AOS-HCI_CombinedResearchPaper_Draft1.3 - AOS-HCI
advertisement

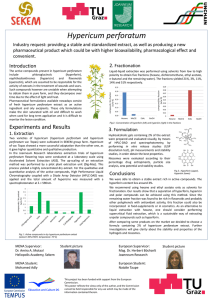
Investigation of the Effects of St John’s Wort on Cancer and Drosophila AOS-HCI Combined Research Paper 2011 Arturo Neo Yong Yao (4S1-02) Daniel Tan Han Jie (4S2-06) Ashley Ferguson (AOS) Caitlin Morris (AOS) Hwa Chong Institution (High School) Ms Sandra Tan Abstract Norepinephrine has been found to play a significant role in the progression of cancer by promoting focal adhesion kinase which is a protein that helps facilitate cell life. Cancer cells have also shown to have increased glycolysis for higher rate of production for ATP. Hypericum perforatum has been found to inhibit the reuptake of norepinephrine in cells, which should then slow the rate of cancer progression. It has also shown to be able to inhibit or denature certain enzymes that are involved in the process of glycolysis, hence slowing down the rate of which the cells are able to proliferate. The purpose of our research is to develop a method to slow the growth and metastasis of a mouse mammary cell line and that of a robust Drosophila cell line using the herb extract Hypericum perforatum. An extract of its main active ingredient, hypericin, was also tested on the mouse mammary cell line. The mouse mammary cell line CRL 2539 was cultured and two concentrations of a glycerol based liquid Hypericum perforatum extract (0.4%, 0.8%) as well as an extract of hypericin. (0.001%) were applied to the cells. The concentration of hypericin applied was based on typical amounts of hypericin found in a Hypericum perforatum plant. To gather data, the cells’ growth was measured using microscope cell counts on intervals of 24 to 48 hours until they reached confluency. The robust Drosophila cell line was also cultured similarly and the same glycerol based liquid Hypericum perforatum extract (2.5%, 5.0%, 12.5%, 50%) was applied to the cells. Cell counts were done using a haemocytometer daily at 24 hour intervals. Each experimental trial was run alongside a control trial in order to compare the growth of each. In addition, Hypericum perforatum extract had also been tested on live Drosophila melanogaster flies to observe if any immune-related proteins are affected by the extract. The flies were fed varying concentrations of the extract and their protein profiles were observed via SDS-PAGE. So far, the hypericin trials have yielded inconclusive results but the Hypericum perforatum extract has been found to slow the cancer’s growth in both the mouse mammary cell line as well as the robust Drosophila cell line. After SDS-PAGE, 4 protein bands observed in the control, which were missing in the experimental set ups, were then sent for MALDI-TOF. 2 of these proteins had been identified to be enzymes essential for glycolysis. In addition, samples of the mouse mammary cells’ media have been taken at the end of each trial so that the norepinephrine levels can be measured in an ELISA kit. Likewise, an ELISA can also be used to observe how the physiopathology of the robust Drosophila cells are affected, especially in the areas of protein kinases and glycolytic enzymes. Trials are continuing to evaluate the effect of hypericin on cell growth and measuring the degree to which varying concentrations of Hypercium perforatum extract slow down the growth of cancer cells. Introduction Hypericum perforatum is a yellow flowering plant found in various locations around the world including West Asia, Europe, and North Africa. This herb is often used in depression treatment. Other areas where Hypericum perforatum is applicable include treating diseases, disorders, minor burns, and injuries (Linde, 2009). Hypericum perforatum has been tested to treat depression in adults and attention deficit hyperactive disorder (ADHD) in children (Weber et al., 2008). Cancer prevention research is one of the main fields that the therapeutic effect of Hypericum perforatum can be investigated because it has been found to inhibit the reuptake of norepinephrine, dopamine, and serotonin, which play a role in the development and spread of cancer. Research has shown that the most significant components of Hypericum perforatum in terms of treating disease are hypercerin and hyperforin (Linde, 2009). One of the older but still practiced methods of cancer treatment involving hypericin is photodynamic therapy (PDT). PDT requires a photosensitizing agent (photosensitizer) and the visible light of a wavelength which correlates with the absorption spectrum of the drug. Alone the light and photosensitizer have no therapeutic effect, but when combined produce cytotoxic products which trigger irreversible tumor destruction and cell damage. Hypericin is probably nature’s stongest naturally photosensitizer so its use in PDT to induce cell apoptosis and necrosis is immense (Anastasia Karloti et al. 2010). Out of 36 species of Hypericum, 27 held hypericin, the most common being Hypericum perforatum (Agostinis, Merlevede, Vantieghem, & Witte, 2002). Cancer describes an immense group of related diseases. Each cancer case is different in terms of how fast the cells grow and its set of genetics changes, which is why cancer treatment is so difficult (Emory University, 2009). More