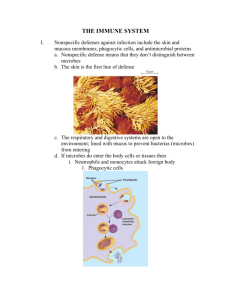
Line 3 2014 - Franklin College
advertisement

Immune System 3rd LineSpecific Immunity(Line3)-characterized by specificity, inducibility, clonality, memory, and tolerance. The Lymphocyte response in depth: #1-4 Overview: I. Development of Dual lymphocyte systemStem cells diverge into 2 populations. B lymphocytes-bone marrow (born in and educated the same place-home school) which are involved in humoral immunity (antibody production) and Tlymphocytes (born in bone marrow and then move out to thymus to become educated). Tinvolved in CMI (cell mediated immunity). Education involves becoming committed to responding to one antigen. Gene shuffling-build the T and B receptors on these cells during the education process. Also, lymphocytes that respond to self antigens are eliminated(tolerance). Resulting cells are now immunocompetent (primed to react should they encounter their partner antigen). Once educated, lymphocytes migrate to regional lymphoid tissues. Here they provide a lifelong source of immune cells. B lymphocytes secrete protective antibodies. T lymphocytes help B-cells and also kill host cells that harbor pathogens or have become cancerous. II. Entrance and Processing of Antigens and Clonal selectionWhen foreign cells enter fluids of host they encounter blood, lymph and RES. Here they encounter a battery of cells that screen, trap, and eliminate. Macrophages/Dendritic cells usually first to react. Ingest, process at the site of infection. Then they migrate (it takes about a day) to the lymph tissue (especially nodes) and present the antigens to the B and T lymphocytes. This presentation (usually along with additional signals) activates a specific lymphocyte. Usually activation of a B lymphocyte requires assistance of T-helper cells. III. Activation of Lymphocytes and Clonal expansion Challenge by ag stimulates B and T cells to differentiate and proliferate. The response is so powerful that it could kill normal body cells if the process were not properly regulated. Activation requires multiple signals in the right sequence (binding ag and lymphokines that bind to receptors on cell's surfaces for example). Nuclear launch analogy. If activated, lymphocytes forms clone, some of which are memory cells, and others effector cells (B-plasma cells for example) IV. Activated B lymphocytes and their products-Humoral Immunity B cells found in spleen, MALT, lymph nodes, and a small % in the blood. Active progeny of B cells are effector plasma cells (no surface receptors, they secrete proteins). Synthesize and secrete antibodies into fluids (tissue fluids, lymph, and blood-humoral). Antibodies mark antigens for destruction or neutralization. Memory cells also produced. V. Activated T lymphocytes and CMIT cell types and responses are extremely varied. T cells- found in the blood (70-80% of the circulating lymphocytes are T cells) and lymph nodes. T-cells only respond to AGs that are part of cell surfaces. Sensitization by antigen causes T cell to differentiate into either: 1) T helper-2) suppressor cells 3) cytotoxic cells. T cells produce lymphokines but no secreted antibodies. Essential preliminary concepts to understand specific immunity (3rd line of defense) 1. cell surface molecules (MHC,CD, B and T cell receptors) 2. Cytokines 3. antigens 4. antibodies. 1. Cell surfaces- molecules (MHC antigens, CD receptors, and B and T cell receptors). Several different types of receptors on each cell. Major function of receptors are 1) perceive and attach to non-self (antigens) 2) help in recognition of self 3) receive and transmit chemical messages among other cells of the system 4) aid in cellular development. Major cells with receptors are lymphocytes and macrophages/dendritic cells #5 A. MHC molecules MHC surface molecules (antigens) are glycoproteins on all cells except rbc. Also known as HLA antigens (first discovered on leukocytes). MHC molecules play a key role in recognition of foreign vs. self and are key players in the process of antigen presentation (dendritic, macro, b-cell). The MHC holds and positions the antigen fragments for presentation to other immune cells. #6 2 routes of antigen processing . Intracellular pathogens class I, extracellular class II. Class I-On all cells except rbc. First recognized because they regulate acceptance or rejection of tissue grafts. Each person inherits a unique set of class I genes. Closer people related, more likely it is that their MHC profile is similar. Individual differences exist however. You have enough unique Class I antigens that your tissues are likely to be antigenic to others. Each nucleated cell has a fragment (epitope) of every cell polypeptide