Leydecker et al.ASC.2004
advertisement

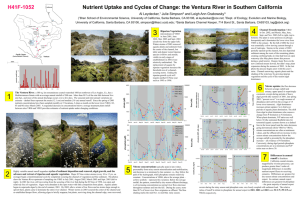
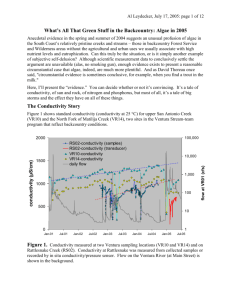
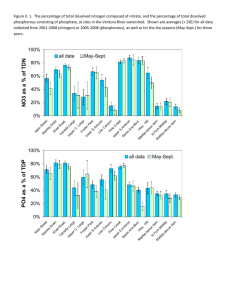
NUTRIENT UPTAKE AND CYCLES OF CHANGE: THE VENTURA RIVER IN SOUTERN CALIFORNIA Al Leydecker, Julie Simpson, Leigh Ann Grabowski1 and Mark Lim2 Dept. of Ecology, Evolution, and Marine Biology, University of California, Santa Barbara, CA 93106 2 Dept. of Chemistry and Biochemistry, University of California, Santa Barbara, CA 93106 1 Santa Barbara Channel Keeper, 714 Bond St., Santa Barbara, CA 93103 Introduction Excess nutrients in surface waters can threaten health, increase nuisance and toxic algal blooms, degrade general water quality and aquatic ecosystem functionality, and raise municipal water treatment costs (Correll, 1998; Howarth et al., 2000). Agricultural and urban development often results in elevated concentrations of nitrogen and phosphorus in adjacent streams, lakes and estuaries (Johnes, 1996; Pionke et al., 1996; Carpenter et al., 1998). In Southern California, nutrient loading is increasing as coastal watersheds undergo conversion to urban and agricultural land uses. In this region, short stream lengths, pervious coastal plain sediments and a “Mediterranean” climate magnify anthropogenic impacts by reducing in-stream uptake, limiting the dilution of concentrated wastewaters and enhancing wet-season flushing of stored deposition. Mediterranean watersheds are characterized by extreme seasonal and inter-annual rainfall variability. This variability engenders cycles of sediment deposition and removal, algal growth, and the advance and retreat of riparian and aquatic vegetation. In turn, these changes dramatically alter the appearance and biological functioning of rivers and streams, regulating the uptake of nutrients. Desert and Mediterranean streams are unusual in that nitrogen, instead of phosphorus, is often the limiting nutrient due to the high availability of light; the result of flood-created wide stream channels and a limited riparian overstory (Marti et al., 1997). Nutrients are retained through uptake by algae, bacteria and plants (Marti and Sabater, 1996), adsorbtion by streambed sediments (Meyer, 1979) and interactions within the riparian zone (Hill, 1996). Here we use monthly (3-times a month since March 2003) measurements of dissolved nutrient concentrations at four locations on the lower 9 kilometers of the Ventura River to examine the seasonal and inter-annual variability in nutrient uptake on a coastal, Mediterranean, stream. The data span 3 years, 2001, 2002 and 2003, representing above normal, drought and normal flow regimes, respectively (annual runoff of 19, 0.6 and 14 cm). Random transects were used at each sampling site to quantify the relative abundance of plants and algae during 2003. We use the sequential decrease in nutrient concentrations below a treated sewage outfall at km 7 to provide estimates of nutrient uptake under changing conditions. Project Location The Ventura River, a 580 sq. km coastal watershed, flows out of the Western Transverse Ranges (max. catchment elev. 1660 m) 100 km northwest of Los Angles, California. The river flows through an approximately 12 ha estuary within a 2 km wide floodplain in its last km; elsewhere (up to river km 15) the floodplain is less than 0.8 km wide. Although up to 8 USGS gauging stations have been operated on the river in the past, only one gauge is presently maintained. The geology consists primarily of Tertiary marine sediments, mainly sandstones and shales, with substantial deposits of alluvial and colluvial material in the valley bottoms (~30 and 150 m thick in the Ventura and Ojai valleys, respectively)(Keller and Capelli, 1992). The watershed is roughly 45 % mountains, 40 % foothills, and 15 % valley; 75 % is rangeland (shrub/brush) and 20 % forest (half of the catchment is within the Los Padres National Forest)(Bureau of Reclamation, 2002). While the basin is mostly undeveloped, urbanization, cattle raising and oil production dominate the coastal plain and adjacent foothills. The average annual rainfall is 500 mm, but the seasonal and inter-annual variation in discharge