurus and AmmosaurEls, and the primitive crocodile StegomoJuchzcs of the upper Portland Formation with closely related reptiles of the Navajo Sandstone of northeastern Arizona, suggesting that these formations were closely contemporaneous. Galton, on the basis of previous authors, presumed the Portland Formation to be Upper Triassic, and therefore suggested that the Navajo Sandstone might also be Triassic. However, most previous investigators have assigned a probable Early to Middle Jurassic age to the Navajo (Galton, 1971, p. 791, Text-fig. 13). If the sedimentation rate mentioned above for the lower Brunswick Formation can be applied to the 2000+m of Portland Formation, a minimum 6.25 m.y. duration would be indicated. This would be less than the extent of time indicated by palynological evidence, suggesting that the average rate of sedimentation for the Portland Formation was about half that given by Van Houten (1363) for the Brunswick Formation. Comparison of similar types of sediment were made for sections of the lower Brunswick Formation along the Delaware River, N.J., and the Portland Formation along the Connecticut and Wesfield rivers, Mass. Pdleoecology Several types of environment for source plants have been proposed for assemblages dominated by Corollina pollen, particularly C. torosus: (1) coastal swamp habitat to account for Corollina pollen in offshore marine deposits and poor Corollina representation in back delta environments (Hughes and Moody-Smart, 1367); (2) coastal or lagoonal environment under dry climatic conditions to account for sediments rich in Corollina, laterally associated with strata containing marine microplankton assemblages in one direction, laterally grading to strata rich in Monosulcites-type pollen in the other direction (Pocock and Jansonius, 1961; Venkatachala and Gocza'n, 1964); (3) upland sandy slope environment with source plant tolerance of drought conditions (Vakhrameev, 1970). Occasional association of megafossils of Corollina- producers with Equisetites, or of Corollina pollen with Equisetites and articulate spores, is not significant enough to suggest any particular environmental relationship (Batten, 1973). Similar associations occur in the Hartford Basin, and we suggest that Corollina pollen, because of relatively high density, probably reached some of the same sites of deposition largely through river drainage systems, as did Equisetites. A similar course of transportation could easily account for the dominance of small, broken fragments of Hirmerella and Brachyphyllum at many megafossil localities. The sediments of the central area of the basin are predominantly fine grained. Many of these sediments were probably deposited in variable, shallow to deepwater lakes, as evidenced by turbidites, slumps, and dark, gray black shales and siltstones within reddish brown shale and siltstone sequences (Sanders, 1368), locally containing horizons with reptilean footprints. Since no known evidence of paleosols or root penetration zones exists in these finer grained deposits, and a large footprint excavation (Rocky Hill Dinosaur State Park, Conn.) indicates extensive mudflat conditions, it is safe to assume that Corollina producers did not