it produces (including viral and cancer peptides) displayed on its surface as part of an MHC I so it can be shown to other immune cells. For our purposes- important in elimination of virus infected/cancerous cells. #7 Class II-code for immune regulatory receptors. Found only cell surface of APC (macrophages, dendritic cells, B, and T-helper cells). If an AG can’t be displayed with a MHC II, it can’t trigger an immune response. Thus, MHC’s determine which Ag fragments might trigger an immune response. When a pathogen is phagocytized, pieces of it are loaded onto an MHC II receptor and shipped to the cell surface. This cell can then present ag to other immune cells. T cells are MHC restricted-Class I, T-killer, Class II, t-helper. #8 B. CD molecules-Lymphocytes have additional receptors that define cell type. Various lymphocytes often just referred to as CD-8 (for example) C. B and T Cell Receptors: #9 B cell receptor structure- Ig-G model. 2 heavy and 2 light chains. Light chain is kappa or lambda. Heavy is one of five types (A-E see below). Hinge region between the light and heavy chains (allows swiveling of Fab region to fit angle of antigen better). Hypervariable region (model of ab-this is the hands). Igg has a valence of 2, allows cross linking. B cell receptors are unique (compared to T cells) in that they have the same basic structure (minus the membrane anchoring piece in the B-cell) as the antibody molecule that B cells secrete. This is not the case for T-cells because they secrete no antibody. Each receptor recognizes on one specific antigen. 50,000 copies of same bcr/cell #10 T-cell receptor structure (similar to B except one heavy and one light chain)- Binding of ag to tcr/mhc complex activates a cascade that triggers cell development and cytokine production. TCR complex has two components, one that recognizes self (MHC) and one that recognizes foreign (ag bind site). The receptor only recognizes an epitope if it associated with an MHC receptor (restricted). T cells can’t bind free ag, only antigens that are prat of cell surfaces. How are the B and T cell receptors (and antibodies) made? How many foreign antigens will a person encounter in their life? Potentially, billions. Each antigen will need a cell with receptors for that specific antigen (1 ag/receptor). How many different types of immune cells are there? Possibilities? A. One (e very immune cell has receptors on its surface for every different antigen)? Nonot enough room on the cell surface for all of those different receptors (even one copy of each). B. There billions of different types of cells each with specificity for a different antigen? Yes-it has been shown that every B or T lymphocyte carries a unique receptor for a unique antigen. Generation of Receptor Diversity: So where do all the diverse cells with unique receptors come from? The progenitors of all B and T cells are stem cells. How do stem cells have enough genes to pass on to their offspring to code for all of this diversity of receptors? It is estimated that there are 1023 different types of B and t cells each with a unique receptor. Does that mean that stem cells have that many genes that code for receptor structure and that they pass these onto their offspring? No-there are only about 20-25,000 genes in the human genome. In reality, there are slightly more than 200 genes in stem cells that code for lymphocyte receptors. How do 200 genes give rise to 10 23 receptors? As part of their education process in bone marrow (B cells) and thymus (t-cells) the lymphocytes randomly recombine segments from 3 immunoglobulin regions in DNA (like shuffling a deck of cards) #11 Lymphocyte development involves shuffling blocks of genes (while B cells are developing in bone marrow or T-cells developing in Thymus). RSS sites (recombination signal sequence) are a key to this process. An enzyme called RAG (recombination activating gene protein) randomly combines one of each kind of various segments to form a BCR gene. Segregation, rearrangement and assembly results in 2 complete transcriptional units-one that codes for an H chain and one that codes for a light. This phase of development is antigen independent. #12 The result is 5 X 10 6 receptor combinations. Where does the remaining diversity come from to get to 10 23? (that one hundred billion trillion -10 X the stars in the universe) A. RAG randomly removes portions of D and J segments before joining them together B. Another enzyme randomly adds nucloetides to the heavy chain VDJ combination. C. B cell genes in the lymph node populations udergo rapid mutation in the V-regions D. As memory cells divide, they become increasingly adapted to better fit antigen