JEC_1795_sm_AppendixS3
advertisement

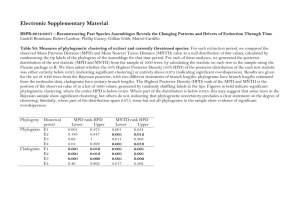
Appendix S3 Supplemental methods, results and discussion pertaining to the assembly of plant communities along an environmental gradient in the Serengeti ecosystem. Supplementary Methods Laboratory methods – Accessions of each species were collected from Serengeti and voucher specimens were deposited at NHT; however, because numerous specimens of each species were collected as part of the study, the voucher specimen deposited at NHT may not be the same individual from which DNA was extracted. DNA was extracted from leaves using the DNeasy Plant Mini Kit (Qiagen, Valencia, California, USA). The polymerase chain reaction (PCR) was performed using Eppendorf Mastercycler gradient or Mastercycler personal thermal cyclers in 50 µl volumes with the following reaction components: 1 µL template DNA (~ 10-100 ng), 1X ExTaq buffer (PanVera / TaKaRa, Madison, Wisconsin, USA), 200 µmol/L each dNTP, 3.0 mmol/L MgCl2, 0.1 µmol/L each primer, and 1.25 units ExTaq (PanVera / TaKaRa). Reactions included bovine serum albumin at a final concentration of 0.2 µg/µL, which is known to improve amplification from difficult templates. The PCR parameters for the psbA-trnHGUG intergenic spacer region were preceded by an initial denaturation phase of 80°C for 5 min followed by 35 cycles of denaturation at 94°C for 30 s, annealing of primers at 50°C for 30 s, and extension of new strands at 72°C for 1 min). Each run also included a final extension at 72°C for 10 min. The following primers were used for both PCR and sequencing: trnHGUG (CGC GCA TGG TGG ATT CAC AAT CC) (Tate & Simpson 2003) and psbA (GTT ATG CAT GAA CGT AAT GCT C) (Sang et al., 1997). Because the average length of this region is relatively short (~550 bp), only the trnH primer was used in sequencing in most cases. PCR and external sequencing primers for the rpL16 intron, rpL16F71 (GCT ATG CTT AGT GTG TGA CTC GTT G) and rpL16R1516 (CCC TTC ATT CTT CCT CTA TGT TG), are from Shaw et al. (2005). The PCR thermal cycling parameters for this gene region were preceded by template DNA denaturation at 80°C for 5 min and followed by a final extension step of 5 min at 65C. The PCR cycling conditions were 30 cycles of denaturation at 95C for 1 min, primer annealing at 50C for 1 min, followed by a ramp of 0.3C/sec to 65C, and primer extension at 65C for 4 min. PCR products were checked on 1% agarose gels before being cleaned with ExoSAPIT (USB, Cleveland, Ohio, USA). In a few species, polyA/T regions prohibited clean sequencing of the rpL16 intron with the external primers. Additionally, we could not get the internal sequencing primers of Zhang (2000) to work; therefore, four internal sequencing primers were created based on the alignment of all of the sequences in a preliminary data set containing species from all three subfamilies. The sequences of the internal primers were: rpL16A (ATA GTT GTA GCA ACT GCA), rpL16B (TGC AGT TGC TAC AAC TAT), rpL16C (GAT TCA AAC CTA ACC ATT), and rpL16D (AAT GGT TAG GTT TGA ATC). Primers rpL16A and rpL16C both sequence toward rpL16R1516 and primers rpL16B and rpL16D sequence toward rpL16F71. All DNA sequencing was performed with the ABI Prism BigDye Terminator Cycle Sequencing Ready Reaction Kit, v. 3.1 (Perkin-Elmer / Applied Biosystems, Foster City, California, USA) and electrophoresed and detected on an ABI Prism 3100 automated sequencer (University of Tennessee Molecular Biology Resource Facility). Sequencher 4.7 (Gene 1 Codes) was used to edit the DNA strands. All sequences listed in Appendix 2 are being deposited in GenBank (accession numbers are pending). Data analysis and phylogenetic assessment – Alignment of DNA sequences was initially performed with ClustalX (Thompson et al. 2001), with subsequent manual adjustment by eye and cpDNA region concatenation in Mesquite v. 2.6 (Maddison & Maddison, 2009). Variable positions in the data matrix were double checked against the original chromatogram files using Sequencher 4.8 (GeneCodes Corp., Ann Arbor, Michigan, USA) to make sure that all base calls were true at all variable positions. In all cases, alignment of potentially informative positions was unambiguous. Because indels have been shown to provide approximately one-third of the potentially informative characters in a cpDNA data set (Shaw et al. 2005), indels were coded as binary characters except in the case of one poly-A/T run toward the trnH end of psbA-trnH and six separate polyA/T regions in the rpL16 intron. All of these poly-A/T regions were omitted from the data set as they may be PCR artifact and not reflective of the phylogenetic