specifically, cancer is “any malignant growth or tumor caused by abnormal and uncontrolled cell division; it may spread to other parts of the body through the lymphatic system or the blood stream” (Princeton University, 2010). Breast cancer is “..cancer that affects the breast”. Most cases of breast cancer occur in women although men have been found to develop it as well (Health Information Publications, 2010). The Drosophila melanogaster is commonly known as the fruit fly or vinegar fly. According to Lawrence Reiter et al. (2001) 714 distinct human disease genes (77% of disease genes searched) match 548 unique Drosophila sequences. Of the 548 Drosophila genes related to human disease genes, 153 are associated with known mutant alleles and 56 more are tagged by P-element insertions in or near the gene. Cross-genomic analysis with significant sequence matches can be carried out to prove the effects of drugs on diseases. In recent studies, researchers have been examining the effect of stress on the progression of cancer. The body responds to stress by releasing catecholamines such as norepinephrine and epinephrine. These catecholamines prevent the anoikis (programmed cell death) of cancer tumor cells by increasing focal adhesion kinase. Focal adhesion kinase, when activated, is a protein that promotes cell life. In cancer research when different factors are looked at in isolation, stress has been found to play a main role in tumor cell proliferation. This is because the cancer cells reuptake the catecholamines released by the body furthering the cells’ ability to live (Sood et al., 2010). Norepinephrine is one of the major catecholamines that plays a role in developing and spreading cancer. There are many norepinephrine receptors on organs and in the blood stream, and in the brain, which, when activated, may be used as message systems to spread cancer because of the intracellular processes taking place in the organs or blood stream (Fitzgerald, 2008). One field of research that has been focusing on a way to prevent the cancer cell reuptake of the catecholamines is complementary and alternative medicine (CAM). One of the main herbs that have been looked into is for its anticancer abilities is Hypericum perforatum (Sood et al. 2010). Glycolysis has also proven to be a new target for therapy in cancer. (Gatenby et. al. 2007) According to Rui-hua Xu et. al. (2005), the Warburg Effect has shown that cancer cells frequently exhibit increased glycolysis and depend largely on this metabolic pathway for generation of ATP to meet their energy needs. Under physiologic conditions, generation of ATP through oxidative phosphorylation in the mitochondria is an efficient and preferred metabolic process, which produces far more ATP molecules from a given amount of glucose compared with glycolysis. However, when the ability of cells to generate ATP through mitochondrial oxidative phosphorylation is compromised, cells are able to adapt alternative metabolic pathways, such as increasing glycolytic activity, to maintain their energy supply. Mitochondrial respiratory function can be negatively affected by multiple factors, including mutations in mitochon- drial DNA (mtDNA), malfunction of the electron transport chain, aberrant expression of enzymes involved in energy metabolism, and insufficient oxygen available in the cellular micro-environment. It is known that mtDNA contains a displacement loop, and the coding gene sequence for 13 important protein components of the mitochondrial respiratory complexes without introns. Mutations in mtDNA are likely to cause alterations of the encoded protein and compromise the respiratory chain function. Thus, the frequent mtDNA mutations observed in a variety of human cancers are thought to contribute to respiratory malfunction in cancer cells. The constant generation of reactive oxygen species within the mitochondria and the increased free radical stress in cancer cells may cause further damage to both mtDNA and the electron transport chain, thus amplifying respiratory malfunctions and dependency on glycolysis. Hypoxia is another important factor that contributes to the Warburg effect. The fast growth of cancer cells and rapid expansion of the tumor mass usually outpace new vascular generation, resulting in an insufficient blood supply to certain area of the tumor tissues. Such a hypoxic environment within the tumor mass limits the availability of oxygen for use in mitochondrial respiration and synthesis of ATP and forces the cancer cells to up-regulate the glycolytic pathway as the main route of energy production. The ability of oxygen to regulate glucose metabolism is know as the Pasteur effect and is mediated through several pathways involving various kinases. In this case, the increased glycolytic activity in cancer cells is not necessarily due to intrinsic mitochondrial defects but is induced by the tumor microenvironment through a series of metabolic adaptation processes, including preferentially increased expression of enzymes required for glycolysis. Also, according to Gatenby et. al.