during clonal selection #13 Isotype class switch – Once a cell has undergone gene shuffling, it always produces an antibody/receptor with the same antigen specificity. However, it can produce several classes of antibody with that same specificity (class switching) depending on circumstances. This involves associating a new Fc region (5) with the other gene products. The most common class switch is from IGM to IGG. Some lymphocytes then switch IGG to IGA. Making memory B- cellsMemory are B-cells that have already undergone switching. Long lived. B-cells that haven’t class switched produce suicide protein (apoptosis). After switching, cells produce an antisuicide protein. During their life time memory cells hyper mutate. #14,15 Once synthesized the IG is transported to cell surface and acts as a receptor. Now the cell is antigenically committed but it is still immature (naïve cell). It needs additional signals to give rise to effector and memory cells. Upon stimulation by appropriate antigen/cytokine, B cells form plasma and memory cells and plasma cells secrete antibody with same antigenic specificity as receptor. In order for this to happen, a minimum of one signal is required (ag binding). Usually a second signal is also required (lymphokines). # 16,17 Clonal DeletionSince T ( and B) cell receptors are generated randomly (in the thymus and bone marrow respectively), it’s inevitable that some cells will be formed that will react autoantigens. Body eliminates these through clonal deletion (in thymus and bone marrow). Exposure of these cells to autoantigens stimulates them to undergo apoptosis. If this doesn’t happen properly, you can have autoimmune disease. # 18 Clonal Selection- When then surviving non-self antigen binds to its assigned lymphocyte, it stimulates that cell to divide many times to form a clone of identical cells (these then form effector cells and memory cells). This process is called clonal selection. 2. Cytokines-soluble regulatory proteins that act as intracellular messengers Most immune cells won’t react to antigens without receiving the appropriate chemical signals (along with stimulation by an antigen). Many cytokines are redundant and form a complex cytokine network. #19 a. Interlukins signal among leukocytes. Designated in order of discovery (35 identified to date). b. Interferons-(gamma secreted by T-helpers activate macrophages). c. growth factors-stimulate leukocyte stem cells to divide. Body can control progression of immune response by limiting the production of growth factors. d. Tumor necrosis factor-Macrophages and T cells secrete to kill tumor cells and regulate the immune response and inflammation. e. chemokines-stimulate leukocytes to moves to site of infection (chemotaxis) #20-Selected immune cytokines 3. Antigens:(Antibody generating molecules). Antigenicity has special requirements. Size, shape, and complexity key elements. Molecules with complex composition (proteins, glycoproteins) are more immunogenic than large repeating polymers. MW must be > 1000 to stimulate abs. #21 Lymphocyte responds to a small piece of Ag molecule (ag determinant). Many complex molecules have many ag determinants (mosaic antigens). These stimulate many different types of lymphocytes (polyclonal response). Examples include components of bacterial cell walls, capsules, pili, flagella, toxins, and capsids of virus. #22 Haptens-small foreign molecules (drugs, metals, env. chems.) that are too small to elicit immune response (an example of an individual ag determinant). Immunogenic if conjugated to a larger carrier (serum proteins)-allergies. #23 Types of Ags-(many are from human sources)1) autoantigens (self)- may be hidden when immune system is developing. (eye). If later exposed, may stimulate abs (autoimmune). Basis of rheumatoid arthritis? 2) alloantigens-found on cells of some members of species but not others. Basis of blood grouping and MHC profiles. Tissue rejection. 3) Heterophile ags-same ag found in different species (cross reactions-false serological tests). Grp A strep and heart valve tissue (rheumatic fever). Homologous ag-that which induces the ab (strep), heterologous ag, other which react with the induced ab (heart valve)(cross reactions) 4) Allergens 5) Superantigens-Potent stimulators of T-cells. (enterotoxin of Staph). Cross link T cell receptors and trick T-cells to release massive amounts of cytokines. Damages blood vessels and cause multi organ failure. Basis of toxic shock syndrome. 