Emission and utilization of methanol in higher plants
advertisement

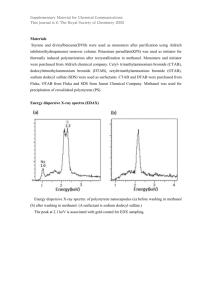
生态环境 2006, 15(6): 1258-1263 Ecology and Environment http://www.jeesci.com E-mail: editor@jeesci.com Emission and utilization of methanol in higher plants YANG Yueqin, YI Xianfeng* College of Agriculture, Henan University of Science and Technology, Luoyang 471003, China Abstract: Methanol, as one of the major volatile organic compounds (VOCs) found in the atmosphere, has been proved to be emitted from leaves of most plant species. The formation, emission and metabolism of methanol in higher plants were reviewed in this paper. Photosynthetic processes and chlorophyll a fluorescence parameters were measured on peony leaves treated with different methanol concentrations. The primary results revealed that photosynthesis was greatly improved by methanol, as indicated by higher photosynthetic rates and stomata conductance (GS). Strikingly different patterns were observed for photochemical quenching (qP), non-photochemical quenching (qN and NPQ), and electron transport rate (ETR). Decreases in Fm/Fo, Fv/Fo and PSII caused by methanol revealed dual effect of methanol (stimulation or inhibition) on the peony leaves, which were determined by the concentration of methanol and time duration. The data suggested that methanol can not only serve as carbon source, but also regulate energy distribution and dissipation, especially for non-photochemical quenching and photorespiration. Key words: methanol; emission and utilization; photosynthesis; chlorophyll a fluorescence; peony CLC number: Q945.11 Document code: A Article ID: 1672-2175(2006)06-1258-04 We firmly know that plants produce a great deal of volatile organic compounds (VOCs)[1-5]. These VOCs play a number of important roles in plant physiology, plant-herbivore & plant-microorganism relationships, and in pollination and adaptation to abiotic factors[6]. As one of the major VOCs found in the atmosphere, methanol (CH3OH) has been proved to be a natural product of plant metabolism and evidenced to emit it from leaves of most plant species. Despite the fact that methanol emissions have been relatively difficult to describe accurately, it has now been clear that a large amounts of methanol are released from plants substantially[7-8]. It has documented that all C3 plants emitted substantial amounts of methanol under natural conditions. Young leaves are found to emit much more methanol than fully expanded leaves, sometimes -1 g dry wt-1[5]. Fall and Benson[9] estimate the total amounts of methanol emitted from plants to the atmosphere to be greater than 0.1 billion ton globally, similar to that of monoterpenes [5]. The exact pathway for methanol formation in plants is not yet clear, however one hypothesis is put forward that pectin methyl esterase (PME) accounts for methanol emission during cell wall growing, especially during the formation of the intercellular spaces of leaves[8]. Obendorf et al. [10] suggested that methanol was a byproduct of demethylation of pectin constituents catalyzed by pectin methyl esterase [11-12] in the wall of meristomatic cells during their differentiation (Figs. 1 and 2). Alternatively, Ander et al. [13] stated that methanol can be formed from enzymatically degradation of methyl ether groups of lignin by the action of methylotrophic bacteria and fungi in the plant cell wall[14-16]. Field and lab experiments evidenced that leaf methanol emission rates are primarily consistent with stomata activities. For example, field experiments with sweetgum (Liquidambar