(2007), the upregulation of glycolysis in cancer cells results in increased glucose consumption, which leads to micro-environment acidosis. This allows for the cells to develop phenotypes which make them resistant to acid-induced toxicity, giving them a powerful growth advantage which ultimately leads to uncontrolled proliferation. However, therapies that involve glycolytic inhibitors have yet to yield any results in a human trials and further research is still being carried out at the present moment. The immune related proteins found in the Drosophila melanogaster are being further investigated in our experiments. Rationale and Objectives This project explores the possibility of using Hypericum perforatum as a treatment for cancer or a supplement for cancer therapy because while it has been most prominently used in the treatment of depression. About 1,596,670 new cancer cases are expected to be diagnosed in 2011 in the United States of America (Cancer Facts & Figures 2011) of which about 571,950 are expected to die of cancer. This numbers are astonishing and while survival rate has increased from 50% in 1975 – 1977 to 68% in 1999 – 2006, the number of people contracting cancer has been increasing as well. Hence we have the following objectives for our project: For the team in Hwa Chong, we aim: 1. To determine which proteins (if any) are affected by Hypericum perforatum 2. To determine the effects of Hypericum perforatum extract on a Drosophila cell line For our partners in Academy of Science (Virginia): 1. To determine the effects of Hypericum perforatum extract and hypericin on a mouse mammary cell line 2. To determine whether hypericin is the main active chemical constituent that gives Hypericum perforatum its medicinal properties This project would thus allow us to compare the effects of the Hypericum perforatum on the Drosophila melanogaster and the mouse mammary cell line, both of which are able to make cross-genetic comparisons to human diseases. Also, it allows us to compare the effects of the extract on a vertebrate (mouse mammary) and an invertebrate (robust Drosophila) cell line. Materials and Methodology Rate of Proliferation of Mouse Mammary Cell Line (AOS) Mouse models of human breast cancer are used widely to study molecular pathways of tumorigenesis. The overexpression of oncogenes known to be involved in human breast cancer, such as c-Myc, HER2/Neu, and Ccnd1, with tissue-specific promoters, induces tumors in the mammary gland of transgenic mice. Likewise, the inactivation of tumor suppressor genes via introduction of viral oncogenes or by conditional knockout using CreLox technology canpromote tumorigenesis also contributes to cancer. (Cristina Montagna et al. 2003). The procedure for our preliminary test for cancer cell binding is summarised in Figure 1. Culture 6 flasks (25cc) CRL 2539 Mouse Mammary Cells with 10,000 cells each Hypericum perforatum extract (0.4% & 0.8%) and Hypericin were added to the culture medium and grown. 3 other flasks were kept as controls Cells were counted every 24 hours through the use of a microscope A haemocytometer was used to count the cells after reaching confluency 1 ml of sample is frozen after each trial so that an ELISA kit measuring norepinephrine levels can be run in the future Fig. 1: Procedure for Rate of Proliferation of Mouse Mammary Cell Line (AOS) Materials: - Liquid Hypericum perforatum extract - Hypericin extract - CRL 2539 Mouse Mammary Cell Line - Cell Culture Medium (10% Fetal Bovine Serum) Independent Variable(s) - Constant Variables Number of mouse - Temperature mammary cells - Growth Medium Used or Hypericin) - Volume of extract used Concentration of extract - Type of cell line Extract added in medium (Hypericum perforatum - Dependent Variable(s) - used Rate of Proliferation of CRL1963 Robust Drosophila Cell Line (HCI) The Drosophila genes have a 77% match with the human diseases genes and through the investigation of the effects of the Hypericum perforatum extract on the robust Drosophila cell line, we would be able to gauge the effects it would have on human cancer cells. A summary of the experimental procedure is found in Figure 2. Sub culture 2 flasks of Drosophila cells into 5 flasks (25cc) Apply varying concentrations of Hypericum perforatum extract (2.5% - 50%) to the cell culture medium Cell counts at 24 hour intervals with the aid of a haemocytometer Check the rate of growth of the cells by plotting a graph showing the cell count against time & comparing it across concentrations Fig. 2: Procedure for Proliferation of Robust Drosophila Cell Line (HCI) Materials: - CRL1963 Robust Drosophila Cell Line - Hypericum perforatum Extract (2.5%, 5.0%, 12.5%, 50.0%) - Cell Culture Medium - Sterile Water (for control) Independent Variable(s) - Concentrations of Dependent Variable(s) - Hypericum perforatum Constant Variables Number of robust - Temperature Drosophila cells - Growth Medium Used - Volume of extract Extract added - Duration of incubation - Age and type of cell line Effects on Immune-Related Proteins of the Drosophila melanogaster (HCI) There are 3 phases to this experiment: Preparing Drosophila Samples SDS-PAGE MALDI-TOF Fig. 3: Outline of experiments for tests on Drosophila Melanogaster (HCI) Phase 1 involves preparing the Drosophila melanogaster by feeding them the Hypericum perforatum extract that was commercially purchased. Materials: - Wild Type Drosophila