mice exhibit high levels of serum PPi and are hypophosphatemic
advertisement

Appendix Murine models of Vascular Calcifications Genetic mutations in humans and murine models have identified a variety of genes that inhibit VC processes that can be classified into three categories: modulation/transport of inorganic phosphate (Pi) metabolism; inorganic pyrophosphate (PPi) availability; and bone formation and homeostasis. Solute carrier family 20, member 1 and 2 are genes that encode the type III sodiumdependent phosphate transporters PiT1 and PiT2, which are predominantly expressed on vascular smooth muscle cells (VSMCs) and transport extracellular Pi into the cytoplasm to regulate phosphate homeostasis (1a). In vitro studies show VC is dependent on PiT1 (2a)2, and in vivo murine studies have identified that these transporters serve redundant roles in regulating Pi homeostasis (3a). Recently mutations in PiT2 have been linked to the brain-specific VC in the basal ganglia of patients with idiopathic basal ganglia calcification (IBGC) (4a). Inorganic pyrophosphate (PPi) is well-known potent endogenous inhibitor of calcium phosphate precipitation in tissues (5a). Tissue non-specific alkaline phosphatase (ALPL in humans, Akp2 in mice) hydrolyzes endogenous PPi and promotes ectopic mineralization. Akp2-/- mice exhibit high levels of serum PPi and are hypophosphatemic. Conversely, humans with generalized arterial calcification of infancy (GACI) mutations in the gene ectonucleotide pyrophosphatase/phosphodiesterase 1 (ENPP1) cannot metabolize extracellular ATP to AMP and PPi. The reduction in PPi results in drastic VC of the large arteries of GACI patients and ENPP1-/- murine models (6a). Breeding of the Akp2-/- and the ENPP1-/- mice rescues the phenotype (7). The tiptoe walking (ttw/ttw) mouse is a naturally occurring mutant line that develops dystrophic cardiac calcinosis and the genetic defected was found to occur in the ABCC6 gene (8a), the genetic cause of pseudoxanthoma elasticum (PXE) in humans (9a-11a), which can present clinically similar to GACI (12a). Though the substrate of ABCC6 remains to be identified, a recent study identified that ABCC6-/- mice have very low levels of plasma PPi, which suggests that like GACI, PXE is the result of insufficient circulating PPi (13). Similar to the models above, mice with mutations in the progressive ankylosis (Ank) gene develop ectopic calcification in the joint capsules and soft tissues that ultimately causes paralysis and death, which stems from a drastic decrease in extracellular PPi (14a). In addition to Pi/PPi homeostasis, murine models have identified protein factor can negatively modulate VC and perhaps contribute to VC in MMS. Klotho (KL) is a gene that regulates insulin sensitivity and vascular homeostasis, and KL-/- mice exhibit VC similar to that seen in the elderly (15a) and humans with mutations in KL develop tumoral calcinosis due to diminished FGF23 signaling (16a). Mothers against decapentaplegic homolog 6 (Smad6) is a transcription factor downstream of TGF- and BMP signaling expressed in the heart and blood vessels. Smad6-/mice develop medial VC and elevated blood pressure (17a), and families with congenital cardiovascular malformations (CVM) were found to harbor nonsynonymous variants in the Smad6 gene (18a). Matrix GLA protein (MGP) is produced by vascular smooth muscle cells and chrondrocytes and binds to minerals in the extracellular matrix. MGP-/- mice develop extensive calcification in the arteries and die due to vessel rupture, illustrating the potent inhibitory role MGP plays in preventing ectopic mineralization (19a). Keutel syndrome (KS) is an autosomal recessive human disease stemming from mutation in MGP, and these patients develop aberrant calcification in soft tissues (20a). Similar to PPi, osteopontin (OPN) is a