styraciflua) saplings conducted by MacDonald and Fall[7] indicated that leaf methanol fluxes paralleled changes in stomatal conductance over the course of the day. In lab experiments, methanol emissions from bean (Phaseolus vulgaris) leaves decreased to very low levels after stomata were induced to close by cutting the petiole or administering abscisic acid[8]. These data suggest that free methanol contained in the leaf air space exits leaves along with transpired water vapor through stomata (Fig. 3). However, others postulated that some 基金项目:河南科技大学人才科研基金项目(2006ZY005);中国科学院王宽诚博士后工作奖励基金资助 作者简介:杨月琴(1975-),女(藏族),实验师,硕士,研究方向为植物生理生态学。 *通讯作者:易现峰(1975-),男,副教授,博士,研究方向为生态学。Tel: +86-379-64282340;E-mail: yxfeng1975@126.com 收稿日期:2006-05-31 杨月琴等:高等植物对甲醇的释放和利用 1259 Fig. 1 Portion of a pectin polymer chain. G represents galacturonic acid, R denotes rhamnose, and the solid branched lines indicate arabinogalactan side-chains (introduced from Galbally and Kirstine [17]) An early work of Cossins[18] showed that plants can metabolize 14C-methanol into 14C-carbon dioxide, organic acids, sugars, lipids, insoluble residues and amino acids, mainly methionine and serine. With the help of 14C-methanol labeling, Colas des Francs-Small et al. [19] postulated that metabolic pathway of methanol in higher plants was estimated to be stepwise oxidation and successive formation of formaldehyde, formic acid and carbon dioxide (Fig. 4). The fast metabolic transformation of methanol into CO2 and the subsequent assimilation of CO2 by the photosyn- Fig. 2 Possible pathway for the formation of methanol from pectin (introduced from Galbally and Kirstine [17]) methanol Methanol formation during cell wall growth Free water in inter-cellular cavity Gas phase sub-cuticular cavity Conversion to formaldehyde by methanol oxidase Methanol release to atmosphere Plant surface with stomata The release pathway of methanol from leaves (introduced from Galbally and Kirstine [17]) Fig. 3 CH3OH HCHO C1-folate pool, serine, methionine, purines, thymidylate HCOOH CO2 Calvin-Benson cycle Fig.4 Methanol metabolism in plants and other organisms (introduced from Fall and Benson [9]) methanol might be emitted from the cuticular surface[7], because a small fraction of the methanol (3%~6%) released from the non-stomatal surface in the hypostomatous leaves[8]. Despite methanol diffuses through the cuticle, the level of release is low compared with those released via stomata. thetic Calvin-Benson cycle, was an explanation of the possible stimulating methanol role on biomass production[9]. By 13C-NMR analysis, Gout et al.[20] further demonstrated that the carbon atom of methanol is also found in a variety of methyl groups, e.g. in those of methionine and phosphatidylcholine. One of the 生态环境 第 15 卷第 6 期(2006 年 11 月) 1260 enzymes of this pathway, formate dehydrogenase, is known to occur in a variety of plant tissues[21], and is especially abundant in mitochondria of non-photosynthetic tissues[19, 22]. However, to date, the identity of enzymes that oxidize methanol to formate in plants is unclear. Holland and Polacco [16] found that plant surfaces provide an ecological niche for pink-pigmented, facultative methylotrophs (PPFMs), which is responsible for activity of methanol dehydrogenase (or oxidase). It is reasonable to predict that PPFMs may be closely related the oxidization process of methanol to formate in higher plants. 1 of (1-qP) was calculated after Van Kooten and Snel [26]. Before fluorescence measurements, leaves were adapted to darkness for 20 min. The measurements of photosynthetic and fluorescence parameters took no more than 30 minutes as a whole. 