melanogaster Flies - Hypericum perforatum Extract - Sterile Saline - Sterile Water Independent Variable(s) - Concentrations of Dependent Variable(s) - Immune related protein Hypericum perforatum which strengthens the Extract Applied immune system of Constant Variables - Volume of extract added - Drosophila Number of Drosophila melanogaster - Duration of exposure This was done so with the following procedure: Varying concentrations of Hypericum perforatum were coated on the inner surface of the falcon tube (Control contains only sterile water Flies are transferred from original containers using chlorofoam Centrifuge the tubes at 10oC, 7000rpm for 10 minutes before washing the Hypericum perforatum extract using sterile saline Flies are placed in the coated falcon tubes for 24h before extraction Flies are crushed in 200µl of sterile saine and put on ice to ensure all bodily fluids are realsed into the solution Fig. 4: Procedure for preparing the Drosophila fly samples Phase 2 of the experiments would be Sodium Dodecyl Sulfate Poly-Acrylamide Gel Electrophoresis (SDS-PAGE). It is a technique used to separate proteins according to their electrophoretic ability in order for us to identify the unique protein profiles of the flies if they are any apparent. As suggested by the name of the procedure, a gel electrophoresis is run with the solutions prepared from the previous phase in order for us to determine whether any immune-related proteins of the fly were affected by the introduction of Hypericum perforatum extract. Materials: - Acrylamide Gel (5% Stacking Gel, 10% Running Gel) - Loading Dye - Protein Ladder for Electrophoresis - Solution of crushed Drosophila melanogaster flies Independent Variable(s) - Dependent Variable(s) - Solution of crushed Protein Profiles of each Constant Variables - sample Drosophila (placed in Concentration of Acrylamide Gel - varying concentrations Volume of solution added into each well Hypericum Perforatum Extract The samples were first prepared by mixing 2 units (10 µl) of the crushed Drosophila solution with 1 unit (5 µl) of loading dye in an eppendorf tube before placing them in a hot water bath (50oC) for 10 minutes, following which the standard SDS-PAGE procedure was carried out as follows: Load first well with 10µl of protein ladder, subsequent wells to be filled with prepared Drosophila solutions Set the electrophoresis kit to 25mA and allow the gel to run until the protein column runs to the end of the gel Stain the gel using Coomassie Blue & if any significant proteins are identified, they will be sent for MALDI-TOF Fig. 5: Procedure for SDS-PAGE M ctrl 50% 37.5% 25% 12.5% 2.5% 2% 1.5% 1% 1 2 1 3 1 4 1 Fig. 6: SDS-PAGE Gel showing the 4 samples of unknown proteins to be tested Phase 3 is the experimental procedure known as Matrix Assisted Laser Desorption/Ionization – Time-Of-Flight Mass Spectrometry (MALDI-TOF). This procedure allows for the analysis of biomolecules or large organic molecules. A co-precipitate of a UVlight absorbing matrix and a biomolecule is irradiated by a nanosecond laser pulse. Most of the laser energy is absorbed by the matrix, which prevents unwanted fragmentation of the peptides. The ionized peptides are accelerated in an electric field and enter the flight tube. During the flight in this tube, different peptides are separated according to their mass to charge ratio and reach the detector at different times. In this way each molecule yields a distinct signal. The intensity of the signal is directly related to the abundance of that specific peptide. The experiments were carried out in a laboratory located in Australia. Rate of Proliferation of CRL1963 Robust Drosophila Cell Line in a High Glucose Content Environment (HCI) In order to test whether Hypericum perforatum slows down the proliferation of the cells by retarding the rate of glycolysis by the cells, flasks with glucose added to the cell culture medium were tested. Materials - CRL1963 Robust Drosophila Cell Line - Hypericum perforatum Extract (50%) - 10% Glucose Solution - Cell Culture Medium - Sterile Water Independent Variable(s) - Presence of Hypericum perforatum extract - Presence of glucose Dependent Variable(s) - Constant Variables Number of robust - Temperature Drosophila cells - Growth Medium Used - Volume of extract/glucose added solution - Age and type of cell line A procedure similar to that shown in Figure 2 was used. We have tested the rate of proliferation of cells in high glucose content media in both the presence and the absence of Hypericum perforatum, as well as having a control set with only sterile water and another with only Hypericum perforatum added. Triplicates were conducted and cell counts are collected every 24 hours. Results & Discussion Introduction of Hypericum perforatum Extract slows down the rate of proliferation of Mouse Mammary Cell Line; Hypericin does not encompass the full effect of the Hypericum perforatum (AOS) Control 0h 24h 96h 144h Flask 1 10000 22919 347311 4900000 Flask 2 10000 29971 386097 4300000 Flask 3 10000 24682 281492 4550000 0h 24h 96h 144h 168h 264h Flask 1 10000 26445 42900 91088 145154 4800000 Flask 2 10000 24682 72871 124585 229190 4050000 Flask 3 10000 18218 74046 135751 257398 4350000 0h 48h 72h 