6) T-dependent and T-independent Ags: A. T independent ag's are in the minority. They can directly stimulate B cells to secrete ab. Only 1 signal required for this . These are usually simple molecules with repeating units (CHO's). They activate B cells to to make IG-m only (no memory response). Why? No T-helper involved so no interlukins involved (second signal). Thus no class switching (IGM to IGG)because that is mediated by by IL-4. B. T-dependent-Majority of ags. These antigens can only activate B cells if a T-helper is involved. Remember, T-helper are MHC II restricted. They can only react with an antigen presented to them by an APC. This process involves internalization and presentation of ag by APC . Antigens stimulate antibodies. 4. Antibodies #24 AB structure- (just like B-cell receptor except secreted) Ig-G model. 2 heavy and 2 light chains. Light chain is kappa or lambda. heavy is one of five types (see below). Hinge region between the light and heavy chains (allows swiveling of Fab region to fit angle of antigen better). Hypervariable region (model of ab-this is the hands). Igg has a valence of 2, allows cross linking. Fc region-effector region. Fc binds to receptors on body's cells (granulocytes and agranulocytes). Effect antibody binding has on cell depends on type of cell (if it is a mast cell it may degranluate, if it is a macrophage it may stimulate phagocytosis). Fc region influences permeability of ab (crossing placenta, distribution of ab (is it secreted). It also may or may not fix complement (attach). Fc region determines class which antibody belongs to. Threats confronting the immune system are variable so its not surprising that there are several different classes of antibodies to respond to these varied threats. Class of Ab produced depends on the type of invading antigen, portal of entry, and the antibody function required (determined by the Fc region of the antibody). #25 Roles of ABs- (why make Abs handout)-react with ags and immobilize them, tags antigens for destruction (opsonins-kick me), neutralizes (fills surface receptors on virus and alters structures of toxins), stimulates cell destruction (turns on complement system), agglutinates (basis of many serological tests), killing by oxidation (antibodies catalyze reactions that produce substances that kill bacteria (H2o2, ozone), and antibody cellular toxicity (Fc regions bind to NK cells, these secrete perforin (forms channels in cell membrane) by which granzymes enter, these cause apoptosis). This is similar to opsonins in that ab’s coat the surface of pathogens. However, opsonins result in phagocytosis of the antibody coated pathogen rather than apoptosis. Some abs can cause disease (autoantibodies). #26 5 Classes of Abs (isotypes)-Type of AB produced depends on type of Ag, portal of entry, and antibody function required 1. M-(macro)-1st class secreted by B cells. Found mainly in blood. Most effective in fixing complement also an opsonin. Once it produces IG-M, a plasma cell (if it gets the right signal) may recombine its variable region with a new Fc region (class switching) to produce a new class of Ab. The most common class switch is to produce IG-G. This only happens in response to Tdependent antigens. 2. G-produce by activated memory cells. Makes up majority of class of proteins in blood serum called gamma globulins (80%). Longest lasting. Half-life is 23 days versus 4-5 for other classes. Neutralizes toxins, opsonin, fixes complement, antibody dependent cell toxicity, and only one that can cross the placenta. It also can easily leave the blood and enter extracellular space so it can bind to the pathogen before it enters the circulatory system. 3. A-2 forms-serum (12% of blood antibodies) and secretory. Secretory form prevents binding of ags to mucosal surfaces. Found in mucous secretions of salivary glands, intestine, nasal membranes, breast, lung, and urogenital tract. Secretory piece added by gland itself. Protects ab against digestive enzymes. Most important in local immunity to enteric and urogenital pathogens. Also secreted into milk. Total amount of secretory A produced is greater than of serum IG-G. 4. D-basically a BCR. Triggers B cell activation and involved in immune suppression. Not all mammals produce this. Role? 5. E- Fc region reacts with eosinophils, trigger release of cell damaging molecules onto the surface of cells (especially parasitic worms). Also binds to basophils and mast cells receptors to cause degranulation. Initiates inflammatory response, mediates allergies. In developed countries, IG-E more often associated with allergies than parasitic worms. Primary vs Secondary Immune response. #27 Primary response-ag is being concentrated in lymphoid tissues, processed by APC’s, and b synthesis finally begins. Anamenestic response yields 1000X amount of ab as primary. Faster because memory B cells don’t have to go through the early steps of activation and require fewer signals to form plasma cells. #28 Types of Immunity 1. Active-Immunological stimulus activates T and B lymphocytes-results in antibody release. Characterized by memory and is long lasting. Takes a long time to develop. Naturalbiological experience (subclinical can stimulate protection). Artificial-induced medically (vaccines and immune serum). Vaccines for 24 diseases. 