circulating protein that inhibits the formation of hydroxyapatite (21a) and OPN-/- mice crossed to MGP-/- mice exhibit exacerbated medial calcification on the arteries (22a). Stemming from the above human and animal data, a few animal and clinical trials have been undertaken to identify the role of pyrophosphate and alkaline phosphatase inhibitors for the treatment of VCs. In vivo animal studies have showed that pyrophosphate injected into uremic rats prevents VC in these animals (23a). Bisphosphonates are structurally similar to PPi and inhibit both hydroxyapatite crystal formation and ALPL/Akp2 activity (24a). Proof-of-concept experiments used VSMCs from ENPP1-/- and ank/ank mice to show that inhibition of tissue nonspecific alkaline phosphatase is a potential method to impede and/or prevent VC (25). A clinical trail tested the effectiveness of bisphosphonates as treatment for GACI, and found that early administration of bisphosphonates to GACI patients diagnosed shortly after birth increased survival by slowing, and one case reversing, the progression of VC (26a, 27a). REFERENCES 1a. Giachelli CM. The emerging role of phosphate in vascular calcification. Kidney Int 2009;75:890-7 2a. Li X, Yang HY, Giachelli CM. Role of the sodium-dependent phosphate cotransporter, Pit-1, in vascular smooth muscle cell calcification. Circ Res 2006;98:905-12 3a. Crouthamel MH, Lau WL, Leaf EM, Chavkin NW, Wallingford MC, Peterson DF, Li X, Liu Y, Chin MT, Levi M, Giachelli CM. Sodium-dependent phosphate cotransporters and phosphate-induced calcification of vascular smooth muscle cells: redundant roles for PiT-1 and PiT-2. Arterioscler Thromb Vasc Biol 2013;33:2625-32 4a. Wang C, Li Y, Shi L, Ren J, Patti M, Wang T, de Oliveira JR, Sobrido MJ, Quintans B, Baquero M, Cui X, Zhang XY, Wang L, Xu H, Wang J, Yao J, Dai X, Liu J, Zhang L, Ma H, Gao Y, Ma X, Feng S, Liu M, Wang QK, Forster IC, Zhang X, Liu JY. Mutations in SLC20A2 link familial idiopathic basal ganglia calcification with phosphate homeostasis. Nature genetics 2012;44:254-6 5a. Fleisch H. Diphosphonates: history and mechanisms of action. Metab Bone Dis Relat Res 1981;3:279-87 6a. Okawa A, Nakamura I, Goto S, Moriya H, Nakamura Y, Ikegawa S. Mutation in Npps in a mouse model of ossification of the posterior longitudinal ligament of the spine. Nat Genet 1998;19:271-3 7a. Hessle L, Johnson KA, Anderson HC, Narisawa S, Sali A, Goding JW, Terkeltaub R, Millan JL. Tissue-nonspecific alkaline phosphatase and plasma cell membrane glycoprotein-1 are central antagonistic regulators of bone mineralization. Proc Natl Acad Sci U S A 2002;99:9445-9 8a. Meng H, Vera I, Che N, Wang X, Wang SS, Ingram-Drake L, Schadt EE, Drake TA, Lusis AJ. Identification of Abcc6 as the major causal gene for dystrophic cardiac calcification in mice through integrative genomics. Proc Natl Acad Sci U S A 2007;104(11):4530-5. 9a. Bergen AA, Plomp AS, Schuurman EJ, Terry S, Breuning M, Dauwerse H, Swart J, Kool M, van Soest S, Baas F, ten Brink JB, de Jong PT. Mutations in ABCC6 cause pseudoxanthoma elasticum. Nature genetics 2000;25:228-31 10a. Le Saux O, Urban Z, Tschuch C, Csiszar K, Bacchelli B, Quaglino D, Pasquali-Ronchetti I, Pope FM, Richards A, Terry S, Bercovitch L, de Paepe A, Boyd CD. Mutations in a gene encoding an ABC transporter cause pseudoxanthoma elasticum. Nat Genet 2000;25:223-7 11a. Ringpfeil F, Lebwohl MG, Christiano AM, Uitto J. Pseudoxanthoma elasticum: mutations in the MRP6 gene encoding a transmembrane ATP-binding cassette (ABC) transporter. Proceedings of the National Academy of Sciences of the United States of America 2000;97:6001-6 12a. Nitschke Y, Baujat G, Botschen U, Wittkampf T, du Moulin M, Stella J, Le Merrer M, Guest G, Lambot K, Tazarourte-Pinturier MF, Chassaing N, Roche O, Feenstra I, Loechner K, Deshpande C, Garber SJ, Chikarmane R, Steinmann