frequently grow there, Predominantly coarse-grained sediments in the marginal parts of the basin support fluvial origin (Krynine, 1950). It is possible that Corollina producers grew on these fans and deltas, thus supporting, in part, Vakhrameev's sandy slope environment. It is interesting that this fluvial environment also suggests an association with small basin swamps, lakeside habitats, and lagoons. However, it is difficult to envision a relatively pure population of Corollina producers restricted to the basin. Corollina accounts for more than 30% of the palynoflora in sediments from large areas (in absolute terms, non-Corollina palynomorphs are usually less than 350 per gram of sediment), and there do not seem to be sufficient areas of potential Corollina- producer environments in the basin proper to account for the masses of pollen found. The absence of true coals and scarcity of coniferous megafossils in the basin argue against extensive swamps such as those dominated by Taxodium or Rhizophora today. The dominance of Corollina producers in the younger sediments of the Culpeper, Newark, and Hartford basins suggests that a large geographic area was covered by this assemblage. Others have suggested that dominance of Corollina pollen probably indicates close proximity of marine or coastal environments (Pocock and Jansonius, 1961; Wall, 1965; Batten, 1373). Some previous authors (Pocock and Jansonius, 1361; Venkatachaia and Gdcza'n, 1964; Vakhrameev, 1970) have claimed that Corollina pollenproducers occurred in regions with generally arid to semiarid climatic conditions, but have not given convincing evidence to support this opinion. However, the source plants show no convincing xeromorphic adaptations -- microphyIly by itself is not necessarily such an adaptation. Furthermore, Krynine (2850) proposed a warm, humid savanna climate, witha heavy but seasonally distributed rainfall and a dry season of two or three months for the Hartford Basin. Recent recognition of microlaminated (varved) micritic lake beds in the Shuttle Meadow Fm., probably produced in a meromictic lake under the influence of a seasonal climate (Comet, et al., 1373), might support Krynine's wet-dry climatic interpretation. It is highly improbable that the Hartford Basin flora, which includes many kinds of cryptogams (based on spore diversity), including large-leaf forms of Clathropteris, and has few known representatives with well-developed xeromorphic cuticular adaptations, could tolerate long periods of aridity. Acknowledgments The authors acknowledge the support of The National Science Foundation (Earth Sciences Section, grant number GA-36870) for all phases of this project. We also acknowledge the generous advice on field problems and stratigraphic correlation, as well as in location of palyniferous zones, of P. E. Olsen, Yale University. Professor Donald Baird, Princeton University, has provided us with the sample designated No. 11, from the Newark Basin. Professor F. G. Van Houten, Princeton University, read portions of the manuscript and suggested useful improvements. R~erences ANTONESCU, E., 1~73, QNElyNes donne'es sur ki PL?1~'120laRje du Lias soils faCi~J. de G res te n de R(,s( malzie. Palynology