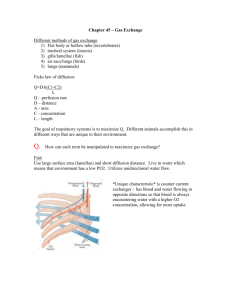
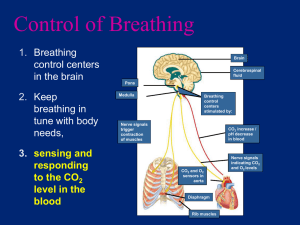
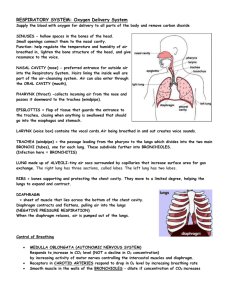
Chapter 42 b: Gas Exchange in Animals What Physical Factors Govern Respiratory Gas Exchange? • The respiratory gases that animals must exchange are oxygen and carbon dioxide. • Adequate ATP production for nutrient molecule oxidation in cells requires oxygen from the environment. • Oxidative metabolism (cellular respiration) in cells produces carbon dioxide as an end product, which must be lost to the environment to prevent toxic effects. Gas exchange occurs across respiratry surfaces by diffusion Diffusion is driven by concentration differences • Biologists express the concentrations of different gases in a mixture by partial pressures of those gases. • Partial pressure of oxygen is 21% and Partial pressure of carbon dioxide is .03% • The solubility of a gas is another factor that influences the concentration of respiratory gases in a liquid. Fick’s law applies to all systems of gas exchange • Fick’s law of diffusion summarizes the rate at which a substance such as O2 diffuses between two locations. • Fick’s law can be expressed as an equation: Q = DA (P1 – P2/L) • Q is the rate at which a substance diffuses between two locations. • D is the diffusion coefficient; it depends on the diffusing substance, the medium, and the temperature. • A is the cross-sectional area over which the substance is diffusing. • P1 and P2 are the partial pressures of the gas at two locations. • L is the distance between these locations.. Other adaptations for maximizing respiratory gas exchange must influence the surface area for exchange (A) or the partial pressure gradient across that surface area [(P1 – P2)/L]. Thin membranes shorten the diffusion path (L). Ventilation brings in fresh air with the high PO2 and the low PCO2. Perfusion by the circulatory system helps maintain the low PO2 and the high PCO2 on the inside of exchange surfaces Air is a better respiratory medium than water • It is easier to obtain O2 from air than from water. • The O2 content in air is about 20 times higher than in water and O2 diffuses 8,000 times more rapidly in air. • Breathing air, which is less dense and less viscous than water, requires less work for an animal. ••Diffusion of O2 in water is so slow that even cells with low metabolism must be only 1–2 mm from the O2 source. • Animals that have no internal systems for distributing O2 are either severely limited in size or have evolved bodies that are flat, hollow, or have specialized structures. High temperatures create respiratory problems for aquatic animals • However, warm water holds less dissolved O2 than cold water. • Therefore, more energy must be expended to get a decreasing O2 supply. O2 availability decreases with altitude • Air has 20.9 percent O2. • At the top of Mount Everest, the PO2 is only about 50 mm Hg. • The diffusion rate of O2 into cells under reduced PO2 is greatly reduced, and O2 uptake is constrained. • Humans venturing to great heights must breathe O2 from pressurized containers. Insects have airways throughout their bodies • The tracheal system of insects enables them to exchange respiratory gases to all tissues in the body. • This system of air tubes begins at openings on the outside of the body called spiracles, which admit air. Fish gills use countercurrent flow to maximize gas exchange • Gills in fish are constructed to enable water to pass into the mouth, over the gills, and out the opercular flaps • This constant water flow over the gills maximizes the PO2 on the external surfaces. • Blood flow on the internal side minimizes the PO2 by sweeping O2 away as rapidly as it diffuses across the gill. • Highly divided, the gills present enormous surface area for gas exchange. • The subunits of gill filaments and lamellae minimize the path length for diffusion. • The perfusing blood flow on the inner surface of the lamellae is unidirectional. • Afferent (to gills) and efferent (away from gills) blood vessels ensure a countercurrent (opposite direction) flow to maximize the PO2 gradient. • Some fish, such as sharks and tuna, swim almost constantly with mouths open to ventilate their gills. • Most fish use a two-pump mechanism activated by opening and closing of the mouth to push water over the gills. Birds use unidirectional ventilation to maximize gas exchange • Birds can sustain high activity levels