2 Results and Analysis 2.1 Changes of photosynthesis of peony leaves treated with different methanol concentrations As shown in Table 1, Photosynthetic rate of peony leaves treated with different methanol concentrations increased significantly compared with the control, with the effect of 15% methanol most evident (t = 3.769, df = 29, P = 0.001). Application of different methanol concentrations also apparently enhanced stomata conductance of peony leaves (Table 1). 2% methanol treatment increased stomata conductance 1.75 times more than the control (t = 2.296, df = 29, P = 0.029). Correspondent with stomata conductance, transpiration rates increased under the treatments of methanol (Chi-Square = 11.207, df = 4, P = 0.024). Slightly higher intercellular CO2 concentrations were also observed when administrated different methanol concentrations (Chi-Square = 2.884, df = 4, P = 0.208). 2.2 Changes of chlorophyll a fluorescence parameters of peony leaves treated with different methanol concentrations Fo and Fm increased with the application of different methanol concentrations, however, no significant changes were observed in PSⅡ photochemical activity, i. e., Fv/ Fm (Chi-Square = 8.658, df = 4, P = 0.070) (Table 2). The maximum ratio of photochemical quantum yields and concurrent non-photochemical processes, Fv/ Fo decreased along the methanol concentrations. ΦPSⅡ, which reflects the actual quantum yield of electron transport in PSⅡ, decreased under the treatment of methanol except for 15% (Table 2). Different concentration of methanol inhibited electron transport rate (ETR) except for 15% methanol. Material and Methods Peony (Luoyanghong, Paeonia suffruticosa L.) leaves were evenly sprayed with 0% (distilled water), 2%, 5%, 8% and 15% methanol concentrations in natural condition between 10:00 and 11:00 am. Photosynthetic rate, stomata conductance (GS), transpiration rate and intercellular CO2 concentration were measured using portable photosynthesis systems (CIRA-1, PP Systems, Ltd., UK) 30 min after spraying on ternate top leaves with measurement light intensity 800 μmol·m-2·s-1. The measurement of Chl a fluorescence induction and parameters was done at ambient temperature by using a fluorescence monitoring system fluorometer (FMS-2, Hansatech Ltd., Norfolk, UK) as described by Rohacek and Bartak [23]. The minimal (dark) (Fo) and maximal (Fm) fluorescence yield were obtained with weak modulated light (0.04 μmol·m-2·s-1) then with a 1 s pulse of saturated light (8 000 μmol·m-2·s-1). The ratio of Fv/ Fm was served as a measure of the maximum photochemical efficiency of PSⅡ. The efficiency of energy conversion in PSII (ΦPSII), was calculated as (Fm′-Fs)/Fm′( Fs=stationary level of fluorescence emission, Fm′= maximum fluorescence during illumination) [24]. Photochemical quenching (qP) and non-photochemical quenching (qN and NPQ) were calculated according to Schreiber et al. [25]. The level Table 1 Photosynthetic parameters of peony leaves treated with different methanol concentrations Photosynthetic parameters Treatments Transpiration rate /(mmol·m-2·s-1) Stomata conductance /(mmol·m-2·s-1) Photosynthetic rate /(μmol·m-2·s-1) Intercellular CO2 concentration /(×10-6) Control 2% methanol 5% methanol 8% methanol 15% methanol 4.55 0.66 7.05 1.99 5.22 0.45 5.64 1.82 4.94 0.66 294.04 + 68.80 513.17 245.22 495.67 62.00 466.00 187.67 358.17 74.17 5.06 1.49 4.98 1.16 6.88 1.18 7.75 1.28 8.30 1.97 312.20 11.98 317.50 10.83 317.17 5.83 314.50 7.17 298.83 11.11 杨月琴等:高等植物对甲醇的释放和利用 1261 Therefore, dual effect of methanol (stimulation or inhibition) on the peony leaves was determined by the concentration of methanol. 