of Mesophyte, 3rd Internat. Palyn. Conf., Proc., p. ~ ~-5 7. BAER, F. M. and MARTIN, W. H., 15)48, Some neu' finds offossil Ranoids in the I/irginia Tviassic: Science, v. 110, n. 2863, p. 684-686. BAIN, G.'W., 1~41, The Holyoke Rtrnge and Connecticut I/L/lley structure: Amer. Jour. Sci~, v. 2~9, p. 261-27r. BALME, B. E., 13~7, Spores and pollen grains from the MeJozoic of western Alrlstral;a: C.S.I.R.O. (Australia), Coal Res. Sect., T.C., v. 25, 48 p. BALME, B. E. and HENNELLY, J. P. F., 1~56a, Monolete, monocolpate, and alete sporomorphs front A//stralian PL·rmian sediments= Austral. Jour. Bet., v. 4, p. 5j4-67. 13SGb, Trilete sporomorphs from A//stralian PL·rmian sediments= Austral. Jour. Bet., v. 4, p~ 240-260. BARNARD, P. D.'W., 1~68, A new species ofMasculostrobus Seward Producing Classopollis pollen from the JNrassic of Iran= Jour. Linn. Sec. (Bet.), v. 61, n. 384, p. 167176. BATTEN, D. J., 1~68, Probable dispersed spores of Cretaceous Equisetites: Palaeontology, v. 2, n. 4, p. 633-642. ---- 1973, Prllynology of Early Cretaceozls soil beds and associated strata= Palaeontology, v. 16, n. 2, p. 3~~424. BOLKHOVITINA, N. A., 23~3, spore-pollen character of Cretaceous deposits of the central provinces of the USSR= Trudy inst. Geol. Sci., Acad. Sci. USSR, 145, Geol. Set. n. 61, 184 p. (in Russian). ---- 1356, Atlas of spores and potlen from JNrasJic and Lower Cretaceozls deposits of the ~ilyt/isk basin. Inst. Geol. Sci., Acad. Sci. USSR, Trans., v. 2, 186 p. (in Russian). BROPHY, G. P., FOOSE, R. M., SHAW, F. C. et al., 1367, Tt·iassic geologicfeatz~res in the Connecticut ~C/alley near Amherst, Massachusetts, Trip D. Gir/idebook for Feld trips in the Connecticut F/alley of Massachusetts, New England Intercoll. Geol. Conf., S3th Ann. Meet., Oct. 1~-1~, 1367, p. 61-72. BURGER, D., 196~, Si,me new species of Classopollis from the JNrasJic of the NetherlandJ: Leid. Geol, Meded., v. 3~, p. 63-63. CHALONER,`W. G., 1C)62, Rha~to-Liassic plants IC?Om the HL' nFeld borehole: Geol. Survey Gr. Brit., 'Bull., v. 19, p. 1G·28. CHANG, L. C., 1363, Nomenclature and distribution of several Mesozoic gymnospermous ~ollen: Acta Paieont Sinica, v. 11, n. 3, p. 43~-~42 (in Chinese and Russian). CHAPMAN, R. W., 1~65, Stratkgraphy and petrology of the Hcrmpden basalt in central Cnnnecticut: State Geol. Nat. Hist. Survey Conn., Rept. Invest. n. 3, 38 p. COLTON, R. B. and HARTSHORN, j. 1-1., 1966, BedrocR Keolo~y oJ~ the 1E~est Sprin~eFelri eNcrdrrrnRle: U.S. Geol. Survey, Map GQ-537. COOKSON, I. C., 1347, Plant microfossils from the lignites of Kcrg~elen Archi~elago: B.A.N.Z. Antarctic Res. Exped., 1323-135 2, Rept. Ser. A, v. 2, p. 127-142. CORNET, B., TRAVERSE, A., and MCDONALD, N. G., 197~, Fossil spores, pollen, and fishes from Connecticut indicate Early JNrctssic age for part ofthe NezuarR Gvollpr Science, v. 182, n. 4118, p. 1243-1246. COUPER, R. A., 195~, I/pper iMesozoic and Cainozoic spores and ~Clollen grains from Neu, Zealand= N. Zea. Geol. Survey, Paleont Bull, v. 22, p. 1-77. ---- 1958, British Mesozoic microspores and ~ollen grains, A systematic and stratigraphic study: Palaeontographica, B, v. 10~, p. 75-179. CREER, K. M., 1371, Mesozoic palaeomagnetic reversal colu7mn: Nature, v, 233, p. 54~-546. DANZE-CORSIN, P. and LAVEINE, J.