Report_-_Fermentation_and_EtOH_Recovery
advertisement

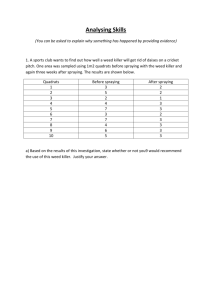
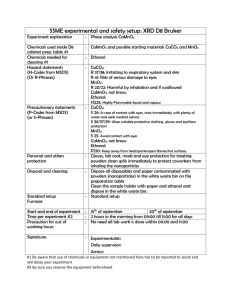
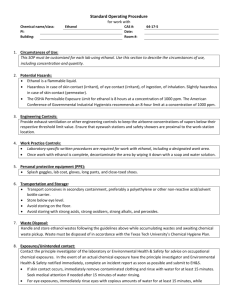
Fermentation (from Lu) Microorganisms used for fermentation must be robust and have a high productivity. Also, the microorganism selected must be able to convert a mixture of different sugars to ethanol. Other favorable traits include high tolerance to ethanol or other inhibitors and resistance to contamination by undesired microorganisms. Three main microorganisms will be considered as possibilities for this fermentation reaction Saccharomyces cerevisiae – S. cerevisaiae has been considered the workhorse for ethanol fermentation for years. This microorganism has high ethanol tolerance and operates at low-pH fermentation conditions. Low pH conditions can be utilized to minimize the potential for contamination. Wild type strains can ferment glucose, mannose and fructose as well as disaccharides like sucrose and maltose, but they cannot ferment pentoses which make up 40% of total biomass carbohydrates. Recombinant DNA technologies have been applied to increase this function. Major enzymes required for xylose fermentation must incorporate xylose flux into the pentose phosphate pathway, these enzymes are xylose reductase and xylitlo dehydrogenase which can be found in yeasts like P. stipitis and Candida shehatae. Additionally, xylose must be transported across the cell membrane. It is believed that this is carried out by glucose transporters. These transporters have a higher affinity for glucose which delays the transport of xylose until the majority of the glucose has been consumed which decreases the effectiveness of the co-fermentation process. This delay results in an increase the time required for fermentation as well as increased capital costs. Zymomnas mobilius – The Entner-Doudoroff pathway is utilized to anaerobically produce ethanol from sugars. Since only one mole ATP is generated per mole sugar consumed, glucose metabolism must be high to compensate for low energy yield which results in high ethanol productivity. Compared to traditional yeast, a 5-10% increase in yield and a 5 fold higher volumetric productivity is observed. Z. mobilius must be metabolically engineered to ferment xylose in addition to hexoses. Some success has been achieved. Major hurdles include industrial adaptation of the recombinant type, increased likelihood of contamination due to operation at neutral pH and less robust cells. Escherichia coli – Wild type E. Coli can ferment a mixture of sugars to acids with ethanol as a minor by-product. Only half the theoretical yield of ethanol can be achieved. By overexpressing pyruvate decarboxylase and alcohol dehydrogenase the ethanol production and tolerance can be increased. Based on this information it seems that using S. cerevisiae will be the best option for this process. It is the most robust microorganism and being able to operate over a wide range of conditions will allow us to optimize the productivity of the fermentation reaction. S. cerevisiae Preparation (from Yong-Su Jin) Inverse metabolic engineering can be utilized to create strains of S. cerevisiae that can ferment xylose in addition to glucose. The recent advances in genome sequencing are taken advantage of using this method. A library of strains is screened for the desired phenotype and then the genetic modifications correlated to that phenotype are determined using sequencing. This approach identify strains that will likely yield the desired phenotype opposed to the older approach of random mutation which are more difficult to trace. Genes were borrowed from Pichia stipitis in order to construct a metabolic pathway through which xylose could be assimilated. Xylose reductase and xylitol dehydrogenase, or XYL1 and XYL2 respectively, were initially chosen to be expressed in S. cerevisiae by Jin et al. These strains had inefficient xylose assimilation and the accumulation of xylitol bounded the ethanol production. With further study, the XYL3 and PsTAL1 genes were found to both improve xylose uptake and ethanol production in S. cerevisiae without inhibiting the yeast’s growth ability on glucose. It was found that strains with these two genes overexpressed were able of rapidly ferment of glucose-xylose mixtures. Strains and Plasmids Media and culture conditions Construction of Pichia stipitis genomic library Yeast transformation Serial transfer and screening of fast-growing transformants Insert identification and sequence analysis Enzymes, primers and chemicals Plasmid construction Preparation of crude extract and enzyme assay Analytical methods Nucleotide sequence accession number ****Ask Professor Hu how much detail he wants on the above bullet points? The genes of interest were transferred to recombinant strains of S. cerevisiae that contained copies of XYL1 and XYL2. These strains were grown up on minimal xylose media for 10 transfers and then plated onto either a glucose or xylose agar plate. The colonies on the glucose plate were of similar sizes, while