2.3 Changes in quenching coefficients of peony leaves treated with different methanol concentrations Photochemical quenching (qP) was increased by 8% and 15% methanol concentrations. Non-photochemical quenching (qN), proved to protect the photochemical apparatus against the destructive effects of excess light energy [27], decreased under administration of methanol concentrations from 2% to 15%. The photochemical quenching coefficient, (1-qP), indicating the estimated reduction state of PS II Van Kooten and Snel[26] and photoinhibitory damage[28], decreased under higher concentration treatments. 2-15% methanol concentrations significantly decreased non-photochemical quenching, qN and NPQ. 3 Discussion Treatments with different methanol concentrations promoted stomata movement, consequently enhanced transpiration rate and stomata conductance (Table 1). Increases of intercellular CO2 concentration were observed when administrated with methanol, which may be due to an increase in the availability of CO2 via the cellular transformation of methanol[29], and consequently a decrease in non-photochemical energy dissipation was induced, which results in efficient nicotinamide adenine dinucleotide phosphate (reduced) and adenosine triphosphate synthesis, benefiting photosynthetic accumulation. It was postulated that methanol might be incorporated into carbon metabolism as a single carbon compound, likely formaldehyde and serine as primary products[20]. However, it was worth to analyze relationship between the stimulatory effect of methanol on photosynthesis and the photosynthetic activity parameters (Fv/Fm and ΦPSII). As seen from Table 1 and 2, the increase in the photosynthetic rate induced by methanol was not followed by an increase in the photosynthetic activity. And, the PSII activity supported by the water splitting system and the oxygen evolution was not significantly increased, even decreased (Table 2). Moreover, there was no improvement in the PSII photochemical efficiency (Fv/Fm) compared with control. When compared with control, no increase was seen in fluorescence parameters concerning the PSII activity such as the basal quantum yield of non-photochemical processes (Fo/Fm), the maximum ratio of photochemical quantum yields and concurrent non-photochemical processes (Fv/Fo), and the photochemical quenching value (qP) under methanol-treated conditions (Table 2). However, we found that qP and NPQ, was strongly competitively affected by methanol, which indicated that methanol increases the energy transfer from PSII to PSI at higher concentrations. Therefore, the improvement in the photosynthesis treated with higher concentration methanol appeared to be related to the alteration in energy dissipation processes associated with the primary photosynthetic activity. Furthermore, the discrepancy between stomata conductance and transpiration rates and whether applied methanol can decrease leaf temperature and depress respiration needs further investigations. In summary, the stimulatory effect of methanol was found to be based on the increase in CO2 concentration in leaves, which resulted from methanol oxidative degradation in photosynthetic tissues [18, 29]. On the other hand, the effect of methanol seems to change the mechanism of energy distribution during photosynthesis and the final energy storage into Calvin cycle. Further studies are called for NPQ components, Table 2 Chlorophyll a fluorescence parameters of