Purkinje cell layer
advertisement

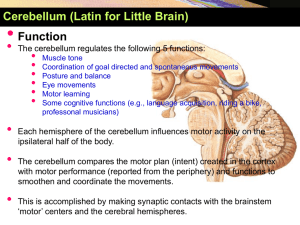
بسم هللا الرحمن الرحيم Central nervous system 1 Date : 20/2/2011 Anatomy lecture # 9 Done by: Ahmad H. Abu-Hanieh In the last lecture, we talked about the anatomy of the cerebellum. And we started discussing the pathways reaching it. - The cerebellum is mainly concerned with unconscious proprioception, and so, its main inputs are coming from the muscles and joints. (Remember that the main functions of the Cerebellum are to Coordinate between the agonist and antagonist, to Compare between the intended and actual movements, and to send Corrective signals to the cerebral cortex). - Afferent pathways of the cerebellum: Dorsal (posterior) Spinocerebellar tract o The afferents of this pathway arise –mainly but not exclusively- from the muscle spindle and golgi tendon organ of the lower limb and lower trunk. Beware that if a pathway’s name begins with “spino-“ , it doesn’t mean that it arises from the spinal cord, but rather from the peripheral receptors (muscle spindle, golgi tendon organ, touch and pressure receptors,….etc). o These afferents are Ia and II fibers (for muscle spindle). And Ib fibers (for golgi tendon organ). o The cell bodies of these afferents are located within the dorsal root (sensory) ganglion. o They enter the spinal cord, and run along the Gracile tract of the dorsal column of white matter. o Remember that the dorsal column consists of two parts: the Gracile tract (which brings sensations from the lower limb and lower trunk) and the Cuneate tract (which brings sensations from the upper limb and upper trunk) o When they reach the L2 spinal segment they synapse in a nucleus called dorsal nucleus of Clarke which is located in lamina VII of spinal segments C8-L2. o After synapsing within this nucleus, these 2nd order neurons ascend ipsilaterally forming this dorsal spinocerebellar tract. o They enter the cerebellum through the inferior cerebellar peduncle (ICP), and then they reach the medulla and from the medulla to the cortex. Cuneocerebellar tract: o It’s the upper limb counterpart of the dorsal spinocerebellar tract. So, it transmits sensory signals from muscle spindle (Ia and II) and from golgi tendon organ (Ib) of the upper limb and upper trunk. o In this pathway, the afferents enter the spinal cord and run along the cuneate tract of dorsal column, and then they synapse within the accessory cuneate nucleus (the counterpart of Clarke’s nucleus in lower limb) that’s located within the medulla. o After synapsing with the accessory cuneate nucleus, the 2nd order neurons also ascend ipsilaterally and form this cuneocerebellar tract. Ventral (anterior) Spinocerebellar tract: o Its inputs are exclusively Ib afferents from golgi tendon organ. o This pathway conveys information to the cerebellum about the on-going spinal reflexes. Mainly, those occurring in the lower limb (e.g. stretch reflex, inverse stretch reflex, walking reflex and part of it the withdrawal reflex,….) o As we already know, the spinal reflexes are accomplished mainly through excitatory and inhibitory interneurons. Now, to inform the cerebellum about the spinal reflexes going on, branches or collaterals of these interneurons project to the transmission cells (mother cells of this tract) of the ventral spinocerebellar tract, which are located within laminae V, VI, VII. o These fibers cross to the opposite side of the spinal cord (this is the first crossing in this pathway) and continue upwards reaching the medulla, then the pons, then at the level of the midbrain they enter through the superior cerebellar peduncle and go to the opposite side of the cerebellum (this is the second crossing). o Undergoing two crossings in this pathway, the effect will certainly be ipsilateral. (This confirms the last sentence mentioned in the previous sheet, that each cerebellar hemisphere controls muscles of its ipsilateral side of the body). o The counterpart of this tract in the upper limb is called Rostral Spinocerebellar tract, and it’s said to be very poorly developed in humans. (the doctor didn’t mention anything else about this tract and even considered it not worth mentioning :S ) The cerebellar cortex - It consists of three layers: 1- Molecular layer: The most superficial layer. There are two types of cells: Stellate and basket cells. There are two types of dendrites: dendrites of purkinje cells (main cells of the 2nd layer), and dendrites of golgi type II cells (which are found in the 3rd layer). Also, it contains T-shaped parallel fibers and these are the axons of the granule cells (which are the main cells of the 3rd layer). Many interactions between these cells occur in this layer, and most of these are in the form of Axo-dendritic synapses (synapses between the axon of one cell and the dendrites of another) The significance of these synapses is to assure that if –for example- a message reaches the granule cells (which send their axons into this layer); this msg can be conveyed to other types of cells (such as purkinje cells or golgi type II cells) whose dendrites are found within this layer. 