03_tsetse_disease_transmission
advertisement

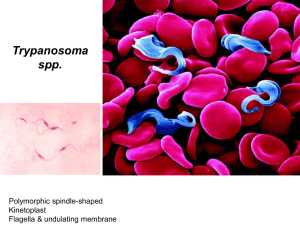
Arthropod vectors Tsetse flies Tsetse flies Author: Dr Reginald De Deken Licensed under a Creative Commons Attribution license. DISEASE TRANSMISSION The transmission of trypanosomes by the tsetse fly occurs cyclically. This means that the vector, when he feeds on an infected host, is not immediately infectious but that the pathogen during “the extrinsic incubation period” has to pass through a developmental cycle inside the vector. During this period the trypanosome undergoes substantial morphological and metabolic changes. The developmental cycle of Trypanosoma vivax is restricted to the mouthparts and lasts 5 to14 days. Trypanosoma congolense develops in the midgut and the proboscis and its development takes generally around 12 - 18 days. Trypanosoma brucei species have the most complicated cycle with trypanosomes migrating from the midgut to the salivary glands of the tsetse fly. The cycle takes between 17 and 45 days. The basic reproductive rate of an infection is the mean number of secondary cases caused by an infected individual into a population with no pre-existing immunity to the disease and in the absence of control interventions. It is often denoted R0. However, what is often claimed to be R0 is, in fact, often simply the assessment of a threshold (the disease spreading or not spreading), not of the average number of secondary infections. This is particularly true in case of a vector-borne disease because calculations to assess the R0 of a vector-borne disease are very complex (since both the vector and the host are responsible for the secondary cases and since several vectors and hosts may be present) and the exact value of several contributing parameters are often unknown. Thresholds will merely demonstrate whether a disease will die out (if R0 < 1) or will be able to spread in a population (if R0 > 1). The important parameters governing the spread of a disease may be demonstrated by the formula (based on the work of Macdonald (1957) and Smith et al. (2007)) to assess the basic reproductive rate of a vector-borne infection: m a2 b c pn R0 = --------------r g Where: m = rate of vectors to hosts (amongst others this will depend on the space utilization of the host and alternative hosts in relation to tsetse mobility and distribution). a = feeding rate of the vector on the host (depends on the feeding rate “f” at that moment, and also on the feeding preference “q” of tsetse flies for a particular host) so a = fq 1|Page Arthropod vectors Tsetse flies b = probability that a host may become infected once bitten by an infected vector c = the probability that the vector becomes infected from a bite on an infected host p = daily survival rate of the vector, so p = e-g and the death rate g = -ln(p) g = death rate of the vector n = extrinsic incubation period (number of days before the vector becomes infectious) r = the rate with which the host cures from the infection. Regarding m: The rate of vectors to hosts (m) may decrease as a result of: vector control, disappearance or fragmentation of tsetse habitat, or a massive immigration of cattle in an area (e.g. temporarily available pasture land) The rate of vectors to hosts (m) may increase as a result of: - livestock movements (e.g. cattle moving into game reserves), - particularly beneficial climatic conditions for tsetse (at the end of the rainy season the tsetse fly population density is generally at its top because of an increased reproduction in the previous months and an increased longevity at that time), - tsetse flies seeking refuge in forest galleries during the hot dry season resulting in a temporarily increased transmission risk for cattle at watering points. Notwithstanding the tsetse population is not at its highest density at that time high trypanosomosis incidences may be observed, - sudden immigrations of tsetse flies (these may be particularly high in case of tsetse control operations when the control area is not isolated making the control effort often unsustainable). Regarding a: This important parameter depends on the number of feeds taken by the vector per unit of time, which will increase when the climate becomes dry and hot, but it depends also on the feeding preference (see the biology section). Furthermore, it was demonstrated that upon colonization of the tsetse salivary glands with Trypanosoma brucei, the anti-haemostatic activity of the saliva changes resulting in a prolonged probing/feeding time, which may result in an increase of interrupted feeding and thus contribute to an increased parasite transmission (Van den Abbeele et al., 2010). On the other hand, increased probing activity was observed in T. congolense infected G. morsitans flies, which is believed to be caused by physical interference of the parasite with phagoreceptors in the proboscis in combination with a reduced diameter of the tsetse labrum due to the presence of parasite rosettes (Livesey et al., 1980). Regarding b: This may depend on the “trypanotolerance” or “susceptibility” to trypanosome infection of the host. Trypanotolerant animals are more difficult to infect (smaller value of “b”) and once infected they may clear the infection more rapidly, resulting in a larger value for the parameter “r”. The parameter b will also be reduced when prophylactic trypanocidal drugs are used. 2|Page Arthropod vectors Tsetse flies Regarding c: This will depend on the vectorial competence (ability to become infected) and vectorial capacity (the ease with which the development cycle in the vector is completed) of the vector. In most tsetse flies taking up trypanosomes, these protozoa will be killed by the combined action of proteases, lectins, immunopeptides and oxidative stress (Lehane, 2004; MacLeod, 2007). Young flies infect themselves more easily with trypanosomes than old ones. However, it was shown that even older flies can show a similar vectorial capacity in case the vector undergoes a stress of some kind which affects the fly’s immune gene expression (Akoda, 2009). The morsitans group of tsetse flies has a higher infection rate than those of the palpalis group (except for T.vivax in West Africa). The fusca group is generally a poor vector for the T.brucei group of trypanosomes but experiments showed it to be a good vector for T.vivax and T.congolense. Regarding p and g: these depend on the longevity of the vector, which will be influenced by environmental conditions (climate, host availability, tsetse density dependent mortality) and possibly by local tsetse control operations. Regarding n: to become infectious the longevity of an infected tsetse fly must be superior to the extrinsic incubation period of the trypanosome. This period varies according to the trypanosome species and the environmental conditions (e.g. when temperatures are rising, the pathogen will develop more rapidly in the vector). It is obvious that the formula above is too simple to give a correct reflection of what happens in the field; for example some trypanosomes can infect several livestock species (e.g. T.congolense infects ruminants as well as pigs or dogs) and thus the vector can become infectious by feeding on any one of those infected hosts. Feeding rate will differ depending on the particular host, its density and the tsetse fly species involved. Available multimedia On the website of TDR animations can be found on the life-cycle of a Trypanosoma brucei sp. In the human host http://www.who.int/tdr/publications/documents/t-brucei-humanhost.swf and the complex development cycle of T. brucei in the tsetse fly http://www.who.int/tdr/publications/documents/t-brucei-tsetse-fly.swf 3|Page