much longer and at higher altitudes than mammals can. • This ability is achieved through a unique structure allowing air to flow unidirectionally through the lungs rather than bidirectionally through the same airway, as in mammals. • Mammalian lungs never completely empty of air during exhalation. • Birds also have air sacs in the body and air spaces within the wings to hold air but not exchange it. • In most air-breathing vertebrates, air enters the trachea, and then passes to the bronchi, to smaller bronchioles, and finally to dead-end membranous air sacs where gases are exchanged. • In birds, the trachea leads to air sacs, then to parabronchi connected by air capillaries in the lungs where gas exchange occurs. • Air next passes to bronchi and to other air sacs, which then vent back to the trachea, bypassing the lungs. • This arrangement ensures a unidirectional flow of fresh air through the lungs. •Tidal ventilation produces dead space that limits gas exchange efficiency • In mammal lungs, ventilation is tidal: air flows in and out by the same route. • A spirometer is a device that measures the volumes of air that a person breathes in or breathes out. • Measurements of lung capacity: • At rest, the amount of air that moves in and out is the tidal volume. • The additional volume of air that we can take in by inhaling deeply is the inspiratory reserve volume. • The additional volume we can exhale is the expiratory reserve volume. • The total of these three volumes is the vital capacity. • Even with forceful breathing, there is residual volume keeping the lungs from collapsing. Some of this exists in what is called the anatomical dead space— airways in which gas exchange cannot occur. • The total lung capacity is the sum of the residual volume and the vital capacity. • Tidal breathing severely limits the PO2 gradient because fresh air is not moving into the lungs during half of the respiratory cycle. • The incoming air also must mix with the stale air remaining in the lungs. • The volume of this stale air is the sum of the residual volume and, depending on how deeply one is breathing, some or all of the expiratory reserve volume. • Tidal breathing also reduces gas exchange efficiency by not permitting countercurrent gas exchange between air and blood. • To offset the inefficiencies of tidal breathing, mammalian lungs have an enormous surface area and a very short path length for diffusion. How Do Human Lungs Work? • Figure 48.10 details the human respiratory system. • The air pathway in humans consists of the following components: • An oral or nasal cavity, followed by the pharynx (an area for both food and air). • The larynx (voice box), which leads to the trachea. • The trachea, which branches into two bronchi (both of these have cartilage support). • The bronchi, which branch repeatedly into bronchioles. • The bronchioles, which terminate in the alveoli. • The alveoli, which are thin-walled air sacs and are the sites of gas exchange. • There are about 300 million alveoli in human lungs.) • The proliferation of alveoli gives a gas exchange surface area one-fourth the size of a basketball court. • Capillary blood vessels closely surround the air sacs, resulting in a diffusion path of less than 2 um, which is less than the diameter of a red blood cell. • Respiratory tract secretions aid ventilation • Two adaptations that aid the breathing process in mammals are mucus production and surfactant production. • Mucus production: • Cells lining the airways produce a sticky mucus that captures dirt and microbes that might be inhaled. • This mucus is cleared by cilia beating upward toward the trachea and pharynx, where it is swallowed. • This phenomenon has been called the mucus escalator, and it can be immobilized by smoking. • The genetic disease cystic fibrosis causes respiratory problems by affecting the respiratory mucus. • Surfactant production: • A surfactant is a chemical substance in the lungs that reduces the surface tension of liquid preventing them from collapsing. • The aqueous lining of the lung has surface tension that must be overcome to permit inflation. • Lung surfactant is a fatty substance that is critical for reducing the work necessary to inflate the lungs. • Cells in the alveoli produce surfactant molecules when they are stretched. • Premature babies may develop respiratory stress syndrome if they are born before cells in the alveoli are