is extreme. Since 1930, annual runoff has varied from 0.01 to 70 cm/ha, with a mean of 12 and median of 4 cm. More than 90 % of the rain falls between Nov. and April, and a majority of the annual discharge usually occurs over 3 to 7 days. The river is hydrologically “flashy” and responds within hours to storms and changes in rainfall (e.g., average daily and peak 15 min flows during a storm on Feb. 12, 1992 were 245 and 1250 m3/sec, respectively). Sampling locations in this paper are labeled VR06 (at Foster Park, above the sewage treatment plant at km 9), and VR03, VR02 and VR01, at km 5, km 2 and km 0 (the tidal limit), respectively. Methods Samples of river water, collected manually from just below the surface in the center of flow, were analyzed for nitrogen (dissolved organic, nitrate and ammonium) and phosphate (soluble reactive phosphorus: SRP). Samples were stored in HPDE bottles, rinsed three times with deionized water, and three times with filtered stream water, prior to collection. Filtering was done in the field using a nucleopore 47mm filter holder equipped with a 47mm Gelman A/E 1-micron glass fiber filter. A syringe was used to force the sample thru the filter unit. Samples were transported on ice and stored at 4° C until analyzed. Nitrate (NO3 + NO2), ammonium (NH4) and phosphate (SRP) were determined colorimetrically using standard methods on a Lachat-Zellweiger auto-analyzer; total dissolved nitrogen (TDN) was measured as nitrate after persulfate digestion (Valderrama, 1980); and dissolved organic nitrogen (DON) computed as the difference between TDN and dissolved inorganic nitrogen (DIN: nitrate and ammonium). Discharge during sampling below the sewage outfall was measured by area-velocity; above the plant the USGS gauge at Foster Park was used (#11118500, at river km 9.6, 34° 21’ 05’’, 119° 18’ 23”). Percent cover of plants and algae represents the average of five randomly selected cross-stream transects at each site, and was assessed visually at five equidistant points along each transect using an underwater viewing bucket with a grid of 50 dots on the clear, bottom surface. Plants and macroalgae were identified to genus, and the plant or algal type underneath each dot recorded. Results and Discussion Nitrate concentrations, on the river and it’s tributaries, typically peak in late winter, presumably from rainy season mineralization and mobilization (Pionke et al., 1999), and decrease to a minimum by late summer, i.e., they follow the path of the hydrograph; phosphate remains relatively constant. The seasonal variation in both is considerable (0 to 380 µM for nitrate, 0 to 35 µM for phosphate). Concentrations at VR06, above the sewage plant, represent this variation (Figure 1). Below the sewage outfall, the pattern, controlled by dry-season effluent discharge, is of increasing concentrations as normal river flows decrease throughout summer and into the fall. During dry years, more than 90 % of lower river flow originates as effluent. Major winter storms, such as occur during severe El Nino years (peak flows > 1000 m3/s), begin a transformational cycle by completely scouring the channel of vegetation and fine sediment; this occurs, on average, once every 10 to 12 years (the interval has varied from 3 to 30 years). The scoured channel, with warmer water temperatures, the absence of shade and a nutrient rich environment, becomes dominated by filamentous algae (principally Cladophora, Rhizoclonium, Enteromorpha and Spirogyra spp.). In contrast, drought years occasion exuberant plant growth and the competitive replacement of algae by aquatic vegetation. Absent scouring winter flows, perennial aquatic plants become established, trapping fine sediment and narrowing the wetted channel; the rapid growth of riparian vegetation (Arundo donax and Salix spp.) provides increased shade to the narrowed waterway. These processes increasingly stabilize the channel and elevate the threshold flow of a scouring storm; the major storm of 2003, following the 2002 drought year, produced appreciably less channel transformation than a similarly-sized storm in 2001 (peak flows ~ 500 m3/s). Figure 1. Nitrate and phosphate concentrations on the Lower Ventura River. VR06 is above the sewage outfall and represents the normal pattern of seasonal variation. Below the outfall, concentrations and flow are controlled by effluent discharge. Shaded areas represent the winter rainy season. 