2. Passive-receive antibodies produced by a donor-no memory, immediate protection, short lived. Natural-maternal antibodies-up to one year. Breast milk-intestinal protection not available otherwise. Artificial-horse serum, injected with toxins. Pooled gamma globulins (from recovering patients)-rabies, and hepatitis. CombinationsNotes-natural active-Many viral diseases cause life-long immunity. Other pathogens immunity that last a few months to years. Even a subclinical infection can stimulate active immunity. Part B-Line 3-Specific Immunity-Interplay of B/T lymphocytes and macrophages #29 2 Branches to Immune System-Humoral and Cell Mediated. Lots of overlap between two branches. T-helper cells are involved in both Humoral and CMI. #30 I. Cell Mediated Immunity Cell mediated- The body uses cell-mediated immune responses to fight intracellular pathogens such as viruses that have invaded body cells, as well as abnormal body cells such as cancer cells. CMI respond to intracellular pathogens by activating T-killers . T cells have a unique T-cell receptor and are MHC restricted (they can only see foreign antigen in the spatial context of the molecular architecture of cell surfaces). #31 Additionally T cells express CD proteins that define what type of cell they are (T-killers have CD8, T-helpers CD-4). CD molecules serve multiple roles including receptors, cell adhesion, and cell communication. CD 8 recognizes MHC I and CD4 MHC II The players in CMI: 1) Antigen presenting cells: present antigen to both TH-1 (Ag + Class II MHC) and Tc cells (Ag+ Class I MHC) 1) T-helpers (1)-CD4: T-helper most prevalent of all T cells- 65% of total. Hiv depress them (binds to CD-4 receptor). Help regulate activity Tc (and B cells) by providing necessary signals and growth factors. After stimulation by APC, TH-1 stimulates Tc cell via cytokine signaling 2) T-cytotoxic (CD 8). After activation by TH-1 and infected cell (Ag + Class I MHC on infected cell), Tc kills host cell by perforin/granzyme pathway. This process accounts for much of our immunity to foreign cells (fungi and protozoans), viral infections, and cancers (T deficient people more prone to cancer). It is also responsible for rejection of transplanted tissue. #32 Ist event in CMI is usually Tc activation. Steps involved in activation of cytotoxic T cells: Tc activation require 2 signals (activation in lymph nodes):TCR/mhc I/ag APC complex binding ad IL-2 produced by activated TH-1. 1) Antigen presentation by APC to Th-1 and Tc cells. Dendritic cells phago. pathogens at site of infection (skin for example) and then migrate to lymph nodes (it takes about a day to do this) T-helper cell in lymph nodes browses all APC’s in every lymph node every day. 2) TH-1 and Tc activation. Activated Tc cell moves to infected tissue and binds to cells that have ag on surface. Tc also produce lymphokines including MAF (macrophage activation factor) , MIF (migration inhibition factor) and, transfer factor (mobilizes other nonsensitized T cells to take on characteristics of Tc) 3) Tc Clonal expansion 4) Tc Self-stimulation #33 Activated Tc cells kill infected/cancerous cells via the The PerforinGranzyme Cytotoxic Pathway# 3435 In the perforin-granzyme pathway, cytotoxic T cells destroy their targeted cells by secreting perforins and granzymes, toxic protein molecules. Perforins perforate cytoplasmic membranes, and granzymes enter the target cell and activate capase enzymes which cause apoptosis. CMI must be regulated carefully regulated or else it could kill normal host cells (as opposed to virus infected/cancerous cells). Tc cells must receive signals in a precise order (combination lock) to be activated. T reg cells (CD-4/25)also control immune response by shutting down innate immunity (when the threat has been eliminated) and also repressing autoimmunity. To prevent autoimmunity, T cells require additional signals from an APC. If they do not receive these signals, they will not respond. # 36 II. Humoral ImmunityGoal is to produce AB that will help clear ag. B cell differentiation-bone marrow . Certain bone marrow sites harbor stromal cells. Huge cells that nurture lymphocytes and provide hormonal signals that influence the development of B cells. These signals stimulate cell division and gene shuffling that results in billions of different distinct B cells each with a unique receptor. These are released to regional lymphoid sites (lymph nodes, spleen, and GALT). 