2- Purkinje cell layer: As the name indicates, the main cells found in this layer are purkinje cells. The dendrites of these cells –as mentioned above- pass into the molecular layer.(and then they participate in synapses with other types of cells) The axons of these cells pass deep into the white matter of the cerebellum, and then synapse with deep cerebellar nuclei. (an exception is mentioned here, as some of these axons form synapses with the vestibular nuclei) Note: the lateral vestibular nucleus is considered one of the deep cerebellar nuclei, which –during development- moved out from the cerebellum. 3- Granular cell layer: The main and most numerous type of cells in this layer is the granule cells, and hence the name. Another type of cells also exists, which is golgi type II cells. An important structure in this layer is called glomerulus (or rosette), which is a complex synaptic nodule. (Please check the figures on page 9 in the slides). - All inputs to the cerebellum must eventually reach its major cells (which are the purkinje cells), they do so in 2 ways: - The majority of these inputs (afferents) terminate by synapsing with granular cells (within the glomerulus), and then along the axo-dendritic synapse (the one between granular cells and purkinje cells), the msg will reach the purkinje. These fibers are called Mossy fibers, and include all the afferent pathways reaching the cerebellum except for the olivocerebellar. (Vestibulocerebellar, spinocerebellar, corticopontocerebellar). - Or, they terminate by synapsing directly upon the dendrites of purkinje cells, and these are called Climbing fibers, and include only the olivocerebellar tract. Note: as we mentioned, the purkinje cells are the main cells in the cerebellum, and any input should eventually reach them (whether directly as climbing fibers or indirectly as mossy fibers). Now, if we applied a substance that is selectively toxic to these cells, or if certain pathology hit these cells, then the whole functions of the cerebellum will be lost. Please now follow with the slides page 10 - We can see the afferents entering to the cerebellum, and they are in the form of either climbing or mossy fibers. - These afferents gain access to the cerebellum via the cerebellar peduncles. (e.g. inf. Cerebellar peduncle for the vestibulo- , olivo-, and spinocerebellar tracts, and middle cerebellar peduncle for the corticopontocerebellar tract). - Once they enter the cerebellum, they give collateral fibers to the deep cerebellar nuclei. And these collaterals, in turn, excite the nuclei. - The afferents ultimately reach the purkinje cells (whether directly or indirectly). Then, once the msg reaches the purkinje, it sends inhibitory signals and inhibits the nuclei. Note: there are 3 pairs of nuclei in the cerebellum: fastigial, interpositus, and dentate nuclei. - As we said earlier, the mossy fibers firstly excite the granular cells, and these cells, in turn, excite purkinje cells, basket cells, stellate cells, and golgi type II cells. (granular cells are the only excitatory neurons in the cerebellar cortex) - Once excited, the Basket and stellate cells inhibit the purkinje cells. - Also, excited Golgi type II cells cause feedback inhibition of granular cells. - Finally, the purkinje cells cause inhibition of the deep cerebellar nuclei. - Each nucleus in the cerebellum is subjected to early excitation (by the afferents) and late inhibition (by the purkinje cells). - At any time, there should be a balance between the excitation and inhibition, thus keeping a continuous output from the nuclei. Note: keep in mind that any input to the cerebellum should reach the nuclei, and any output should’ve been arisen from the nuclei. - The output of the cerebellar nuclei will affect the descending motor pathways. Either at the level of the cortex (via the dentate-rubro-thalamo-cortical tract), or at the level of brain stem centers (e.g. reticular formation via the reticulospinal tract, or at the vestibule via the vestibulospinal tract,…), and all of this is to convey the effect of the cerebellum to the motor pathways. However, the cerebellum never ever deals with the α and γ motoneurons. (thus, there’s nothing called the cerebellospinal tract O.o ) - In the beginning of any voluntary movement, the cerebellum receives inputs about the actual (occurring) movement through the afferent pathways (e.g. spinocerebellar tract), and these afferents cause excitation of the nuclei (the early excitation phase), and these nuclei –in turn- send excitatory signals to the motor pathways (in the motor cortex or brain stem nuclei), which will correct and facilitate the movement of the agonist. - After a few milliseconds, the purkinje fibers cause inhibition of the nuclei (the late inhibition phase), and this conveys signals along the motor pathways to inhibit the agonist, and activate the antagonist in order to