100000 rainy season 400 VR06-NO3 VR03-NO3 VR01-NO3 flow 1000 flow (cfs) nitrate (µM) 300 200 10 100 0 Oct-00 0.1 Jan-01 May-01 Sep-01 Jan-02 May-02 Sep-02 Jan-03 May-03 Sep-03 35 100000 rainy season VR06-PO4 VR03-PO4 VR01-PO4 flow 25 1000 flow (cfs) phosphate (µM) 30 20 15 10 10 5 0 Oct-00 0.1 Jan-01 May-01 Sep-01 Jan-02 May-02 Sep-02 Jan-03 May-03 Sep-03 During the 2002 drought year, dry-season nitrate concentrations at the river mouth (VR01, Figure 1) were reduced to near zero, due to reduced flows, extensive vascular plant growth supporting high rates of denitrification and vegetative uptake, and enhanced sediment processes from increased fine sediment entrapment. Higher nitrate concentrations at the same location in 2003 (circa 60 µM) exhibited a 3-fold increase compared with 2001, an algal dominated year with a similar flow regime, and N uptake below the treatment plant appears to be substantially decreased. Nitrate uptake (the flux decrease between sewage outfall and estuary; upper panel) is surprisingly consistent, except during periods of heavy algal growth (the summer of 2001, late spring 2003). Algal dominance increases nitrate uptake 2 to 4-fold over periods of aquatic plant domination. The shift from algal to plant dominance also causes a change from P-limitation to N-limitation. When plants dominate, N/P ratios are well below 30 and the ratio decreases going downstream (from VR03 to VR01), nitrate concentrations are often at minimum values, and the effluent driven increase in dry-season nitrate concentrations below the sewage outfall is preceded by the phosphate increase – all evidence of Nlimitation. Conversely, during algal periods phosphate concentrations are at a minimum and N/P ratios near or above 30. Figure 2. Nitrate uptake below the sewage treatment plant. The relative abundance of algae and plants at the three sampling sites is also shown. flux-NO3 % plants (left scale) % algae (left scale) flow 1000 10 10 flow at VR06 (cfs) NO3 uptake (kg-N/km-d) the flux decrease between VR03-VR01 100 rainy season 1 Oct-00 0.1 Jan-01 Jun-01 Oct-01 Jan-02 Jun-02 Oct-02 Jan-03 Jun-03 Oct-03 Differences in annual runoff in Southern California coastal streams, magnified by riverine processes, can produce greater than 3 order-of-magnitude differences in monthly nutrient export fluxes to receiving estuaries. Differences are greatest for dry-season nitrate concentrations and export. In contrast, annual export differences are greatest for phosphate – the great majority of annual export occurs during the rainy season and phosphate conc. are closely coupled with sediment load. On the Ventura, the relative ratios of runoff to nitrate to phosphate for annual export in 2001, 2002 and 2003 were 31:1:5, 29:1:5 and 7:1:3, respectively. References 1. Bureau of Reclamation (2002). Hydrology, Hydraulic and Sediment Studies of Without Project Conditions, Matilija Dam Ecosystem Restoration Project. 2. Carpenter, S.R., Caraco, N.F., Correll, D.L., Howarth, R.W. and Sharpley, A.N. Issues in Ecology 3, 112. 3. Correll, D.L. (1998). J. Environ. Qual. 27, 261-266. 4. Hill, A.R. (1996). J. Environ. Qual. 25, 743-755. 5. Howarth, R., Anderson, D., J. Cloern, Elfring, C., Hopkinson, C., Lapointe, B., Malone, T., Marcus, N., 6. Keller, A.E. and Capelli, M.H. (1992). Water Resour. Bull. 28, 813-832. 7. Marti, E. and Sabater, F. (1996). Ecology 77, 854-869. 8. Marti, E., Grimm, N.B. and Fisher, S.G. (1997). J. N. Am. Benthol. Soc. 19, 805-819. 9. McGlathery, K. and Sharpley A. (2000). Issues in Ecology 7, 1-16. 10. Meyer, J.L. Limnol. Oceanogr. 24, 365-375, 1979. 11. Johnes, P.J. (1996). J. Hydrology, 183, 323-349. 12. Pionke, H.B., Gburek, W.J., Schnabel, R.R., Sharpley, A.N. and Elwinger, G.F. (199p). J. Hydrology, 220, 62-73. 13. Pionke, H.B., Gburek, W.J., Sharpley, A.N. and Schnabel, R.R. (1996). Water Resour. Res., 32, 17951804. 14. Valderrama, J.C. (1980). Marine Chem. 10, 109-122.