120h Flask 1 10000 23507 23507 270000 Flask 2 10000 29971 29971 280000 Flask 3 10000 28208 28208 250000 0.4% Hypericum perforatum 0.001% Hypericin Tables 1.1 – 1.3: Showing the number of cells counted in the flasks at fixed time intervals After data of cell populations in the flasks were collected, graphs comparing the control and experimental trials at the time were made to compare how the cells were growing. Experimental Hypericum perforatum (0.4%) Growth vs. Conrol Growth Number of Cells 5000000 Experimental 0.4% Hypericum Perforatum Data Control Data 4000000 3000000 2000000 1000000 0 0 100 200 Time (hrs) Graph 1: Graph comparing 0.4% Hypericum perforatum Data and Control Data Number of Cells Experimental Hypericin (0.001%) Growth vs. Control Growth 900000 800000 700000 600000 500000 400000 300000 200000 100000 0 Control Data Experimental Hypericin Data 0 50 100 Time (hrs) Graph 2: Graph comparing 0.001% Hypericin Data and Control Data Graphs 1 and 2 show the number of cells that were in each of the three control or experimental flasks at certain times. After certain times (around 96 hours for the 0.4% Hypericum perforatum and around 72 hours for 0.001% hypericin) it is clear that the experimental cells are growing slower than the control cells. The difference between the cell populations between the experimental and control flasks increase as time goes on. Control Experimental Mean 25857.33 23115 Variance 13468732 18762499 Observations 3 3 Hypothesized Mean Difference 0 P(T<=t) two-tail 0.4499 Table 2.1: t-Test: Two-Sample at 24 Hours 0.4% Hypericum perforatum Control Mean Experimental 338300 63272.3333 2.8E+09 311619130.3 Observations 3 3 Hypothesized Mean Difference 0 Variance P(T<=t) two-tail 0.0134 Table 2.2: t-Test: Two-Sample at 96 Hours 0.4% Hypericum perforatum Control Mean Experimental 4583333 117141.3333 9.08E+10 540252022.3 Observations 3 3 Hypothesized Mean Difference 0 Variance P(T<=t) two-tail 0.0015 Table 2.3: t-Test: Two-Sample at 144 Hours 0.4% Hypericum perforatum Control Mean Variance Experimental 24682 27228.67 3108169 11165144 Observations 3 3 Hypothesized Mean Difference 0 P(T<=t) two-tail 0.32735 Table 2.4: t-Test: Two-Sample at 48 hours 0.001% Hypericin Control Experimental Mean 86974.67 22918.67 Variance 4.86E+08 4144225 Observations 3 3 Hypothesized Mean Difference 0 P(T<=t) two-tail 0.037581 Table 2.5: t-Test: Two-Sample at 72 hours 0.001% Hypericin Control Mean Variance Experimental 810000 266666.7 6.3E+09 2.33E+08 Observations 3 3 Hypothesized Mean Difference 0 P(T<=t) two-tail 0.007296 Table 2.6: t-Test: Two-Sample at 120 hours 0.001% Hypericin To analyze the data, t-tests were run to compare the mean number of cells at various times between the control and experimental flasks. The purpose of this test is to determine if there is a statistically significant difference between the cell populations at the set times, shown in the resulting p-value. P-values of less than 0.05 demonstrate statistically significant data at 95% confidence when used in comparing biology related data. As shown in tables 2.2 and 2.3, p-values of less than 0.05 were observed for the control compared to the Hypericum perforatum (0.4%) at times 96hrs (p-value 0.013) and 144hrs (p-value 0.0015). As shown in table 2.1, at 24hrs the p-value wasn’t less than 0.05 (p-value 0.45). As shown in tables 2.5 and 2.6, p-values of less than 0.05 were also observed for the control compared to the hypericin (0.001%) at times 72 hrs (p-value 0.037) and 120 hrs (p-value 0.007). As shown in table 2.4, 48 hours the p-value wasn’t less than 0.05 (p-value .016). Graph 3.1: Normal Distribution Comparing the Growth of 0.4% Hypericum perforatum and Control Cells at time 24 Hours Graph 3.2: Normal Distribution Comparing the Growth of 0.4% Hypericum perforatum and Control Cells at time 96 Hours Graph 3.3: Normal Distribution Comparing the Growth of 0.4% Hypericum perforatum and Control Cells at time 144 Hours Graph 3.4: Normal Distribution Comparing the Growth of 0.001% Hypericin and Control Cells at time 48 Hours Graph 3.5: Normal Distribution Comparing the Growth of 0.001% Hypericin and Control Cells at time 72 Hours Graph 3.6: Normal Distribution Comparing the Growth of 0.001% Hypericin and Control Cells at time 72 Hours The normal distribution curves for the Hypericum perforatum vs. control at 96hrs (graph 3.2) and 144hrs (graph 3.3) as well as hypericin vs. control at 72hrs (graph 3.5) and 120hrs (graph 3.6) provide a visual demonstration of the statistically significant difference between the experimental and control means. Because neither of the graphs had overlapping of either data set, the mean values of the control and experimental flasks at times 96hrs and 144hrs for the Hypericum perforatum and at times 72 hrs and 120 hrs for the hypericin were found to be statistically different. The normal distribution curves for the Hypericum perforatum vs control 24 hrs (graph 3.1) and the hypericin vs control at 48 hrs (graph 3.4) showed two data set overlapping demonstrating that there wasn’t a statistically significant difference between the experimental and control means at these times. Introduction of Hypericum perforatum Extract slows down the rate of proliferation of CRL1963 Robust