history of the group or because alignment of these regions was ambiguous. Parsimony and Bayesian analyses were conducted for the same taxon sample using the concatenated data set. Analysis of phylogenetic relationships was first conducted using the optimality criterion of maximum parsimony. Searches for mostparsimonious trees were executed in PAUP* v. 4.0 b10 (Swofford 2002) by a heuristic search with tree bisection-reconnection (TBR) branch swapping and 1000 random sequence addition replicates with the “collapse zero-length branches” option in effect. Bootstrap support (Felsenstein 1985) was estimated with 1000 replications of heuristic search and simple taxon addition with the constraint of ten million rearrangements per replicate. Both the consistency and retention indices (CI and RI, respectively) were used to assess the amount of homoplasy present in the data set. The maximum likelihood model, GTR + I + G was selected by Mr.Modeltest2 (Nylander 2004), which is a simplified version of MODELTEST 3.06 (Posada & Crandall 1998), using the Akaike information criterion (AIC). Bayesian analyses were performed with MrBayes 3.1.2 (Huelsenbeck & Ronquist 2001). No a priori assumptions about tree topology were made. The Monte Carlo Markov Chain (MCMC) process was set to run ten million generations with two runs of four chains each. Aristida adoensis (subf. Arundinoideae) was chosen as the outgroup for all analyses because this lineage is a sister to all other grass species within this study (Matthews et al. 2000; Zhang 2000). Characterization of the cpDNA data set – For the analyses, sequence data were obtained from two different noncoding cpDNA regions, the psbA-trnH intergenic spacer and the rpL16 intron. As expected, given the complete linkage among cpDNA regions, data derived from different cpDNA regions were congruent and thus were combined into a single data set. The combined cpDNA data set consisted of 2006 aligned nucleotide positions (with several large gaps opened to conservatively align regions that were very divergent or contained repetitive sequence motifs). Within the combined data set there were 289 parsimony informative characters (including multi-bp indels coded as binary characters) and 175 parsimony uninformative characters. The aligned data set of psbAtrnH consisted of consisted of 480 bp from the psbA end of the ~550 bp intergenic spacer region. Within the aligned matrix of this region there were 54 parsimoniously 2 informative characters (including multi-bp indels coded as binary characters), 28 variable but parsimony uninformative characters, and 398 constant characters. The aligned data set of the rpL16 intron consisted of 1526 characters, with several multi-bp gaps opened up to conservatively call some regions where alignment was impossible. Within this aligned data set there were 235 parsimony informative characters, 147 variable but parsimony uninformative characters, and 1144 constant characters. Details on a priori model and structural equation modeling (SEM) – Structural equation modeling is a multivariate statistical approach that allows one to tests observed data against an a priori model specified by the analyst (Grace 2006). Our analysis was driven by a set of a priori models that incorporated environmental covariation and the effects, both direct and indirect, of environmental variable and traits on mean phylogenetic distance. The four general a priori models (A-D) driving the analysis are shown in Fig. 1. In the actual analysis the boxes labeled ‘environmental factors’ and ‘trait distance’ are actually represented by multiple indicators: rainfall, proportion dry season rainfall and elevation in the case of ‘environmental variables’ and average distance in SLA and average distance in maximum plant height in the case of ‘trait distance’. The presence of a solid arrow in Fig. 1 suggests that direct effects from one variable to another should be present under that particular model; no hypotheses are made about the direction (i.e. positive or negative) of paths because they depend on specifics such as the particular environmental variable and evolutionary trait relationships. The simplest model (not shown) is a null model, in which none of our selected variables explain variation in MPD. Model A occurs when the relationship between environmental factors and MPD is indirect; under this model, the effect of environmental variation on MPD is mediated through the direct effect of species richness on MPD. Model B occurs when environmental variables, but not trait relationships or species richness, directly explain variation in MPD. Reasonable explanations