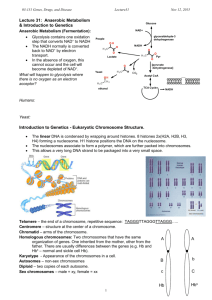
Yeast PAF complex

Supplement information to “Human RNA Polymerase II Associated Factor (hPAF)
Complex: Dysregulation in Cancer”, by Chaudhary et al:
A. Yeast PAF complex
The PAF complex was first identified in yeast (yPAF) as an RNA polymerase IIassociated factor that interacts with TATA Binding Protein (TBP), the elongation factors
Spt4–Spt5, and FACT during transcription elongation {reviewed by (Shilatifard 2006)}
RNAP II-mediated transcription involves successive recruitment of various stage-specific factors and mediators that are required for the formation of a mature elongation complex.
This process is further coordinated by phosphorylation of the carboxy-terminal domain
(CTD) of RNAP II. The yPAF complex acts as a mediator during transcription elongation by stimulating the following steps: Rad6-mediated monoubiquitination of the core histone H2B (Ng et al. 2003; Wood et al. 2003; Wood et al. 2005), Set1-mediated histone
H3-K4 methylation (Dover et al. 2002; Krogan et al. 2003b; Ng et al. 2003), and Dot1mediated histone H3-K79 methylation (Krogan et al. 2003b; Krogan et al. 2003a). FACT, a dimeric protein containing HMG1 (high mobility group) domain plays a major role in this process by binding and displacing the H2A/H2B dimer from the core nucleosome and allowing elongation to proceed over chromatin (Belotserkovskaya et al. 2003)
(Figure 2). The interaction of FACT with the PAF complex is an important step during the process of elongation, and any defects in the PAF complex lead to elongation defects
(Mueller et al. 2004).
To investigate whether all the subunits of the yPAF complex are involved in and interact during the course of transcription, the effects of loss of each subunit was studied.
Isogenic paf1Δ
and ctr9Δ
strains showed severe and nearly identical growth defects, while the Cdc73 or Rtf1 mutants exhibited less severe phenotypes that were a subset of those seen in paf1Δ and ctr9Δ
. The
Leo1Δ
strains had few detectable deficiencies (Betz et al. 2002). These results suggest that Paf1 and Ctr9 have similar roles and are the most important components of the complex.
The yPAF complex also plays essential roles in other cellular functions. The CTD of the RNA polymerase II large subunit contains two important serine residues, Ser2 and
Ser5, which are phosphorylated during transcription. Phosphorylation of Ser5 is required during transcription initiation and the early phase of elongation, while phosphorylation of
Ser2 is linked to elongation (Komarnitsky et al. 2000). Loss of the PAF complex members results in reduced phosphorylation of RNAP II on Ser2, a prerequisite for transcription elongation, and a shortening of poly(A) tails on mRNA. This indicates that the complex acts as a link between transcriptional and posttranscriptional events. Further, the yPAF-RNA polymerase II complex is involved in regulating the expression of a subset of cell-cycle genes. This is evidenced by a 2- to 13-fold reduction in the transcript abundance of many periodically expressed genes, including CLN1, HO, RNR1, and
FAR1 in paf1Δ strain (Porter et al. 2002)
.
B. Rtf1 performs histone methylation independent of PAF complex and regulates
Notch signaling
The human PAF complex differs from the yeast complex in having hSki8 subunit and lacking the yeast Rtf1 homolog. In humans and in Drosophila , the Rtf1 subunit does not appear to be a stable part of the PAF complex. However, Paf1 and Rtf1 seem to associate in the context of actively transcribing RNAP II in Drosophila (Adelman et al. 2006) and dRtf1 plays an important role in histone methylation (Tenney et al. 2006). Thus, the function of this protein is conserved in yeast and Drosophila suggesting that the human
Rtf1 protein may act in tandem with hPAF in histone modification, although not as an integral part of the complex.
Further, Tenney et al. (2006) showed that dRtf1 also plays a role in Notch signaling. The Notch cascade is a crucial signaling pathway that regulates normal development by its effects on cellular differentiation, proliferation and apoptosis.
Alterations in this signaling pathway are associated with various human cancers including T-cell leukemia, which involves translocations of the MLL gene (Weng et al.