Drosophila Cell Line (HCI) 24 hours 48 hours 72 hours Control 13267 10967 113700 2.50% 10133 12600 103275 5% 16267 15900 124800 12.50% 12167 19267 73050 50% 18467 19567 41775 Table 2.1: Comparison of cell numbers of control setup and setups whereby varying concentrations of Hypericum perforatum were added (newer cells set 1) 24 hours 48 hours 72 hours 96 hours Control 13050 14550 21825 12750 2.50% 13050 10800 20025 16425 5% 13050 18150 28725 17550 12.50% 13050 17100 24450 10500 50% 13050 11325 8850 13125 Table 2.2: Comparison of cell numbers of control setup and setups whereby varying concentrations of Hypericum perforatum were added (newer cells set 2) 96 hours 120 hours 144 hours Control 83100 73050 33375 2.50% 83100 92700 72000 5% 83100 94950 44025 12.50% 83100 87225 58125 50% 83100 90975 38850 Table 2.3: Comparison of cell numbers of control setup and setups whereby varying concentrations of Hypericum perforatum were added (older cells) Graph 4.1: Comparison of cell numbers of control setup and setups whereby varying concentrations of Hypericum perforatum were added (newer cells) Graph 4.2: Comparison of cell numbers of control setup and setups whereby varying concentrations of Hypericum perforatum were added (older cells) Tables 2.1 to 2.3 show the daily cell counts of our robust Drosophila cell line. These cells were subcultured and counted from a period of 0h – 72h. As can be observed, the peak of growth occurs between 48h to 72h and the introduction of Hypericm perforatum has resulted in a general slow down in the proliferation of the robust Drosophila cell line. This effect was most significant when 50% Hypericum perforatum extract was added to the cell culture medium as seen from graph 4.1. However, when the extract was tested on the older set of cells (cells grown from 96h – 144h), the Hypericum perforatum had a different effect on the proliferation of the cells. The cell numbers had started decreasing rapidly in the control flask but those flasks containing Hypericum perforatum extract showed a decrease in cell death rate as can be observed from graph 4.2. Observed Expected Chi-square value 2.50% 61650 64706.25 2588250 5% 76762.5 72262.5 1445250 12.50% 48750 58256.25 466050 50% 25312.5 46537.5 93075 Table 3.0: Chi-square values of the results of the newer cells To analyze the data, chi-square tests were run to compare the mean number of cells at 72h between the control and experimental flasks for the cells grown from 0h to 96h. The purpose of this test is to determine if there is a statistically significant difference between the cell populations after the introduction of Hypericum perforatum and reaching peak growth rate. A table (Table 3.0) showing the calculations, proving that the results at 48h are statistically significant. However, the behavior of the cells during the growth cycle from 0h – 144h suggests that Hypericum perforatum is only suitable to be used for treatment of cancer during the early stages rather than the late stages as it slows down rate of proliferation early on and slows down rate of cell death later on. Consumption of Hypericum perforatum extract by the Drosophila melanogaster results in the inhibition of glycolytic enzymes in the Drosophila (HCI) As shown from Figure 6, there were 4 protein bands that were present in the flies of the control setup when compared to those of the flies that have been fed the Hypericum perforatum for a period of 24h. The same bands were also observed to be missing when samples were tested with a low concentration of 0.5% Hypericum perforatum extract even though the effects were less obvious (Figure 7). This shows that Hypericum perforatum is dosage dependent at both high and low concentrations. 0.5% ctrl M Fig. 7: SDS-PAGE Gel showing how at 0.5% Hypericum Perforatum still shows an inhibition of the same 4 protein bands The 4 bands of protein were cut out from the SDS-PAGE gel and sent for MALDITOF for identification. The results that were obtained from the analysis were as follows: Sample Most-Highly Matched Protein Sequence Sequence Coverage B4IS91 1 (unknown 27% function) 2 Enolase 10% B3LX89 3 (unknown 26% function) Fructose4 bisphosphate aldolase 31% Table 4.0: Table showing the results obtained from the 4 unknown protein bands after MALDI-TOF Proteins 1 and 3 have no functions whatsoever and are possibly contaminants that were consumed by the fly. Proteins 2 and 4 have been identified to be Enolase and Fructose Bisphosphate Aldolase respectively. Both these enzymes have been found to be enzymes required to facilitate the process of glycolysis. (Figure 8) Fig. 8: Flow chart showing glycolysis and the enzymes facilitating the process (Source: http://en.wikipedia.org/wiki/File:Glycolysis.jpg) Glycolysis is the metabolic pathway that converts glucose into pyruvate. The free energy released in this process is used to form the high-energy compounds ATP (adenosine triphosphate) and NADH (reduced nicotinamide adenine dinucleotide). The absence of the 2 enzymes, Enolase and Fructose Bisphosphate Aldolase, could lead to a slow down or even stop the process of glycolysis entirely. With a slowed rate of glycolysis, the cancer cells will be unable to maintain their growth advantage and the rate of proliferation