for this model include the measurement of incorrect traits or a spatial/historical biogeographic effect which modifies the pool of regionally available species. Model C occurs when environmental variation controls trait relationships and traits directly affect MPD. Model C is most consistent with support for phylogenetic community assembly theory. Finally, Model D represents some combination or composite of the previous models. Because all our measures of environmental variation occur at the site level rather than at the scale of the 1m2 plots (i.e. average annual rainfall, proportion dry season rainfall and elevation), the data were aggregated to the site level for the SEM analysis. Therefore, the SEM analysis was conducted using an n = 133 sites rather than 1330 individual 1 m2 plots. This is desirable because within-site variation in MPD is essentially unexplainable within the context of an SEM in which no within-site data are available. Analysis of all 1330 plots separately would artificially inflate the samples sizes and create a case of pseudoreplication that would invalidate the assessment of individual path strengths. Evaluation of the a priori models (Fig. 1.) with SEM was based on maximum likelihood procedures and was conducted in AMOS version 17.0 (Arbuckle 2007). Models were fit iteratively using a model-pruning strategy in which non-significant paths ( > 0.05) were trimmed from the model until only significant paths remained. In addition, modification indices with a threshold value set to 4.0 were used in AMOS to 3 identify missing paths that substantially improved the model. Model fit was assessed using the χ2 statistic and the associated P-value. A model was deemed adequate when the observed and expected covariances were not significantly different from one another based on the critical P-value of > 0.05. Supplementary Results Phylogenetic results – The topology of the trees generated with the Bayesian and maximum parsimony analyses were congruent; therefore, we are only presenting the Bayesian tree (Fig. 2). In the maximum parsimony analysis, the heuristic search found 112 equally parsimonious trees with 838 steps and consistency and retention indices of C = 0.70 and R = 0.89, which indicate a small amount of homoplasy in the data set. Bootstrap values are shown below the branches in Fig. 2. The posterior probability support values, generated in MrBayes, were consistently higher for equivalent tree branches compared to the parsimony analysis, which is commonly the case. Posterior probabilities are shown above the branches in Fig. 2. One difference between topologies generated by the separate analyses was that in the parsimony analysis Sporobolus rangei was shown to be sister to the clade containing S. kentrophyllus through S. fimbriatus. A second difference between the two analyses was that in the maximim parsimony analysis Enteropogon macrostachyus was sister to the clade containing E. paspaloides through C. roxburgiana + C. gayana. Comparing the statistical fit of the various SE models – The final accepted model demonstrated a superior fit to the alternative models (Fig. 1) and was most consistent with Model C (2 = 5.88, df = 5, p = 0.32, AIC = 51.88, BIC = 118.36; where AIC is Akaike’s Information Criterion and BIC is the Bayesian information criterion). To assess the fit of Model A the path between grass species richness and MPD was kept in the model despite being non-significant (p = 0.35), which yielded a poor fit to the data (2 = 36.79, df = 7, p < 0.001, AIC = 78.79, BIC = 139.49). For Model B, all paths were significant, but no paths were included from trait distances to MPD (2 = 22.16, df = 6, p = 0.001, AIC = 66.16, BIC = 129.75). Model D was tested by including paths from trait distances, grass species richness and environmental factors directly to MPD. This model included several non-significant paths, including from rainfall --> MPD (p = 0.37), proportion dry season rainfall --> MPD (p = 0.37), and grass species richness --> MPD (p = 0.08), yielding a model with a lower chi-square but larger AIC and BIC then Model C (2 = 1.65, df = 2, p = 0.44, AIC = 53.65, BIC = 128.80). Supplementary Discussion Comparison of the phylogeny with previous work – The phylogeny (Fig. 2) was highly consistent with other published works, as described below. All of the species included in this study were from three subfamilies within plant family Poaceae, subfamilies Arundinoideae, Chloridoideae, and Panicoideae, and most species were from the latter two subfamilies. Within subfamily Arundinoideae, only Aristida adoensis was sampled and this species was used to root