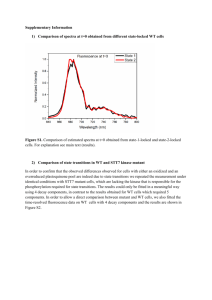
TPJ_4033_sm_Appendix1

Supplementary experimental procedures
Plant materials
Seeds of Arabidopsis thaliana with the as1-1 , as2-1, bp-1 , knat6-2 and spy-1 mutant alleles were obtained from the Arabidopsis Biological Resource Center (ABRC). The knat2-3 allele (SALK_004073) was obtained from the SIGnAL T-DNA express service
(Alonso et al ., 2003). To confirm the positions of insertions, we sequenced the T-DNA left border-flanking DNA for each T-DNA allele after amplification by PCR. The bp-1 allele, originally on the Landsberg erecta background, was outcrossed three times with
Col-0. The new bp allele ( bp-10 ) was isolated as an ethyl methanesulfonate-induced mutation, with a single base-change that resulted in the conversion of the codon for a glutamine residue at position 236, before the homeodomain of BP, to a termination codon. All multiple-mutant lines were identified in segregating populations by
PCR-based genotyping.
Plant genetics
To obtain the as1-1 bp-1 double mutant, we crossed a bp-1 plant with an as1-1 plant.
The as1 bp double homozygote was identified among F2 plants from the characteristic mutant phenotype caused by each individual mutation: the leaf phenotype for as1-1 and the downward-pointing-fruit phenotype for bp-1 . A knat2-3 plant was crossed with a knat6-2 plant to generate the knat2-3 knat6-2 double mutant. The homozygote of knat2-3 and that of knat6-2 were identified with LBb2
(5’-TGAAGGGCAATCAGCTGTT-3’) as T-DNA primer; pU253
(5’-GCCGGTGGGTAGGATTTTCTCTG-3’) and pU213
(5’-GCATAGGGACCTAGGGTGAATGC-3’) as KNAT2 primers; and pU239
(5’-CACAGAGCTTTCAAATAAGTTGCGTGTAG-3’) and pIK122
(5’-CGCTTATTAATTCCGATCCAGAGAA-3’) as KNAT6 primers. To generate the bp-1 knat2-3 knat6-2 triple mutant, a bp-1 plant was crossed with a knat2-3 knat6-2 plant. The as1-1 knat2-3 knat6-2 triple mutant, the as1-1 knat2-3 double mutant and the as1-1 knat6-2 double mutant were isolated from a cross between an as1-1 plant and a knat2-3 knat6-2 plant. To generate the as1-1 bp-1 knat2-3 knat6-2 quadruple mutant, we crossed an as1-1 bp-1 plant with an as1-1 knat2-3 knat6-2 plant. To generate the as2-1 bp-1 knat2-3 knat6-2 quadruple mutant, we crossed an as2-1 plant with a bp-1 knat2-3 knat6-2 plant. The as2 homozygote was identified among F2 plants from the leaf phenotype characteristic of as2 alleles. To obtain the as1-1 spy-1 double mutant, we crossed an as1-1 plant with a spy-1 plant. The spy-1 homozygote was identified with two primers, namely, pIK181 (5’-ACTTTACAAAGACCGTGACAACCTTGATAA-3’) and pIK182 (5’-GAAAACATTAAGCCGCCCTTAATATATATCAACC-3’). The product of amplification by PCR of the mutant allele but not that of wild type was resistant to digestion by Sty
Ι.
Quantification of gibberellin (GA
53
)
Aerial parts of 10-day-old plants were harvested and GA was extracted from approximally 100 mg (fresh weight) of tissue. Quantification of GA
53
was performed as
described previously (Hirano et al ., 2007) with a liquid chromatography-tandem mass chromatography system (UPLC/Quattro Ultima Pt; Waters) with an ODS column
(AQUITY-UPLC BEH-C18, 1.7 µm, 2.1 x 50 mm, Waters).
Construction of 35S:BP and transformation of plants
A cDNA fragment of the BP gene was amplified by PCR and inserted into the pGreen binary vector (Hellens et al ., 2000) with the promoter from the Cauliflower mosaic virus gene for 35S RNA and the terminator region of a gene for nopaline synthase. The construct was introduced into Col-0 plants by Agrobacterium -mediated transformation.
RNA isolation
Total RNA was extracted from aerial parts of 10-day-old plants (Figure 5g and Figure
S2a) or from shoot apices of 12-day-old plants (Figure 6l) with the RNeasy plant mini kit (Qiagen), and poly(A) + RNA was isolated from 1 μg of total RNA with Dynabeads
Oligo(dT) 25 (Invitrogen). First-strand cDNAs were generated with the oligo dT primers
(Invitrogen) and the First-Strand cDNA Synthesis Kit (GE Healthcare).
Accession numbers
Sequence data from this article can be found in the Arabidopsis Genome Initiative or
GenBank/EMBL database under the following accession numbers: BP , At4g08150;
KNAT2 , At1g70510; KNAT6 , At1g23380; AS1 , At2g37630; AS2 , At2g37630;
AtGA20ox1 , At4g25420; and AtIPT7 , At3g23630.
Supplementary references
Alonso, J.M., Stepanova, A.N., Leisse, T.J., Kim, C.J., Chen, H., Shinn, P.,
Stevenson, D.K., Zimmerman, J., Barajas, P., Cheuk, R., Gadrinab, C., Heller, C.,
Jeske, A., Koesema, E., Meyers, C.C., Parker, H., Prednis, L., Ansari, Y., Choy, N.,
Deen, H., Geralt, M., Hazari, N., Hom, E., Karnes, M., Mulholland, C., Ndubaku,
R., Schmidt, I., Guzman, P., Aguilar-Henonin, L., Schmid, M., Weigel, D., Carter,
D.E., Marchand, T., Risseeuw, E., Brogden, D., Zeko, A., Crosby, W.L., Berry, C.C., and Ecker, J.R. (2003). Genome-wide insertional mutagenesis of Arabidopsis thaliana .
Science , 301 , 653-657.
Hellens, R. P., Edwards, E. A., Leyland, N. R., Bean, S., and Mullineaux, P. M.
(2000). pGreen: a versatile and flexible binary Ti vector for Agrobacterium -mediated plant transformation. Plant Mol. Biol ., 42, 819-832.
Hirano, K., Nakajima, M., Asano, K., Nishiyama, T., Sakakibara, H., Kojima, M.,
Katoh, E., Xiang, H., Tanahashi, T., Hasebe, M., Banks, J.A., Ashikari, M., Kitano,
H., Ueguchi-Tanaka, M., and Matsuoka, M. (2007). The GID1-mediated gibberellin perception mechanism is conserved in the Lycophyte Selaginella moellendorffii but not in the Bryophyte Physcomitrella patens . Plant Cell , 19, 3058-3079.