-P., 1363, Microflore: in Flore infraliasique du Boulonnais (Macro- et Microflore), P. Briche, P. Danze'-Corsin. and J.-P, Laveine, Sec. Ge'oi. Nord, Me'm., vl~, p. 57-155. DAUGHERTY, L. H., 1341, The Upper Triasric JPora of Arizona: Carnegie Inst. Wash. Publ., v. 526, 108 p. DELCOURT, A. F. and SPRUMONT, G., 1955, Les sporeJ er les grains de Pollen du Wealdien du Hainazlt= Sec. ]Belg. GCol., Mem., n.s., v. 5, p. 1-73. DORING, H., 1361, Planktonartige Fossilien des JllralKr~ide-GrePzzbereichs der Bohrungen Werle (Mrcklenburg): Geologie, v. 10, Beih.,~v. 32, p. 11~-122. DUNAY, R E., 1972, The palyrtology of the Triassic DocRum Groll~ of Texas, and its application to stratigraphz'e Problems of the DocRzlm Groflp= Ph.D. diss., Pa. State Univ., ~70 p. ERDTMAN, G., 1947, SNRgestions for the classification of fossil and recent pollen grains and spores= Svenska Bet. Tidskr., v. 41, p. 104-114. FISHER, M. J., 1972, Rhaeto-Liassic ~alynomorphJ from the Bcrrnstane R~tilway clltting, NottinghamJhire: Mercian Geologist, v. 4, n. 2, p. 101-106. GALTON, P. M., 1371, The P1·osazcropod dinosaur Ammosaurus, the crocodile Protosuchus, and their bearing on the age of the Navaio Sandstone of northeastern Arizona= Jour. Paleont., v. 45, p. 781-735. GAMERRO, J. C., 136fi, Morfologia det polen de Apteroladus lanceolatus Archang. (Co·niferae) de la Formacio'n Brtquerod, Provincia de Santa Cruz= Ameghiniana, v. 4, n. 4, p. 133-136. GEIGER, M. E. and HOPPING, C. A. 1~68, TrMssic stratigraphy of the Jozlthern North Sea basin: Royal Sec. London, Philos. Trans. Ser. B, v. 254, p, 1-~6. HARRIS, T. M., 2~~1, The fossil flora of Scoresby So21nJ, East Greenland= Meddel. Gz~nland, v. 85, p. 1-104. ---- 1348, Notes on the Jtlrassic 1Qora of Yorkshire. Ann. Mag. Nat. Hist., Set. 12, v. 1, p. 181-213. ---- 15)57, A Liasso-Rhaetic fEora in South Wales= Royal Sec. London, Proc., v. 147, p. 28~-~08. HITCHCOCK, E., JR., Is~~, Descri~tiott of a new species of Ciathropteris, discovered in the Connecticut I/rrlley Sandstone: Amer. jour. Sci., Ser. 2, v. 20, p. 22-25. HOFFMEISTER, W. S~, STAPLIN, F. L., and MALLOY, R. E., 195~, Mississippran plant spores from the Na7dinJb~rg Formation of Illinois and Kentzicky: Jour. Paleont., v. 29, p. 372-389. HUGHES, N. F. and MOODY-STUART, J. C., 1367, Pcr(yno- loKiiirl J~c-ies ~~lrl C~YTel~ti(,ll ill tht;. EI1RliSh I~ealden. Rev. Palaeobot. Palyn., v. 1, p. 2~~-268. IBRAHIM, A. C., 1~~2, See Potonie~, Ibrahim & Loose, 1332. 1933, Sporenformen des Argirhorizonts des Ruhr-Reviers= diss., Tech. Hochsch., Berlifl, 48 p. JANSONIUS, J., 1~62, Pal3inology of Permian and Triassic sediments, Peace River area, w~stern CL,nada: Palaeontographica, B, v. 110, p. 35-38· JERSEY, N. J. DE, 1~62, Tviassic spores and Pollen Rrains from che I~swich coalfield: Geol. Survey Queensland Publ., v. 307, p. 1-18. ---·---- 1971, Eclrly Jrrrassic miospores from the Hclidon Sandstone: Gcol. Survey Queensland Publ. 351, Paiaeont Pap. 25, p. 1-49. JERSEY, N. J. DE and PATEN, R. J., 1364, J~raJsic sihoreJ and Pollen grains from the Stlrat Basin= Geol. Survey Queensland, Publ. 322, p. 1-18. JUNG, 73~'. 