will also start to decrease. In order to prove this hypothesis, we tested the effects of adding glucose to the cell culture medium of the robust Drosophila cell line. Addition of Hypericum perforatum and glucose to the cell culture medium causes slow down in rate of proliferation of robust Drosophlia cells (HCI) A B C control 10462.5 9112.5 15412.5 glucose 13275 15075 15300 1237.5 10462.5 3262.5 glucose+ Hypericum perforatum Table 5.0: Comparison of cell numbers 24 hours after the start of experimentation Graph 5.0: Comparison of cell numbers between the setup with glucose, the setup with both glucose and Hypericum perforatum and the control setup From table 5.0 and graph 5.0, we are able to see that even though it has only been a period of 24 hours, the addition of glucose has significantly increased the proliferation of the robust Drosophila cells. However, with the addition of 50% Hypericum perforatum extract, the growth rate has shown to slow down and this has been proven statistically significant as shown by a p-value of 0.0398 on the t-test. While the results might not be conclusive at 24h, the experiment shows much promise as Hypericum perforatum at high and regular dosages might be able to counter the growth of cancer cells in humans if the Warburg effect is significant enough in cancer cell growth. Further experiments will be carried out to test the effectiveness of the Hypericum perforatum extract on robust Drosophila cells grown in a highly glucose-concentrated environment. Conclusion When Hypericum perforatum and hypericin were added to the mouse mammary cell line, the results had shown that cancer cells when exposed to the extract will grow at a slower rate compared to the control cells. It was concluded with 95% confidence that Hypericm perforatum and hypericin extracts did significantly slow the growth of the mouse mammary cancer cells after times of 96hrs for the Hypericum perforatum extract and 72hrs for the hypericin extract. With time, both extracts decreased cell growth at greater rates as shown in the decreasing p-values over time (0.013 to 0.0015 from times 96hrs to 144hrs for the Hypericum perforatum extract and 0.037 to 0.007 from times 72hrs to 129hrs for the hypericin extract). Hypericum perforatum has also shown to be able to slow down the growth rate of the robust Drosophila cells when added in high concentrations. This effect is most obvious during the 48h – 72h time period and it has also been proven to be statistically significant by the chi-squared test. Following which, the effect of the Hypericum perforatum was not so ideal when it slowed down the rate of cell death as compared to the control setup. Showing that Hypericum perforatum is only suitable to be used for early cancer treatment. One possible way that the Hypericum perforatum is affecting the proliferation of the cells is by disturbing the glycolytic pathway of the cells as it denatures or inhibits the production of 2 enzymes essential for glycolysis. Cancer cells have shown increased glycolysis in order to produce more ATP and with Hypericum perforatum affecting this pathway, the lack of energy could result in a higher rate of cell death or slow down in the rate of proliferation of these cells. Future Work ELISA stands for enzyme-linked immunosorbent assay. The purpose of which is to determine if a particular protein is present in a sample and if so, how much of it is present. There are two main variations on this method: one can determine how much antibody is in a sample, or determine how much protein is bound by an antibody. The distinction is whether one is trying to quantify an antibody or another protein. For the AOS side, an ELISA test will be run to determine the amount of norepinephrine in each sample, representative of the amount of norepinephrine secreted by the cells in each flask. The experimental samples should contain greater amounts of norepinephrine than the control samples because Hypericum perforatum is thought to inhibit the reuptake of norepinephrine in cells. Each sample of increasing concentrations of Hypericum perforatum and hypericin should contain greater amounts of norepinephrine. A SDS-PAGE gel can also be run to determine the proteins in the mouse mammary cell line that have been inhibited or denatured by the Hypericum perforatum and further tests by ELISA can determine how the concentration of the extract affects the amount of proteins denatured or inhibited by the Hypericum perforatum. For the Hwa Chong side, we aim to find out more about the physiopathology of the Drosophila cell growth in the areas of protein kinases and the cell glycolysis pathway through the use of an ELISA. Through this, we will be able to understand how the Hypericum perforatum is causing a change in the growth rate of the cells and hence deduce how such an effect can also be observed in human cells. Also, both sides would continue to experiment on varying concentrations to determine how the dosage and concentration of Hypericum perforatum extract would affect the growth rates of both cell lines and ultimately determine what is the optimum for dosage and concentration for usage as a possible clinical drug for trial. References Heck, D., Torshina, N., Zhang, J. (2007). Catalytic