peony leaves treated with different methanol concentrations Chlorophyll fluorescence Parameters Control 2% methanol 5% methanol 8% methanol 15% methanol Fo Fm Fv/Fo Fv/Fm ΦPSII qP qN NPQ ETR 36.50 1.33 243.17 14.17 5.68 0.44 0.85 0.01 0.18 0.03 0.35 0.08 0.79 0.03 2.08 0.22 61.42 8.7 40.67 1.33 268.83 7.22 5.61 0.12 0.85 0.00 0.13 0.01 0.24 0.01 0.57 0.03 1.71 0.20 53.76 5.4 37.00 2.67 244.50 11.17 5.64 0.27 0.85 0.01 0.16 0.02 0.31 0.02 0.60 0.02 1.63 0.18 63.21 6.5 39.83 1.50 247.00 4.00 5.22 0.23 0.84 0.01 0.16 0.03 0.36 0.04 0.63 0.04 1.65 0.24 68.17 4.2 40.33 1.56 248.83 8.17 5.18 0.18 0.84 0.00 0.21 0.02 0.41 0.04 0.60 0.01 1.57 0.09 47.87 8.0 Treatments 生态环境 第 15 卷第 6 期(2006 年 11 月) 1262 such as the energy-dependent quenching qE, the state-transition quenching qT and the photoinhibitory quenching qI. It appeared here that the effect of methanol on peony was a complex of interactions between improvement of CO2 availability via photosynthesis and non-photochemical energy-dissipative processes. Emission and utilization of methanol in higher plants should be highlighted in the near future. 1992, 4: 856-862. [12] GAFFE J, TIEMAN D M, HANDA A K. Pectin methylesterase isoforms in tomato (Lycopersicon esculentum) tissues: effects of expression of a pectin methylesterase antisense gene [J]. Plant Physiology, 1994, 105: 199-204. [13] ANDER P, ERIKSSON M, ERIKSSON K E. Methanol production from lignin-related substances by Phanerochaete chrysosporium [J]. Physiology Plant, 1985, 65: 317-321. [14] CORPE W A, RHEEM S. Ecology of the methylotrophic bacteria on living leaf surfaces [J]. FEMS Microbiology Ecology, 1989, 62: Acknowledgement: The study was supported by Foundation of Talent Recruitment of Henan University of Science and Technology (No. 200408); The authors also gratefully acknowledge the support of K. C. Wong Education Foundation, Hong Kong. 243-250. [15] LEWIS N G, YAMAMOTO E. Liguin: occurrence, biogenesis and biodegradation [J]. Annual Review of Plant Physiology and Plant Molecular Biology, 1990, 41: 455-496. [16] HOLLAND M A, POLACCO J C. PPFMs and other covert contaminants: is there more to plant physiology than just plant [J]? Annual Review of Plant Physiology and Plant Molecular Biology, 1994, 45: References: [1] WINER AM, AREY J, ATKINSON R, et al. Emission rates of organics from vegetation in California’s central valley [J]. Atmospheric En- flowering plants and the global cycle of methanol [J]. Journal of At- vironment, 1992, 26A: 2647-2659. [2] 杨继松, 于君宝, 刘景双, 等. 三江平原湿地岛状林 CH4 和 N2O 排 放通量的特征[J]. 生态环境, 2004, 13(4): 476-479. YANG Jisong, YU Junbao, LIU Jingshuang, et al. N2O and CH4 fluxes in an island forest in wetland, Sanjiang Plain[J]. Ecology and Envi- [3] [5] chondria from nonphotosynthetic tissues as NAD-dependent formate LI Xinhua, LIU Jingshuang, WANG Jinda, et al. Advance in the study of sulfur gases emission from the terrestrial ecosystem[J]. Ecology and plant cells - Carbon-13 nuclear magnetic resonance studies [J]. Plant Environment, 2005, 14(1): 117-120. Physiology, 2000, 123: 287-296. GUENTHER A, ZIMMERMAN P, WILDERMUTH M. Natural [21] SKLERYK R, TYRRELL P, ESPIE G. Photosynthesis and inorganic volatile organic compound emission rates for U.S. woodland land- carbon acquisition in the cyanobacterium Chlorogloeopsis sp. scapes [J]. Atmospheric Environment, 1994, 28: 1197-1210. ATCC27193 [J]. Physiology Plant, 1997, 99: 81-88. GUENTHER A, HEWITT C N, ERICKSON D, et al. A global model [22] SHIMIZU Y, SUGIMOTO S, KAWANISHI S. Laser-induced highly of natural volatile organic compound emissions [J]. J Geophys