'W., 1~6s, Hirmerella miinsteri (Schenk) JNnR nov. comb., eine bedeutJame Kr,nifere des Mesozoikums: Palaeontographica, B, v. 122, n. 1-3, p. 5~-33. KENDALL, M. WT,, 1343, 012 a neu~ cona~fer from the Scottish Lias= Ann. Mag. ~dar. Hist., Ser. 22, v. 2, p. 2 33-3 07. KLAUS, Vlr., 1360, Sporen der karnischen Sr~fe dsr os~alpinen Trias: Jahrb. Geol. Bundesanstalt (7JCrien) Sonderbd., v. 5, p. 107-18~. KRAUSEL, R. and LESCHIK, G., 13~6 (imprint date 1955), Die Kc·~perflora uon Nc~ueu?elt bei BLIJel. II, Die Iso~nd MikroJporen: Schwei~. Pal~ont. Abh., v. 72, p. 1-70. KRYNINE, P. D., 1~~0, Petrology, stratigraphy, and origin of the Triassic sedimentary rocRJ of Connecticzlt= State Geol. Nat. Hisr. Survey Conn., Bull., v. 7~, 247 p. KUMMEL, H. B., 1836, The Nrwark syJtem, report of progresr. !I= Geol. Survey N. Jersey, Ann. Rept. State Geol., p. 25-88. LaNTz, J., 19SSa, I~tzlde des spores et Pollens d'un ~chantillon P~rbeckien de I'lle d'Ole'ron~ Rev. Micropal~ont., v. 1, p. 3~_~7. 1958b, ~tude palynologique de q~etques gchantillons MPsozoiqueJ due Dorset (Grande-Bretagne). Rev. Inst. Fr. Petrole Ann. Combust. Liquides, r~. 13, p. 3 1 7-34 ~. LEHMANN, E. P., 1~5~, The bedrock geology ofthe Middletown q~adrangle u·,ith map: State Geol. Nat. Hist. Survey Conn., Quadrangle Report 8, p. 2-40. LESCHIK, G., 1356 (imprint date, 18~5) in Kr~usel afld Leschik, 1956. MADLER, K., 1864, BemerRensu·erte Sporenformen aus dem Ketcper zlnd t~nteren Lras: Forrschr. Geol. Rheiflland und 'Wesdalen, v. 22, p. 163-200. MALYAVKINA, V. S., 1~49, INdex of spores and pollen, JNrasJic- Cretaceo~s= Trudy VNIGRI, n.s., n. 3~, Gostoptekhizdat, Moscow, 1~13 p. (in Russian). --~--- 1953, Spore and polletz complexes of the Upper Triassic csnd Lower and r~jddle /~rassic from the Eastern and Western Pre- Urals= Trudy VNIGRI, n.s., n. 7~, 5)3-147 (in Russian). MCLAUGHLIN, D. B., 1344, ?er-iassic JtratiRraphy in the Point PleaJant Distvict, Pennsylvania: Pa. Acad. Sci., Proc., v. 28, p. 62-69. ~ 15)46, The Triassic rocks of the HNnterdon Plateair, NewJersey.· Pa. Acad. Sci., Proc., v. 20, p. 88-98. MINER, E. L., I~jS, Iv,~lrt,l,c/trrnjCLrI t·xamination of Crctaieo~s L~nd rrcirtiav~l ionls.· Amer. Midi. Nat., v. 1G, p. 58r-62~. NAUMOVA, S. N., 1~~~, S~ort·s rrnJ Pollen of`the coals of the USSR: 17th Internat. Geol. Cong., 1337, Trans., v. 1, p. 3r~-364. NEWBERRY, J. S., 1888, F,,ssil _Fshes and fossil plants of trG~~ Tviassic rocks of N~u~ JerJ-ey and the Connecttctlt I~rlley= U.S. Geoi. Survey, Mono.. v. 14, p. 1-35. NILSSON, T., 1~~8, ~iber das Vorkommen eines mesezo ische n Saprope Iges re i ns in Schonen: Publ. Inst. Miner. Paleont Quat. Geol. Lund, v. 53, 112 p. PETTITT, J. M. and CHALONER, W. G., 1364, The ultraJtructure of the Mesozoic Pollen Classopoliis: Pollen et Spores, v. 6, n. 2, p. 611-620. PFLUG, H., 19~3, ZNT ErztJtehung und EntwicRlung des angiospermiden P~tlens in der ErdgeJchichte= Palaeontographica, B, v. 35, p. b(r171. PLAYFORD, G. and DETTMANN, M. E., 1965, RhastoLiaJsic plant microfoJsilJ from the Leigh CreeR coal meastlres, South ANstralia: Senckenbergiana Lethaea, v. 46, p. 127-181. Pococrc, S. A. J., 1862, Microf~oral analysis and age determination of strata at the Jl/rassic-Cretaceozls bozlndary in the zoeJtern Ccrnada plains= Pa~aeontographica, B, v. 