therapy of cancer with ascorbate and extracts of medicinal herbs. eCam, 7(2), 203-212. Doi: 10.1093/ecam/nem159 Hulka, B., Moorman, P. (2008). Reprint of breast cancer: hormones and other risk factors. Maturitas, 61(1), 203-213. Doi: 10.1016/j.maturitas.2008.11.016 Leuner, K., Kazanski, V., Muller, M., Essin, K., Henke, B., Gollasch, M., Harteneck, C., Muller, W., (2007). Hyperforin—a key constituent of St. John’s Wort specifically activates TRPC6 channels. The FASEB Journal, 21, 4101-4111. Doi: 10.1096/fj.078110com Linde, K. (2009). St. John’s Wort- an overview. Karger, 16(3), 146-155. Doi:10.1159/000209290 Martinez-Poveda, B., Medina, M., Quesada, A. (2005). Hyperforin, a bio-active compound of St. John’s Wort, is a new inhibitor of angiogenesis targeting several key steps of the process. International Journal of Cancer, 117 (5), 775-780. Doi: 10.1002/ijc.21246 Pomerantz, M., Ahmadiyeh, N., Jia, Li., Herman, P., Verzi, M., Doddapaneni, H., Beckwith, C., Chan, J., Hills, A., Davis, M., Yao, K., Kehoe, S., Lenz, H., Haiman, C., Yan, C., Henderson, B., Frenkel, B., Barrentina, J., Bass, A., Tabernero, J., Baselga, J., Regan, M., Manak, J., Shivdasani, R., Coetzee, G., Freedman, M. (2009). The 8q24 cancer risk variant rs6983267 shows long-range interaction with MYC in colorectal cancer. Nature, 41, 882-884. Doi: 10.1038/ng.403. Rigel, D. (2008). Cutaneous ultraviolet exposure and its relationship to the development of skin cancer. Journal of the American Academy of Dermatology, 58(5), S129-S132. Doi: 10.1016/j.jaad.2007.04.034 Rothenburg, M. (n.d.). Corning Life Sciences: an introduction to cell culture: an introductory lesson on how to culture and handle mammalian cells. Retrieved December 10,2010 from http://mmsrvolbintellor.com/corning/111209/corning.html?go=Launchvideo Sood, A., , Halder, J., Nick, A., Stone, R., Hu, W., Carroll, A., Spannuth, W., Deavers, M., Allen, J., Han, L., Kamat, A., Shahzad, M., McIntyre, B., Diaz-Montero, C., Jennings, N., Lin, Y., Merritt, W., DeGeest, K., Vivas-Mejia, P., Lopez-Berestein, G., Schaller, M., Cole, S., & Lutgendorf, S. (2010). Adrenergic modulation of focal adhesion kinase protects human ovarian cancer cells from anoikis. The Journal of Clinical Investigation, 120(5), 1515-1523. doi:10.1172/JCI40802 The Trustees of Princeton University. (2010). Retrieved October 12, 2010 from: http://wordnetweb.princeton.edu/perl/webwn?s=cancer Wang, Y., Broderick, P., Webb, E., Wu, X., Vijayakrishnan, J., Matakidou, A., Qureshi, M., Dong, Q., Gu, X., Chen, W., Spitz, M., Eisen, T., Amos, C., Houlstoun, R. (2008). Common 5p15.33 and 6p21.33 varients influence lung cancer risk. Nature, 40, 14071409. Doi: 10.1038/ng.273 Weber, W., Stoep, A., McCarty, R., Weiss, N., Biederman, J., McClellan, J. (2008). A randomized placebo controlled test for attention deficit hyperactivity disorder in children and adolescents. Journal of the American Medical Association, 299(22), 2633-2641. Doi:10.1001/jama.299.22.2633 Karloti, A., & Billia, A. R. (2010). Hypericins as potential leads for new therapeutics. Int. J. Mol. Sci. 2010, 11, 562-594. Reither, L. T., Potocki, L., Chien, S., Gribskov, M., & Bier, E. (2001). A systematic analysis of human disease-associated gene sequences in drosophila melanogaster. Genome Res. 2001, 11, 1114-1125. Gatenby, R. A., & Gillies, R. J. (2007). Glycolysis in cancer: a potential target for therapy. Int J Biochem Cell Biol. 2007, 39(7-8), 1358-1366. Xu, R., Pelicano, H., Zhou, Y., Carew, J. S., Feng, L., Bhalla, K. N., Keating, M. J. & Huang, P. (2005). Inhibition of glycolysis in cancer cells: a novel strategy to overcome drug resistance associated with mitochondrial respiratory defect and hypoxia. Cancer Res 2005, 65, 612-621. Gatenby, R. A., & Gillies, R. J. (2004). Why do cancers have high aerobic glycolysis?. Nature Reviews Cancer, 4, 891-899. Pressman, AH. (1998). St. john's wort - the miracle medicine. New York: The Philip Lief Group, Inc. American Cancer Society. Cancer Facts & Figures 2011. Atlanta: American Cancer Society; 2011. Vogel, C., Hager, C. & Bastians, H. (2007) Mechanisms of mitotic cell death induced by chemotherapy-mediated g2 checkpoint abrogation. Cancer Res,67, 339-345 St. John's Wort Side Effects, Interactions and Warnings . (2010). Personal health zone. Retrieved November 4, 2010, from http://www.personalhealthzone.com/stjohnswort.htm Acknowledgements We would like to thank the following people, without whom our project would not have been successful: - Ms Sandra Tan, our HCI mentor, for her unwavering support and excellent mentorship of our project - Ms Jackie Curley, our AOS mentor, for her guidance and support throughout the project - Mdm Lim CF, lab manager of the Science Laboratories at the Science Research Centre, for her most helpful assistance and guidance when carrying out our experiments - Hwa Chong Institution, for kindly allowing us the opportunity to collaborate with Academy of Science, Virginia on this project as well as allowing us to conduct our experiments in the science laboratories - Mr George Wolfe and Mr Duke Writer, for their generous help and advice during the preparatory phase and finalization of the project - Any other person who has helped us along the way in one way or another