Microsoft Word
advertisement

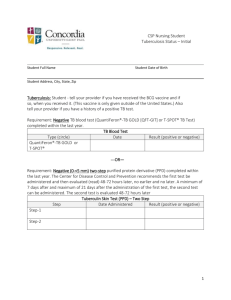
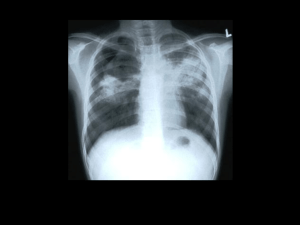
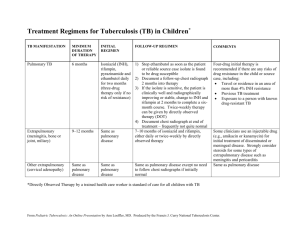
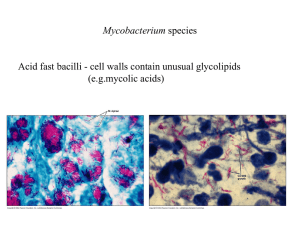
Abstract Tuberculosis remains a scourge of humanity with a high mortality rate and killing a couple of million people every year. Tuberculosis is caused by Mycobacterium tuberculosis which is an incredibly successful pathogen. One third of the world’s population has been infected with M. tuberculosis. Although The World Health Organization has implemented several programs to eradicate this dreaded disease, the number of new cases of tuberculosis being reported every year is increasing at an alarming rate. M. tuberculosis is transmitted in the form of aerosolized droplets generated by the cough of an infected individual. These ingested bacilli are engulfed by alveolar macrophages. Mycobacterium not only survives the microbicidal techniques of the macrophages and evades the immune response but also replicates during the early stages of infection. The site of infection is characterized by the formation of granulomatous lesion or a tubercle. In spite of the highly effective immune response against M. tuberculosis, the bacillus persists within the tubercle, in a latent state, for a prolonged time interval and is called latent tuberculosis. The reactivation of latent tuberculosis sometimes occurs in these infected individuals due to compromised immune system. The mechanism of latency and its subsequent reactivation is not fully characterized and is a subject of active research. The elucidation of these mechanisms will pave the way for new drugs and vaccines for the treatment of tuberculosis. The heat shock proteins have been implicated in the pathogenesis of M. tuberculosis. The GroE and DnaK families are the major heat shock chaperone machineries in M. tuberculosis. The genome of M. tuberculosis has two paralogs of groEL, groEL1 (Rv3417c) and groEL2 (Rv0440). These v GroELs have been previously characterized to be present in uncanonical oligomerisation states. Sequence analyses of more than 200 sequences have revealed that both the M. tuberculosis GroELs have significantly diverged in their functions during the course of evolution. Unlike E.coli GroEL, M. tuberculosis GroEL1 has a histidine rich C- terminal tail which has been demonstrated to be defective in biofilm formation in Mycobacterium smegmatis. M. tuberculosis GroEL1 is a nonessential gene and mutant lacking groEL-1 is defective in cytokine dependent granulomatous response. These intriguing observations emphasize the necessity to further characterize the heat shock machinery of M. tuberculosis. The work embodied in this thesis aims to understand the structural and functional aspects of M. tuberculosis transcription factors and stress proteins, with primary focus being on the Hsp60 family of chaperones. This thesis demonstrates the DNA binding property of un-canonical chaperone GroEL1 for the first time. The role of GroEL1 as a nucleoid associated protein is also elucidated. Furthermore, experiments have been performed to identify the genome wide binding sites of GroEL1. The putative transcriptional repressor protein of M. tuberculosis GroE system, HrcA has also been characterized. The in depth characterization of these proteins have been described systematically in six chapters. Chapter 1 of the thesis introduces the topic of research, reviews the various aspects of the mode of action of M. tuberculosis, the role of heat shock proteins in the pathogenesis and the un- canonical nature of GroE system and its regulation. This chapter describes the recent advancements in the field and also defines the main objectives of the study. The various techniques implemented during the course of this study are described in Chapter 2. This study utilizes the recombinant DNA vi techniques and a spectrum of protein purification procedures. Several DNA protein interaction protocols (Electrophoretic mobility shift assay, Chromatin immuno-precipitation followed by Southern blot hybridization) are employed to address the objective of the study has been described in detail. The procedure for isolation of nucleoid from M. tuberculosis has been elaborately described. The regulation of stress response has an important role in pathogenesis of M. tuberculosis. Chapter 3 focuses on the biochemical and biophysical characterization of HrcA (Heat regulation at CIRCE), a putative transcriptional regulator protein of the GroE operon. ANS fluorescence studies demonstrates that the protein to have exposed hydrophobic residues and Circular Dichroism studies characterize HrcA to be a predominantly α- helical protein. These studies further demonstrate that GroEL1 and HrcA do not interact to each other in the experimental conditions. The probability of cross regulation of both the GroE and DnaK heat shock operons by HspR has been experimentally disproved. The most striking observation of this chapter is the observation of DNA binding ability of GroEL1 to CIRCE instead of HrcA under identical experimental conditions. This chapter reveals the uncanonical nature of stress response in M. tuberculosis, a feature which is further studied in the following chapters. The DNA binding ability of M. tuberculosis GroEL1 and its probable physiological functions have been characterized in Chapter 4. M. tuberculosis genome sequence shows the remarkable absence of many nucleoid associated proteins. The nucleoids of M. tuberculosis have been purified and interestingly discovered that a novel nucleoid associated protein in M. tuberculosis is a sequence homolog of GroEL chaperonin. This chapter reports that M. tuberculosis GroEL1 binds nucleic acid substrates without vii sequence specificity and plays a role in the condensation of DNA in nucleoid formation. Chromatin immunoprecipitation followed by Southern blot hybridization and immunofluorescence imaging of M. tuberculosis confirms the in vivo GroEL-1 DNA interaction in M. tuberculosis. This chapter therefore reveals that GroEL1 has evolved to be associated with the nucleoids. The southern blot hybridization and immuno-fluorescence microscopy reported in Chapter 4 suggested that GroEL1 has a preference for binding some regions of the M. tuberculosis genome. To address this observation, Chromatin immunoprecipitation followed by microarray analysis (ChIP-chip) was performed to identify all the binding sites of GroEL1 across the entire genome. The ChIP-chip experiment and its experimental validations have been reported in Chapter 5. Surprisingly, genome-wide binding analyses revealed preferential enrichment in the genes encoding PE-PGRS class of family by ChIP-chip assay. A consensus motif for the GroEL1-DNA interaction was also identified from the ChIP-chip interaction. The implications of the present research work on the understanding of the heat shock proteins of M. tuberculosis have been described in Chapter 6. This is the first report to describe the DNA binding property of M. tuberculosis GroEL1. The results presented in this thesis characterize this novel property of the unique chaperonin GroEL1 of M. tuberculosis in detail. The present results suggest several directions and opportunities for future research work. viii