the rest of the phylogeny, which consisted of species from subfamilies Chloridoideae and Panicoideae. Our results support earlier works that showed the Chloridoideae and Panicoideae to be monophyletic subfamilies (Grass Phylogeny Working Group 2009). As is apparent in Fig. 2, the species of subf. 4 Panicoideae segregate into two tribes, tribe Paniceae and tribe Andropogoneae. The relationships shown among the taxa of tribes Andropognoeae and Paniceae are perfectly consistent with the results of Aliscioni et al. (2005) who used chloroplastic ndhF sequences to derive a molecular phylogeny of Panicum. Within subfamily Chloridoideae our results are also largely congruent with other molecular studies (Hilu & Alice 2001, Roodt-Wilding & Spies 2006). Species within this subfamily segregate into two clades that strongly agree with taxonomic relationships shown by Roodt-Wilding & Spies (2006) in which the tribes Eragrostideae and Cynodonteae are paraphyletic and therefore referred to as the ‘Eragrostideae’ and ‘Cynodonteae.’ For example, in agreement with Roodt-Wilding and Spies (2006), we show that Dactyloctenium, Oropetium, Leptochloa, Eleusine and Tragus, which are all members of tribe Eragrostideae, segregate with tribe Cynodonteae species (Fig. 2). REFERENCES Aliscioni, S.S., Giussani, L.M., Zuloaga, F.O. & Kellogg, E.A. (2003) A molecular phylogeny of Panicum (Poaceae: Paniceae): tests of monophyly and phylogenetic placement within Panicoideae. American Journal of Botany, 90, 796-821. Arbuckle, J.L. (2007) Amos 17.0 update to the Amos user’s guide. Smallwaters Corporation, Chicago, Illinois, USA. Felsenstein, J. (1985) Confidence limits on phylogenies: an approach using the bootstrap. Evolution, 39, 783-791. Grass Phylogeny Working Group, Barker, N.P., Clark, L.G., Davis, J.I., Duvall, M.R., Guala, G.F., Hsiao, C., Kellogg, E.A. & Linder, H.P. (2001) Phylogeny and Subfamilial Classification of the Grasses (Poaceae). Annals of the Missouri Botanical Garden, 88, 373-457. Grace, J.B. (2006) Structural equation modeling and natural systems. Cambridge University Press. Hilu, K.W. & Alice, L.A.. 2001. A phylogeny of Chloridoideae (Poaceae) based on matK sequences. Systematic Botany, 26, 386-405. Huelsenbeck, J.P. & Ronquist F. (2001) MrBayes: Baysian inference of phylogenetic trees. Bioinformatics, 17, 754-755. Matthews, S., Tsai, R.C. & Kellogg, E.A. (2000) Phylogenetic structure in the grass family (Poaceae): Evidence from the nuclear gene phytochrome B. American Journal of Botany, 87, 96-107. Madison, W.P. & Madison, D.R. (2008) Mesquite: a modular system for evolutionary analysis, ver. 2.5, website: http://mesquiteproject.org/mesquite/. Nylander, J.A.A. (2002) Testing models of evolution—MrModeltest 1.1b. Computer program and documentation distributed by author, website: http://www.ebc.uu.se/systzoo/staff/nylander.html. Peterson, P.M., Romaschenko, K. & Johnson, G. (2010) A classification of the Chloridoideae (Poaceae) based on multi-gene phylogenetic trees. Molecular Phylogenetics and Evolution, 55, 580-598. Posada, D. & Crandall, K.A. (1998) MODELTEST: testing the model of DNA substitution. Bioinformatics, 14, 817-818. 5 Roodt-Wilding, R. & Spies, J.J. (2006) Phylogenetic relationships in southern African chloridoid grasses (Poaceae) based on nuclear and chloroplast data. Systematics and Biodiversity, 4, 401-415. Sang, T., Crawford, D.J. & Stuessy, T.F. (1997) Chloroplast DNA phylogeny, reticulate evolution, and biogeography of Paeonia (Paeoniaceae). American Journal of Botany, 84, 1120-1136. Shaw, J., Lickey, E.B., Beck, J.T., Farmer, S.B., Liu, W., Miller, J., Siripun, K.C., Winder, C.T., Schilling, E.E. & Small, R. (2005) The tortoise and the hare II: relative utility of 21 noncoding chloroplast DNA sequences for phylogenetic analysis. American Journal of Botany, 92, 144-166. Shaw, J., Lickey, E.B., Schilling, E.E. & Small, R. (2007) The tortoise and the hare III: comparison of whole chloroplast genome sequences to choose noncoding regions for phylogenetic studies in angiosperms. American Journal of Botany, 92, 142166. Swofford, D.L. (2002) PAUP*: phylogenetic analyses using parsimony (*and other methods) version 4.0b10. Sinauer Associates, Sunderland, Massachusetts, USA. Tate J.A. & Simpson, B.B. (2003) Paraphyly of Tarasa (Malvaceae) and diverse origins of the polyploid species. Systematic Botany, 28, 723-737. Thompson, J.D., Higgins, D.G. & Gibson, T.J. (2001) ClustalX. Program available at ftp://ftp-igbmc.u-strasbg.fr/pub/clustalx/. Zhang, W. (2000) Phylogeny of the grass family (Poaceae) from rp/16 intron sequence data. Molecular Phylogenetics and Evolution, 15, 135-146. 6