111, p. 1-31i. -------- 1970, Pr,lynotogy of the JNrassic sediments of western Crrnada: Palaeontographica, B, v. 1~0, p. 12-136. PococK, S. A. J. and JANSONIUS, j., 1361, The pollen genus Classopoliis PfE~g 1953: Micropaleontolgy, v. 7, p. 4~8-449. POTONIE, R., 1356, Synopsis der Grrtttlngen der Sporae dispersae, Teil i: geol. Jahrb. Beih., v. 23, 103 p. -------- 1958, SynopJis der Gattzlngen der Sporae dispersae, Teil II= ~eol. Jahrb. Beih., v. 51, 114 p. POTONIE, R., IBRAHIM, A., and Loose, F., 1932, Sporenformen aus detz Flb'zen ~gir ~nd BiJnzasck des R zl hrge bietes: Neues ]ahrb. Miner. Geol. Pal~ont. Beilageband, B, v. 67, p. 4~8-454. POTONIE, R. and KLAUS, W7., 1954, EiniRe S~orenRatGzlnRen des alpinen ScrlzRebirgeJ= Geol. Jahrb., v. 68, p. 517-546. POTONIE, R. and KREMP, G., 1954, Die GtrttunRen der palliozoischen Sporae dispersae zsnd ihre Strcrtigraphie= Geol. Jahrb., v. 69, p. 111-184. -------- 1355, Die Sporae dispersae des Ruhrkarbons. I. Palaeontographica, B, v. 98, 136 p. REESIDE, J. B., JR., APPLIN, P. L., COLBERT, E. H., et al., 29r7, Correkrtion of the Trias~ic Jormations of` North America exclusive of Ctrncrda= Geol. Sec. Amer., Bull., v. 68, p. 1451-1514. REESMAN, R. H., FILBERT, C. R. and KRUEGER, H. VC/., 1973, Potassium-argon dating of the Upper Triassic lavas of the Cr,nnecticut I/Lllley, New England= Geol. Sec. Amer., Abstr., v. ~, n. 2, p. 211. REEVE, S. C. and HELSLEY, C. E., 1972, Magnetic reversal sequence in the upper Portion of the Chinle Formation, Montoya, New Mexico: Geol. Sec. Amer., Bull. 8~, p. 373~-~812. REISER, R. F. and 7JCIILLIAMs, A. J., 1969, Pcrlynology of the Lou?er J~rassic sediments of the northern S,rrat Bdsin, eNeensland: Geol. Survey Queensland, v. 333, REISSINGER, A., 19~0, Dje "Pollenanalyss," ausgedehnt a~f II.· Palat~ontogr_aphic3, B, v. ')0, p. ~~-1~G. REYRE, Y., 19~j~~. S~Ereo.rCall o~~rer.l.r,lj~n.i oN the POIICII Ren~s Classopollis P~uK IY53. Palaeontology, v. 19, n· 2, p. 305-322. ROss, N. E., 194(), 011 a Cletaceol/s pollen and spore hecrrillK clap deposit of Sian;a.· Geol. Inst. Univ. Upsala, Bull., v. ~4, p. 25-43. SAH, S. C. D. and JAIN, K. P.~ 1365, JNrassic spores and pollen grains from the Rr~rlmahal Hills, Bjhar, Illdia, With a di.rcussion on the aRe of the R~~mahal IlltertrapPt·an beds: Palaeobotanist, v. 13, n. 3, p. 264-280. SAMOILOVICH, S. R., 1953~ P,,llen and spores from Pcrmian depositJ of the Cherdy~s~ and AktyubinsR pre-LlralJ: Trudy, VNIGRI, n.s., n. 75, p. 5-57 (in Russian). SAMOILOVICH, S. R., et al., 1961, Pollen and spores of western Siberia, Jurassic-Paleocene, Trudy, VNIGRI, n. 177, GGOp. (in Russian). SANDERS, J. E., 1968, StrcrtiRraphy and primar~ sedimentary srructllves of fine-9rained, u'ell-bedded strata, inferred lake de~ositJ, Upper TrMsJic, central and southern Cnnnecricut: Geol. Sec. Amer., Spec. Pap. 106, p. 2GT-30~. 