producing surfactant. Lungs are ventilated by pressure changes in the thoracic cavity • The human lungs are suspended in the thoracic cavity. • This cavity is bounded by the shoulder girdle at the top, the rib cage on the sides, and the diaphragm muscle on the bottom. • The lungs are positioned in a separate, closed pleural space. • Breathing involves changes in volume of the thoracic cavity. • An increase in its volume creates negative pressure (suction) inside the pleural space. • Even between breaths, there is a slight negative pressure inside the pleural space keeping the alveoli partially inflated. If the thoracic cavity is punctured, air leaks into the pleural space, the lung collapses, and ventilation of the alveoli in the lung ceases. • With inhalation, the diaphragm muscle contracts downward to cause suction, and air flows into the lung. • When the diaphragm stops contracting and relaxes, pushing upward, exhalation occurs. • The diaphragm is not the only muscle that changes the volume of the thoracic cavity: • In the rib cage, intercostal muscles lift the ribs up and down to increase thoracic cavity volume. • External intercostal muscles expand the thoracic cavity and increase the volume of air inhaled. • Internal intercostal muscles decrease the volume of the thoracic cavity and increase the amount of air exhaled. • This system is particularly important during strenuous exercise. How Does Blood Transport Respiratory Gases? • Remember that ventilation and perfusion work together. Ventilation delivers O2 to the environmental side of the exchange surface, where it diffuses into the body and is swept away by perfusion; perfusion delivers CO2 to the exchange surface, where it diffuses out and is swept away by ventilation. • Perfusion of the lungs is one of the functions of the circulatory system, which transports respiratory gases through a network of blood vessels powered by a pump (the heart). • As O2 diffuses from the alveoli into the blood, it is swept away and delivered to the cells and tissues of the body. • Blood plasma (the liquid part of blood) carries only a small portion of O2 in solution. • It is the red blood cell with its oxygen-binding pigment, hemoglobin, that is the pack-horse for oxygen transport. • Hemoglobin has 60 times the capacity of plasma to transport O2. • Hemoglobin combines reversibly with oxygen • Hemoglobin is a protein consisting of four polypeptide subunits. Each polypeptide surrounds a heme (iron-containing) group. • Each heme group can reversibly bind a molecule of O2. • As diffusion of O2 into the blood occurs, it binds to hemoglobin, increasing the PO2 gradient and driving O2 into red blood cells. • When the PO2 of blood plasma is high, as in the lung capillaries, each hemoglobin complex can carry four molecules of O2. • As the red blood cell circulates to the body, the PO2 values drop, and the hemoglobin releases some of the O2 it is carrying. • The relationship between saturation of the hemoglobin polypeptides and PO2 values is complicated and follows a sigmoidal curve. • The O2 binding by hemoglobin polypeptides is influenced by positive cooperativity; that is, binding the first molecule makes the second binding easier, and so on. • It takes a relatively greater PO2 to achieve 100 percent saturation of all four polypeptides. • Carbon monoxide (CO) binds to hemoglobin with a much higher affinity than does O2. • Therefore, CO is a deadly poison, as it destroys the ability of hemoglobin to transport and release O2 to body tissues. • When blood circulates through the body, it releases, on average, only one of the four molecules of O2 it carries. • Although this seems inefficient, it is really adaptive because the hemoglobin keeps a 75 percent O2 reserve for peak demands. • If a tissue is starved for O2 and its local PO2 falls below 40 mm Hg, the hemoglobin will release the reserved O2 to the starved tissue. Myoglobin holds an oxygen reserve • Myoglobin in muscle cells is an oxygen-binding molecule that can take up one molecule of O2. • It has a higher affinity for O2 than hemoglobin does and provides an oxygen reserve for high metabolic demand or when blood flow is interrupted. • Diving mammals such as seals have high concentrations of myoglobin in their muscles, allowing them to stay under water for long periods using their reserves. • Even in nondiving animals, muscles called on for extended periods of work frequently