2004). The MLL gene is the human homologue of yeast SET1 in the COMPASS
(Complex Proteins Associated with Set1) complex involved in histone methylation. The methylation of histone H3K4 by yeast COMPASS requires ubiquitination of histone H2B by the Rad6 and Bre1 proteins (Figure 2 in the manuscript). As discussed previously, the
PAF complex is required to activate Rad6 and Bre1 histone ubiquitination activity in yeast. Tenney et al. (2006) determined that the Drosophila Rtf1 subunit of the PAF complex facilitates Notch signaling, linking histone ubiquitination and methylation to gene activation by the Notch pathway. Interestingly, Bre1 is also required for Notch target gene expression in Drosophila (Bray et al. 2005). Therefore, the important functional links between the PAF complex, Rad6/Bre1 and histone methylation seen in
yeast is conserved in Drosophilla . Considering the molecular mechanisms underlying
MLL action, it has been predicted by Tenney et al. (2006) that the human PAF complex, along with Rtf1, might participate in Notch signaling and might be linked to the pathogenesis of leukemia via the MLL translocations.
References
Adelman K, Wei W, Ardehali MB, Werner J, Zhu B, Reinberg D et al (2006). Drosophila
Paf1 modulates chromatin structure at actively transcribed genes. Mol Cell Biol 26: 250-
260.
Belotserkovskaya R, Oh S, Bondarenko VA, Orphanides G, Studitsky VM, Reinberg D
(2003). FACT facilitates transcription-dependent nucleosome alteration. Science 301:
1090-1093.
Betz JL, Chang M, Washburn TM, Porter SE, Mueller CL, Jaehning JA (2002).
Phenotypic analysis of Paf1/RNA polymerase II complex mutations reveals connections to cell cycle regulation, protein synthesis, and lipid and nucleic acid metabolism. Mol
Genet Genomics 268: 272-285.
Bray S, Musisi H, Bienz M (2005). Bre1 is required for Notch signaling and histone modification. Dev Cell 8: 279-286.
Dover J, Schneider J, Tawiah-Boateng MA, Wood A, Dean K, Johnston M et al (2002).
Methylation of histone H3 by COMPASS requires ubiquitination of histone H2B by
Rad6. J Biol Chem 277: 28368-28371.
Komarnitsky P, Cho EJ, Buratowski S (2000). Different phosphorylated forms of RNA polymerase II and associated mRNA processing factors during transcription. Genes Dev
14: 2452-2460.
Krogan NJ, Dover J, Wood A, Schneider J, Heidt J, Boateng MA et al (2003a). The Paf1 complex is required for histone H3 methylation by COMPASS and Dot1p: linking transcriptional elongation to histone methylation. Mol Cell 11: 721-729.
Krogan NJ, Kim M, Tong A, Golshani A, Cagney G, Canadien V et al (2003b).
Methylation of histone H3 by Set2 in Saccharomyces cerevisiae is linked to transcriptional elongation by RNA polymerase II. Mol Cell Biol 23: 4207-4218.
Mueller CL, Porter SE, Hoffman MG, Jaehning JA (2004). The Paf1 complex has functions independent of actively transcribing RNA polymerase II. Mol Cell 14: 447-456.
Ng HH, Robert F, Young RA, Struhl K (2003). Targeted recruitment of Set1 histone methylase by elongating Pol II provides a localized mark and memory of recent transcriptional activity. Mol Cell 11: 709-719.
Porter SE, Washburn TM, Chang M, Jaehning JA (2002). The yeast pafl-rNA polymerase
II complex is required for full expression of a subset of cell cycle-regulated genes.
Eukaryot Cell 1: 830-842.
Shilatifard A (2006). Chromatin modifications by methylation and ubiquitination: implications in the regulation of gene expression. Annu Rev Biochem 75:243-69.: 243-
269.
Tenney K, Gerber M, Ilvarsonn A, Schneider J, Gause M, Dorsett D et al (2006).
Drosophila Rtf1 functions in histone methylation, gene expression, and Notch signaling.
Proc Natl Acad Sci U S A 103: 11970-11974.
Weng AP, Ferrando AA, Lee W, Morris JP, Silverman LB, Sanchez-Irizarry C et al
(2004). Activating mutations of NOTCH1 in human T cell acute lymphoblastic leukemia.
Science 306: 269-271.
Wood A, Schneider J, Dover J, Johnston M, Shilatifard A (2003). The Paf1 complex is essential for histone monoubiquitination by the Rad6-Bre1 complex, which signals for histone methylation by COMPASS and Dot1p. J Biol Chem 278: 34739-34742.
Wood A, Schneider J, Dover J, Johnston M, Shilatifard A (2005). The Bur1/Bur2 complex is required for histone H2B monoubiquitination by Rad6/Bre1 and histone methylation by COMPASS. Mol Cell 20: 589-599.