Res, selective synthesis of ethylene glycol from methanol in the presence ROSHCHINA V V, ROSHCHINA V D. The Excretory Function of MACDONALD R, FALL R. Detection of substantial emissions of 1993, 27A: 1709-1713. NEMECEK-MARSHALL M, MACDONALD R C, FRANZEN J J, et al. Methanol emission from leaves: enzymatic detection of gas [9] dehydrogenase [J]. Plant Physiology, 1993, 102: 1171-1177. [20] GOUT E, AUBERT S, BLIGNY R, et al. Metabolism of methanol in methanol from plants to the atmosphere [J]. Atmospheric Environment, [8] thesis [J]. Canadian Journal of Biochemistry, 1964, 42: 1793-1802. [19] COLAS D E S FRANCS-SMALL C, AMBARD-BRETTEVILLE F, 李新华, 刘景双, 王金达, 等. 陆地生态系统含硫气体释放研究进 Higher Plants [M]. Berlin: Springer-Verlag, 1993: 56-57. [7] The metabolism of methanol-C14 and its role in amino acid biosyn- SMALL ID, et al. Identification of a major soluble protein in mito- 1995, 100: 8873-8892. [6] mospheric Chemistry, 2002, 43: 195-229. [18] COSSINS E A. The utilization of carban-1 compounds by plants. I. ronment, 2004, 13(4): 476-479. 展[J]. 生态环境, 2005, 14(1): 117-120. [4] 197-209. [17] GALBALLY I E, KIRSTINE W. The production of methanol by of hydrogen peroxide [J]. Chemistry Letters, 1989, 12: 2153-2154. [23] ROHACEK K, BARTAK M. Technique of modulated chlorophyll fluorescence: basic concepts, useful parameters, and some applications [J]. Photosysthetica, 1999, 37: 339-363. [24] GENTY B, BRIANTAIS J M, BAKER N R. The relationship between the quantum yield of photosynthetic electron transport and quenching of chlorophyll fluorescence [J]. Biochimica et Biophysica Acta, 1989, 990: 87-92. phase methanol and relation of methanol fluxes to stomatal conduct- [25] SCHREIBER U, SCHLIWA U, BILGER W. Continuous recording of ance and leaf development [J]. Plant Physiology, 1995, 108: photochemical and non-photochemical chlorophyll fluorescence 1359-1368. quenching with a new type of modulation fluometer [J]. Photosyn- FALL R, BENSON A A. Leaf methanol- the simplest natural product from plants [J]. Trends in Plant Science, 1996, 1: 296-301. [10] OBENDORF R L, KOCH J L. Gorecki, R J. Methanol accumulation in maturing seeds [J]. Journal of Experimental Botany, 1990, 41: 489-495. [11] LEVY S, STAEHELIN L A. Synthesis, assembly and function of plant cell wall macromolecules [J]. Current Opinion in Cell Biology, thetic Research, 1986, 10: 51-62. [26] van KOOTEN O, SNEL J F H. The use of chlorophyll fluorescence nomenclature in plant stress physiology [J]. Photosynthetic Research, 1990, 25: 147-150. [27] WEIS E, BERRY J A. Quantum efficiency of photosystem II in relation to energy- dependent quenching of chlorophyll fluorescence [J]. Biochimica et Biophysica Acta, 1987, 894: 198-208. 杨月琴等:高等植物对甲醇的释放和利用 1263 [28] HE J, CHEE W C, GOH C J. “Photoinhibition” of Heliconia under [29] NONOMURA A M, BENSON A A. The pathway of carbon in pho- nature tropical condition: the importance of leaf orientation for light tosynthesis: Improved crop yields with methanol [J]. Proceedings of interception and leaf temperature [J]. Plant Cell and Environment, the National Academy of Sciences of the USA, 1992, 89: 9794-9798. 1997, 19: 1238-1248. 高等植物对甲醇的释放和利用 杨月琴,易现峰 河南科技大学农学院,河南 洛阳 471003 摘要:植物可向大气中释放一种挥发性的有机气体——甲醇,同时对这种气体可加以利用。在详细阐述甲醇的产生、释放和 可能的代谢机制的基础上,通过不同浓度的甲醇溶液喷施牡丹叶片,测定其对光合作用过程及叶绿素荧光参数变化的影响。 结果表明,光合作用有较大改善:光合速率明显提高,气孔导度增加,叶内 CO2 浓度也有一定幅度改善。叶绿素荧光参数 (qN 和 NPQ)以及电子传递速率(ETR)发生较大改变;Fm/Fo,Fv/Fo 和PSII 的下降可能与甲醇对牡丹叶片的双向效应 (促进性和毒性)有关,这取决于喷施的浓度、次数和时间。初步的结果显示:甲醇提高光合作用速率的同时,并没有伴随 光合效率和机能的改善和提高。期间的气孔导度和胞内 CO2 浓度的提高以及非辐射能耗散(qN 和 NPQ)和光呼吸的降低可 能是牡丹叶片光合作用速率提高的主要原因。 关键词:甲醇;释放与利用;光合作用;叶绿素荧光;牡丹