prevent overshooting and to stop the movement at the proper point. (this activity is known as damping or braking effect of cerebellum) Elaborating example: When you try to touch your nose with the tip of your fully extended index, there’s early excitation of the agonist (flexion of biceps). Then, when the tip of your index reaches the target, what stops it is inhibition of the agonist muscle (biceps) and activation of the antagonist (triceps). But what mainly prevents overshooting (going beyond the intended target) or undershooting (stopping before the target) is activation of the antagonist at the proper time. And this is the breaking or damping effect of the cerebellum. This example is the basis of the finger-to-nose test used in the diagnosis of certain cerebellar lesions, particularly dysmetria. The vestibulocerebellum (flocculonodular lobe): Please refer to page 11 in the slides to check the figure. Inputs to the vestibulocerebellum: o Inputs to the vestibulocerebellum come from the vestibular apparatus of the inner ear, which contains the balance receptors (called semi-circular canals) that are affected by head movements. o There are two types of vestibular fibers that enter the vestibulocerebellum : 1- Primary vestibular fibers: they reach the cerebellum directly from the receptors, without any synapses. 2- Secondary vestibular fibers: after leaving the receptors, they synapse within the vestibular nuclei, and then enter the cerebellum. o Both types enter the vestibulocerebellum through the inferior cerebellar peduncle, and end as mossy fibers of the flocculonodular lobe. o These mossy fibers -as we know- synapse with the granular cells (within the glomerulus), which in turn synapse with the purkinje cells of the flocculonodular lobe. Outputs of the vestibulocerebellum: o The rule states that the output of the purkinje cells should project to deep cerebellar nuclei. But here there’s an exception, as the output projects to the lateral vestibular nucleus that’s located outside the cerebellum. (as we mentioned when talking about the layers of the cerebellar cortex) o From the lateral vestibular nucleus (deiter’s nucleus) arises the lateral vestibulospinal tract, which has a facilitatory effect on the alpha and gamma motor neurons that supply axial and proximal extensor muscles. (those that control gait and posture). Note that the cerebellum can’t directly affect alpha and gamma motor neurons. But it can only act on descending motor pathways via motor cortex or brain stem centers. o Another important tract that arises from the vestibular nuclei, and is also regulated by the vestibulocerebellum, is called the Medial Longitudinal Fasiculus MLF. And it connects between the vestibular nuclei and the nuclei of 3rd, 4th, and 6th cranial nerves. o The main function of this tract is to co-ordinate between head movements and eye movements. Lesions of the vestibulocerebellum: o Lesions are most commonly caused by a tumor called medulloblastoma which arises in the roof of the fourth ventricle. o Manifestations of this particular lesion: Truncal ataxia: meaning that this patient has no control over the axial and proximal muscles, and thus it becomes impossible for this patient to stand non-assisted without falling. Nystagmus: which is spontaneous oscillating eye movements in which the eye moves slowly in one direction and then rapidly to the other when fixing the gaze on an object (called jerky eye movement) As mentioned before, damage to MLF causes nystagmus and diplopia (double vision), and these could be the presenting symptoms of multiple sclerosis in many patients, as this tract is involved usually early in the course of the disease. The spinocerebellum (anterior cerebellar lobe): - When dividing the cerebellum vertically, it corresponds to the vermis and paravermis. - It receives inputs from: o Receptors within the muscles (muscle spindle) and joints about the actual occurring movement through the spinocerebellar tracts (dorsal, ventral and rostral). Which end as mossy fibers. o Cerebral cortex (areas 4 & 6) along the olivocerebellar tract, which ends as climbing fibers. - After ultimately reaching and exciting the purkinje cells which cause early excitation of the nuclei (in this regard, the interpositus nuclei), outputs arise from these nuclei and project upon: o Reticular formation: forming the reticulospinal tracts, which are parts of the medial motor system affecting the trunk and proximal muscles. o Red nucleus: forming the rubrospinal tract, which is part of the lateral motor system affecting distal muscles of the limbs. o Thalamus: passing through the motor cortex (areas 4 & 6) and forming the pyramidal tract, which affects the distal muscles of the limbs. o By giving outputs to parts of the lateral motor system (pyramidal and rubrospinal tracts) and parts of the medial motor system (the medial and lateral reticulospinal tracts), the spinocerebellum affects muscles of the whole body whether axial, proximal, or distal. THE END