B, Shahinyan T, Martorell L, Davies J, Smith WE, Kahler SG, McCulloch M, Wraige E, Loidi L, Hohne W, Martin L, Hadj-Rabia S, Terkeltaub R, Rutsch F. Generalized arterial calcification of infancy and pseudoxanthoma elasticum can be caused by mutations in either ENPP1 or ABCC6. Am J Hum Genet 2012;90:25-39 13a. Jansen RS, Kucukosmanoglu A, de Haas M, Sapthu S, Otero JA, Hegman IE, Bergen AA, Gorgels TG, Borst P, van de Wetering K. ABCC6 prevents ectopic mineralization seen in pseudoxanthoma elasticum by inducing cellular nucleotide release. Proc Natl Acad Sci U S A 2013;110:20206-11 14a. Ho AM, Johnson MD, Kingsley DM. Role of the mouse ank gene in control of tissue calcification and arthritis. Science 2000;289:265-70 15a. Kuro-o M, Matsumura Y, Aizawa H, Kawaguchi H, Suga T, Utsugi T, Ohyama Y, Kurabayashi M, Kaname T, Kume E, Iwasaki H, Iida A, Shiraki-Iida T, Nishikawa S, Nagai R, Nabeshima YI. Mutation of the mouse klotho gene leads to a syndrome resembling ageing. Nature 1997;390:45-51 16a. Ichikawa S, Imel EA, Kreiter ML, Yu X, Mackenzie DS, Sorenson AH, Goetz R, Mohammadi M, White KE, Econs MJ. A homozygous missense mutation in human KLOTHO causes severe tumoral calcinosis. J Clin Invest 2007;117:2684-91 17a. Galvin KM, Donovan MJ, Lynch CA, Meyer RI, Paul RJ, Lorenz JN, Fairchild-Huntress V, Dixon KL, Dunmore JH, Gimbrone MA, Jr., Falb D, Huszar D. A role for smad6 in development and homeostasis of the cardiovascular system. Nat Genet 2000;24:171-4 18a. Tan HL, Glen E, Topf A, Hall D, O'Sullivan JJ, Sneddon L, Wren C, Avery P, Lewis RJ, ten Dijke P, Arthur HM, Goodship JA, Keavney BD. Nonsynonymous variants in the SMAD6 gene predispose to congenital cardiovascular malformation. Hum Mutat 2012;33:720-7 19a. Luo G, Ducy P, McKee MD, Pinero GJ, Loyer E, Behringer RR, Karsenty G. Spontaneous calcification of arteries and cartilage in mice lacking matrix GLA protein. Nature 1997;386:78-81 20a. Munroe PB, Olgunturk RO, Fryns JP, Van Maldergem L, Ziereisen F, Yuksel B, Gardiner RM, Chung E. Mutations in the gene encoding the human matrix Gla protein cause Keutel syndrome. Nat Genet 1999;21(1):142-4. 21a. Steitz SA, Speer MY, McKee MD, Liaw L, Almeida M, Yang H, Giachelli CM. Osteopontin inhibits mineral deposition and promotes regression of ectopic calcification. Am J Pathol 2002;161:2035-46 22a. Speer MY, McKee MD, Guldberg RE, Liaw L, Yang HY, Tung E, Karsenty G, Giachelli CM. Inactivation of the osteopontin gene enhances vascular calcification of matrix Gla proteindeficient mice: evidence for osteopontin as an inducible inhibitor of vascular calcification in vivo. J Exp Med 2002;196:1047-55 23a. O'Neill WC, Lomashvili KA, Malluche HH, Faugere MC, Riser BL. Treatment with pyrophosphate inhibits uremic vascular calcification. Kidney Int 2011;79:512-7 24a. Fleisch H. Bisphosphonates--history and experimental basis. Bone 1987;8 Suppl 1:S23-8 25a. Narisawa S, Harmey D, Yadav MC, O'Neill WC, Hoylaerts MF, Millan JL. Novel inhibitors of alkaline phosphatase suppress vascular smooth muscle cell calcification. J Bone Miner Res 2007;22:1700-10 26a. Rutsch F, Boyer P, Nitschke Y, Ruf N, Lorenz-Depierieux B, Wittkampf T, WeissenPlenz G, Fischer RJ, Mughal Z, Gregory JW, Davies JH, Loirat C, Strom TM, Schnabel D, Nurnberg P, Terkeltaub R, Group GS. Hypophosphatemia, hyperphosphaturia, and bisphosphonate treatment are associated with survival beyond infancy in generalized arterial calcification of infancy. Circ Cardiovasc Genet 2008;1:133-40 27a. Ramjan KA, Roscioli T, Rutsch F, Sillence D, Munns CF. Generalized arterial calcification of infancy: treatment with bisphosphonates. Nat Clin Pract Endocrinol Metab 2009;5:167-72




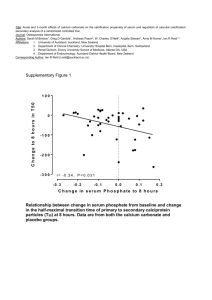
![Historical_politcal_background_(intro)[1]](http://s2.studylib.net/store/data/005222460_1-479b8dcb7799e13bea2e28f4fa4bf82a-300x300.png)