90% of B cells die within a few days. Reseeded by bone marrow. These cells have class I and II MHC receptors plus B cell receptor. However, they are not MHC restricted. Humoral response-antibodies produced by effector B cells (plasma cells) are directed against extracellular pathogens. Plasma cells are basically antibody secreting factories. An activated plasma cells has no BCR and can produce thousands of antibody molecules per second. It dies in a few days. # 37 Inducement of T-Independent Antibody Immunity T-independent antigens can directly stimulate B cell activation without needing the assistance of macrophages and T-helpers Because some large antigens have repeating epitopes that can be processed by B cells without the help of T cells, they are called T-independent antigens. The repeating subunits of Tindependent antigens allow extensive cross-linking between numerous BCRs on a B cell which activates transcriptions which stimulates the B cell to proliferate. Stimulated B-cells produce IGM but do not class switch to produce OGG. T-independent antibody immunity is relatively weak, disappears quickly, and induces little immunological memory. Children have stunted Tindep ag response. Pathogens that have T-ind ags are not eliminated. Example-Hemophilus influenza (capsule ags are t-indep) can cause meningitis if the child has not been vaccinated. # 38,39 Inducement of T-Dependent Antibody Immunity with Clonal Selection # 40,41 T-dependent antigens lack the numerous, repetitive epitopes and large size of T-independent antigens. Immunity against them requires the assistance of type 2 helper T (Th2) cells. These ags don’t cross link BCR (which is a normal B-cell activation signal) so these cells need help (signals) from other cells (T-helpers). If the Ag is T-depend, B cell activation requires 2 signals (Ag biding and cytokines) # 42,43 A T-dependent antibody immune response involves the following series of interactions among antigen-presenting cells, helper T cells, and B cells, which are mediated by cytokines: 1. Antigen presentation for Th activation and cloning. Activated T-helper produces membrane protein CD-40L which binds to the CD-40 on the B lymphocyte. This provides the second signal for AB production. 2. Differentiation of helper T cells into Th2 cells 3. Activation of B cell by the Th2 cell such that it proliferates rapidly and differentiates into either a memory B cell (retains its BCR and can last decades) or plasma cell (does not have a BCR and dies in a few days) Once B-lymphocyte activated, it produces clones of itself. These clones produce ab with a slightly higher specificity for ag because of hypermutation. Plasma cells die in a few days but ab’s can last a few weeks. Once plasma cells start secreting AB, they can class switch if they receive the proper signal. IGMIGG-GIGA (possibly). As the antibody immune response progresses, plasma cells that secrete antibody with a higher affinity for the epitope survive at a higher rate such that the antibody specificity becomes progressively better over the course of the immune response. Summary of T/B cell activation requirements: THo activated by APC+ag/MHC IITH1 and Th2 B lymph acitiv by TH-2 and ag/MHC II Tc-activated by TH1 and APC/ag/MHCI Memory B Cells and the Establishment of Immunological Memory A small percentage of the cells produced by B cell proliferation do not secrete antibodies but survive as memory B cells, long-lived cells (some last a lifetime) with BCRs complementary to the specific antigen that triggered their production. In a primary immune response, it can take days for relatively small amounts of antibodies to be produced. When an antigen is encountered a second time, the activation of memory cells in the secondary immune response ensures that the immune response is rapid and strong. Enhanced immune responses to subsequent exposures are called memory responses. Summaries- # 44-46 Additional Considerations: IR also affected by numerous factors1. Virus-AIDS 2. Sterols-tends to depress wbc. Stress produces cortisols. Cortisols used to suppress immune system in tissue transplants. Sterols also used to treat chronic inflammation. 3. Exercise-positive benefits on immune system. Produces endorphins and enkephalins (natural opiates). Also enhances immune activity. Also stimulates production of IL-I and If. 4. Mind IS connection. IL-2 also affects brain and nervous system. Nervous system produces ILHeavy enervation of lymphoid tissue and bone marrow (connection between mind and immunity). Modified 11-14