have more myoglobin than muscles used for short, intermittent periods do. The affinity of hemoglobin for oxygen is variable • Various factors influence the oxygen-binding properties of hemoglobin, including the chemical composition of the hemoglobin, pH, and the presence of 2,3-bisphosphoglyceric acid. • Hemoglobin composition: • Hemoglobin of normal human adults consists of two kinds of polypeptide chains: • Before birth, human fetuses have different globin chains • These globin chains result in a greater affinity for O2 in fetal hemoglobin. • This difference in O2 affinity facilitates transfer of O2 from the mother’s blood to fetal blood in the placenta. • Mammals such as llamas, which live at high altitudes, have hemoglobin that becomes saturated with O2 at lower PO2 values than that of other animals. • Hemoglobin and pH: • The influence of pH on the function of hemoglobin is known as the Bohr effect. • This effect holds when the pH of the blood falls and the oxygen-binding curve shifts to the right. • The hemoglobin will then release more O2 to the tissues. • Another regulator of hemoglobin function is 2,3-bisphosphoglyceric acid (BPG), a metabolite of glycolysis. • Mammalian red blood cells have a high concentration of BPG (increases in exercise), which combines with deoxygenated hemoglobin and causes it to have a lower affinity for O2. • The result is that the hemoglobin releases more of its bound O2 to tissues than usual. Carbon dioxide is transported as bicarbonate ions in the blood • Carbon dioxide (CO2), a metabolic waste, must be taken away from living tissues by the blood. • CO2 is highly soluble, moving easily through cell membranes into the blood, where the partial pressure of CO2 is lower. • However, most CO2 is transported via blood to the lungs as bicarbonate ion (HCO3–). (See Figure 48.14.) • The endothelial cells of capillaries and the red blood cells produce carbonic anhydrase enzyme, which speeds the conversion of CO2 to H2CO3 (carbonic acid). • The H2CO3 dissociates, and bicarbonate ions enter the plasma in exchange for chloride ions (Cl–). • This conversion reduces the partial pressure of CO2 in these cells and in plasma, promoting the diffusion of CO2 out of the tissue cells. • Although most CO2 goes to the lungs as bicarbonate, some CO2 is also carried in chemical combination with hemoglobin. • In the lungs, the CO2 and bicarbonate reactions are reversed. • Carbonic anhydrase speeds the conversion of bicarbonate to water and CO2. • CO2 diffuses from the blood plasma into the alveolar air and is exhaled. • As the partial pressure of CO2 in the blood falls, more bicarbonate is converted into CO2. How Is Breathing Regulated? Breathing is controlled in the brain stem • The autonomic nervous system maintains breathing and modifies depth and frequency to meet body demands. • The brain stem generates and controls the breathing rhythm. • Groups of neurons within the medulla increase their firing rate just prior to inhalation. • With increased firing, the diaphragm contracts, and inhalation occurs. • Suddenly, the firing stops, the diaphragm relaxes, and exhalation occurs. • Exhalation is actually a passive elastic recoil of lung tissue. • When breathing demands are high, such as during exercise, the motor neurons for the intercostal muscles are fired to increase inhale/exhale volumes. • Brain areas above the medulla modify breathing to allow speech, eating, coughing, and emotional states. Regulating breathing requires feedback information • Sensors measuring the PO2 and PCO2 in the blood provide feedback to regulate the breathing rhythm. • In humans and other mammals, CO2 sensitivity is very high, but O2 sensitivity is remarkably low. • This relationship is reversed for water-breathing animals, in which oxygen is the primary feedback stimulus for breathing. • Figure 48.17 shows how the sensitivity of the respiratory control system changes with exercise. • CO2 sensors are located on the medulla surface near the neurons that generate the breathing rhythm. • O2 sensors also reside in tissue nodes on the aorta and carotid arteries.
 0
0
advertisement
Related documents






Download
advertisement
Add this document to collection(s)
You can add this document to your study collection(s)
Sign in Available only to authorized usersAdd this document to saved
You can add this document to your saved list
Sign in Available only to authorized users