those on the xylose plate varied greatly indicating that those with the higher growth rate had increase xylose assimilation. (Graphic?) Of 16 plasmids isolated, 10 contained PsXYL3 and one contained an inserted highly homologus to the sequence of ScTAL1. This confirmed the hypothesis that increased xyluokinase activity by either XYL3 or XKS1 would increase the efficiency by which xylose was utilized by S. cerevisiae. It was found that over expression of PsTAL1 would increase the xylose assimiliation while have no effect to the yeasts ability to grow on glucose rich media, while over expression of ScTAL1 inhibited the yeast cell’s ability to grow on glucose as a carbon source. In general over expression of PsTAL1 dramatically improved xylose uptake, cell growth and ethanol production. Under aerobic conditions, the specific ethanol productivity of strains with PsTAL1 over expressed was increased by 70% from the base strain and a 100% increase in the specific growth rate of the cells on xylose media. (Graphic?) The best strain created by Watanabe et al was found to produce 5.94 g ethanol per liter with a yield of 0.43 g ethanol per g of total consumed sugars which was 84% of the theoretical yield. Fermentation Activity (from Dombek) Under optimal conditions S. cerevisiae can produce ethanol at a rate of 50 mmol ethanol per hour per gram of cell protein. During batch fermentation this optimal rate is only maintained for a brief period followed by a rapid decline as ethanol accumulates in the broth. Molecular oxygen and magnesium have been identified as critical factors for maintaining high fermentative rates, but do not completely prevent the rate decrease. (Graphic?) Accumulation of ethanol is known to reduce membrane permeability and can increase hydrogen ion flux into the yeast cells. This increased proton flux has been proposed as the cause in the ethanol-induced reduction of fermentation rates. However, replacement of the fermentation broth containing ethanol does not immediately restore the fermentative activity. It was found that ethanol concentrations below 12% (vol/vol) do not denature the glycolytic enzymes or cause significant irreversible inhibition of the fermentative activity. Reactor Design and System Train of fed batch reactors Aerobic/Anaerobic? Separator to recycle yeast cells and send product ethanol on to distillation sequence Ethanol Recovery and Purification (from Lu and Aden) Ethanol vaporizes at 78°C and water vaporizes at 100°C. If distilled an azeotropic mixture of ethanol and water will form the distillate (95.6% ethanol to 4.4% water). Since the ethanol-water mixture is not ideal a multi-stage process must be implemented. Typically the fermentation mixture will be fed to a beer column. Carbon dioxide leaves out the top with water coming out the bottom and ethanol is removed from a side-draw. This ethanol is fed to a rectifying column, the product of which is the azeotropic mixture. This mixture cannot be further purified using distillation. The effluent is therefore fed to a dehydration unit. Typically a molecular sieve adsorption unit can be used. The zeolites in the molecular sieve unit selectively adsorb water from vapor mixtures, further purifying the ethanol. The energy input of the distillation/dehydration unit must be minimized to make the process economically feasible. Pressure swing adsorption can be implemented to remove the water from the azeotropic mixture, this gives a further optimization consideration for the process. Resources 1. Aden, A., M. Ruth, K. Ibsen, J. Jechura, K. Neeves, J. Sheehan, B. Wallace, L. Montague, A. Slayton, and J. Lukas. Lignocellulosic Biomass to Ethanol Process Design and Economics Utilizing Co-Current Dilute Acid Prehydrolysis and Enzymatic Hydrolysis for Corn Stover. National Renewable Energy Laboratory, 2002. Print. 2. Dombek, K. M., and L. O. Ingram. "Ethanol Production during Batch Fermentation with Saccharomyces cerevisiae: Changes in Glycolytic Enzymes and Internal pH." Applied and Environmental Microbiology 53.6 (1987): 1286-291. Department of Microbiology and Cell Science, University of Florida, Gainesville. Web. 6 Dec. 2009. 3. Jin, Yong-Su, Hal Alper, Yea-Tyng Yang, and Gregory Stephanopoulos. "Improvement of Xylose Uptake and Ethanol Production in Recombinant Saccharomyces cerevisiae through an Inverse Metabolic Engineering Approach." Applied and Environmental Microbiology 71.21 (2005): 8249-256. Department of Chemical Engineering, Massachusetts Institute of Technology. Web. 4 Dec. 2009. 4. Lu, Yulin, and Nathan S. Mosier. "Current Technologies for Fuel Ethanol Production from Lignocellulosic Plant Biomass." Genetic Improvement of Bioenergy Crops. Ed. Wilfred Vermerris. Gainesville, FL: Springer, 2008. 161-77. Print. 5. Watanabe, Seiya, Ahmed Abu Saleh, Seung Pil Pack, Narayana Annaluru, Tsutomu Kodaki, and Keisuke Makino. "Ethanol Production from xylose by recombinant Saccharomyces cerevisiae expressing protein-engineered NADH-preferring xylose reductase from Pichia stipitis." Microbiology 153 (2007): 3044-054. Kyoto University. Web. 6 Dec. 2009.