III. E. Methods: Age Structure Analysis
advertisement

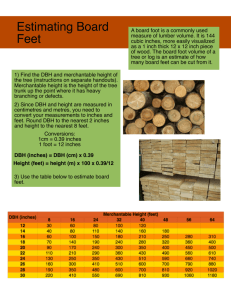
Methods of Analyzing Vegetation Change Part One: A brief introduction to inference from stand structure Possible objectives: Regeneration status (persistent or not persistent) Regeneration mode Reconstruction of stand history Prediction of future composition Required information: 1. The species pool and where species "segregate out along environmental gradients" (i.e. which species can tolerate a particular site). 2. Basic autecological information such as dispersal mechanisms, vegetative reproduction, longevity, seed production and periodicity, etc. 3. Quantitative data on stand structure Types of data collected: i. tree population age structures (sometimes supplemented with size data)=the basic data ii. tree spatial patterns (clumping, associations of size/age classes by species and micro-site) iii. disturbance history (documentary, photographic, dendroecological, etc.) iv. understory data (interference from non-tree species, seedling abundances and micro-sites) v. tree canopy structure (density, cover; estimated or measured) vi. vertical structure (relative tree heights; emergent, dominant, suppressed, etc.) Types of Age Structures Types of age structures include even-aged versus uneven-aged. The uneven-aged class is further divided into intermittent (sporadic), uniform, or “balanced” age structures. See the discussion of this in Veblen 1992. Limitations to Age Structure Analysis A. Confusion of variation in past mortality and past recruitment rates. Static-age structure curves (frequency distributions of numbers of trees in age classes) are shaped both by variations in past recruitment (e.g., past “pulses” of recruitment) and past mortality (e.g., effects of a past drought). This is the fundamental reason why static-age structure curves are not the same as a survivorship curve determined from a fixed cohort life table; tree mortality rates cannot be derived from static-age structure. B. Errors in determining tree ages: 1. Ring-counting errors. 2. Missed pith. 3. Time required to grow to coring height. C. Missing age data due to rotten centers. D. Great sampling effort required to age trees over a large area. Limitations to size (dbh) structure analysis (i.e. substitute tree size for tree age). Restrictions/guidelines for size structure analysis. A. Minimize dependence on size data and maximize collection of age data. B. Test age/dbh relationship by sampling ages of large range of tree sizes in a uniform habitat. C. Adjust the detail of the interpretation (and analytical procedures) according to the strength of the age/dbh relationship. Even if there is a strong relationship between age and dbh (e.g., correlation coefficents of 0.9) you can only make general inferences (e.g., persistence versus non-persistence) from size structure analysis and you cannot reconstruct the details of stand history or reach conclusions about the quantitative stability of the population. D. Interpret data on a stand-by-stand basis before attempting to lump data from dispersed sample sites (likewise this is necessary in age-structure analysis). Problems with lumping result from: age/dbh relationship changes with site; disturbance history changes with site; regeneration modes become obscured; always try to combine age/size analysis with precise dating of disturbance events. Substitution of Space for Time Approaches Chronosequences Many studies combine age structure analysis with the chronosequence approach. A chronosequence depends on the assumption that the sites being compared are ecologically identical and that the only variable that varies among the sites is age or time since disturbance. Natural experiments: Comparison of islands/mainlands. Comparisons of affected and unaffected sites as a substitute for before and after the event. What are the assumptions underlying these approaches?