1970, S,rati~raphy and strrrcture of the Triassic strata cifthe Gaillard Graben, south-central Connecticuti State Geol. Nar. Hist. Survey Conn., Guidebook n. 3, SANDERS, J. E., ~UIDOTTI, C. V., and WILDE, P., 1363, Foxorr F~lult and G(lillard GI·aben in the TriasJic of southern Connecticut: State Geol. Nat. Hist. Survey Conn., Rept. Invest. n. 2, p. 1-16. SCHAEFFER, B., 1967, Lrlte TriassicfiJhesfronz the u'eJtern United States. Amer. Mus. Nat. I-list., Bull. 135, p. 287-542. SCHEURING, B. W., 1970, P~rlynoloRiJche und palyno. Jtrati~rcrphische (/tltersz(chllngen des Keupers im chentunnel (Sr,lothurner _I//ral: Schweiz. Palaont. Abh., v. 88, p. 1-119. SCHNABEL, R. M., 1960, Bcdroc~ geology of the AI,on e~adran~le, Connecticut. U.S. Geol. Survey, Map GQ-154. SCHULZ, E., 1967, Sporenprrl;iontoloRische U,lterszlchungen rlitoliaJ·sischev SL-hichten im Zentra(teil des Gcrmanischen Beckens: Pal~ont. Abh. (B), v. 2, n. ~, p. 547-633. SINGH, C., 1864, rl4jrvof~ora of the Lllu~er Cretaceoz/s M~lnnville group, east-central Al/,ertcr: Res. Council Alberta, Bull. n. llj, 2~9 p. SOMERS, G., 1952, A preliminary study of the fossil spore content of the Loic~er J//bilee seam of the S~,dney coalfield, Nr~zia Scotia: Nova Scotia Res. Fdn., p. 1-30. SUKH DEV., l')Gl, The fossil flora of thee Jubalpicr series. 1!1, Spores and polleN Krains. Palaeobotanist, v. 8, p. 4~-5~. THIERGART, F., 1~49, D~r stratiRraphischer l~crt 7rresozoischer ·Pollen und Sporen: Palaeontographica, B1 v~ 89, p. 1-34. THOMSON, P. W. and PFLUG, H., l~f~3, Pollen und Sporen deJ· mitteleuvopaischen TL·rtik'vs .· Palaeontographica, B, v. 94, p. 1-138. TRAVERSE, A., 13r8, Locating plant microfossils on mixed slides: Micropaleontology, v. 4, n. 2, p. 207-208. TRAVERSE, A., AMES, H. T., and SPACKMAN, W., 1974, McJozoic Pollen and spores: Cat. Fossil Spores, Pollen, v. 38, p. 1-246. VAKHRAMEEV, V. A., 1970, Range and palaeoecology of Mesozoic conifers, The Cheirolepida ceae: Paleont. Zhur., n. 1, p. 1~-34 (in Russian). Translation, Pafeont Jour., 1370, n. 1, p. 12-2~. VAN HOUTEN, F. B., 1969, Lrrte Triassic Newark Group, North Central New Jersey and adjacent Pennsylvania and Ncu, Yor&= in Geology of Selected Areas in New Jersey and Eastern Pennsylvania and Guidebook of Excursions, Field Trip No. 4, Subittky, S. (ed.), Geol. Soc. Amer., 1969 Ann. Meet., New Brunswick, N. j., Rutgers Univ. Press, p. 314-~47. VENKATACHALA, B. S., 1366, Mesozoic oPerctllate Pollen and their m~phology: Palaeobotanist, v. 1T, p. 88-101. VENKATACHALA, B. S. and- GOCZAN, F., 1964, The spore-pollen pora of the Hungarian "Kiissen facies": Acta Geol. Acad. Sci. )-lungary, v. 8, p. 203-228. VOLKHEIMER, W., 1971. Algt~nos adelantos en la microbioestratiRrlrfia del JNrasico en la Argentina y com~racirin con otras regidnes del Hemisferio A//stral, Ameghiniana, v. 8, n. 3, 4, p. 34~-3~5. ---- 1972, E.crudio palinoldRico de Iln carbo'n Crlloviano de NczlRElen y conJideracidnes sobve los paleo. climas J~rbsicos de la Argentina: Rev. Mus. La Plam, n.s., Paleont., v. 6, p. 101-1r7. WALL, D., 1965, Microplankton, pollen, and spores from the Lower _Iurassic in Britain: Micropaleontology, n. 2, p. 1~1-190. WILLS, L. J., 1970, The Tviassic szcccession in the central Midlands in its regional setting: Geol. Sec. London, Quart. Journ., v. 126, p. 22~-28~. WILSON, L. R. and WEBSTER, R. M., 1946, Plant microfossils from a Fort Union coal of Montana. Amer. Jour. Bet., v. 33, p. 271-2 78. WODEHOUSE, R. P., 13~~~ Tertiary pollen. II, The oil shaleJ of the Er,cene Green River formation: Torrey Bet. Club, Bull., v. 60, p. 47~-524.