진보환-20130522
advertisement

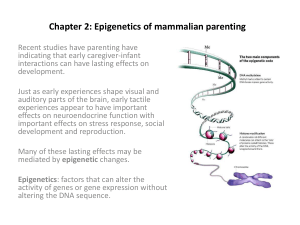
Epigenetic regulation of liver specific genes in the mouse Gyeonggi Natural Products Research Institute BoHwan Jin, DVM, PhD Epigenetics • 인체를 이루고 있는 다양한 조직 및 장기의 세포는 동일한 유전자 정보를 가지고 있지만, 그 기능과 모습에서 큰 차이가 있음. • 이는 각 조직과 장기의 세포에서 특정한 유전자들만 발현이 되기 때문임. 특이적인 유전자 발현은 세포가 분화하면서 확립되는데, 조직 특이적인 transcription factor 작용, DNA methylation, histone의 변경, 세포의 시그널 등이 복합적으로 작용하여 이루어짐. • 세포의 분화는 DNA염기서열의 변화없이 이루어지지만, 세포분화를 통하여 획득한 형질적인 특성은 세포분열을 통하여 전달됨. • 이와 같이 유전자염기서열과 무관한게 획득한 특정 형질이 자손에게 전달되는 것을 Epigenetics라 함. • Epigenetics란, DNA염기서열의 변화, 즉 유전자 변이 없이 일어난 유전자의 기능변화를 연구하는 학문임. • Epigenetics는 발생과 분화, X-염색체의 불활성화, 유전자 각인(Gene Imprinting)에 중요한 역할을 함. • DNA methylation, Histone modification(methylation과 deacetylation)에 의한 유전자 발현 억제 등이 포함되어 있음 • DNA methylation은 대표적인 epigenetics 변화로서 유전자 코드의 변화없이 유전자의 기능을 직접 변경시키고, 전달될수 있으며, 유전자의 발현 억제 및 종양의 발생과 관련이 있는 것으로 알려져 있음 DNA methylation • 유기화학에서 methylation : CH3그룹이 첨가되는 알킬화 과정 • DNA구성 : Adenine(A), Guanine(G), Cytosine(C), Thymine(T) • 1948년 : 다섯번째 염기인 5-methylcytosine(m5C)발견 • 진핵생물에서 DNA methylation은 DNA합성 후에 일어나며, guanidine 앞의 cytosine pyrimidine ring의 5번 탄소에 S-adenosyl-methionine에 의하여 제공되는 메틸그룹의 첨가가 DNA methyltransferases(DNMTs)에 의한 촉매 반응임. • 인간에서 약 1%의 DNA 그리고, DNA에 존재하는 모든 CpGs의 60-70%정도가 methytion이 되어있음. • CG dinucleotide는 CG basepair와 구분하기 위하여 CpG로 표기, 비유전자에 위치한 CpG들은 대부분 methylation 이 되어 있음 • 성인 체세포에서는 CpG dinucleotide에 발생하고, embryonic stem cell에서는 non-CpG methylation 이 우세한것으로 알려져 있음 • Methyl 그룹을 cytosine 링에 전달하는 효소 : cytosine 5-methyltransferase 또는 DNMTs • 포유류의 DNMTs : Dnmt1, Dnmt3a, Dnmt3b • Dnmt1 : 포유류의 주요 DNMTs로서 어디서나 존재하며, maintenance methylation • Dnmt3a, Dnmt3b : de novo methylation 기능, 초기배아, 발달하는 생식세포에서 고농도로 발현되지만 분화된 체세포에서 낮은 농도로 발현됨. 배아줄기세포나 착상 후 배아에서 일어나는 de novo methylation에 필수적이며, 생식세포에서 gene imprinting의 패턴유지 CpG islands • 인간의 유전체에서 CpG dinucleotide의 빈도는 매우 낮음.(CpG suppression) • 특정부위에서는 CpG의 빈도가 기대수치 이상으로 관찰됨, DNA 평균 100kb당 발생하고 길이 가 0.5-5kb 정도임 • 세포생존에 필수적인 단백질을 coding하는 90%이상의 house keeping genes와 특정조직에서만 발현되는 단백질을 coding 40% tissue-specific genes의 promoter에서 CpG Is가 발견 • CpG Is특징 : Unmethylated, High CG content, 개방된 chromtin 구조임. 즉, 이 부위의 60-70%의 CG content를 보이고 chromation은 acetylation, histone 1의 결핍을 나타냄 • CpG Is가 아닌 다른 부위의 CpG는 대부분 메틸화가 되어있으며, methylcytosine은 자연적으로 deamination에 의해 thymine으로 변환되어 CpG가 TpG로 전환, 전환된 TpG mismatch가 DNA복제가 일어나기 전에 DNA repair system에 의하여 교정되지 않으면, 자손세포로 전달됨. • CpG Is 내의 methylation은 생리적으로 genomic imprintiong 과 X-염색체의 불활성화, 별리적으로 발육성질환, 암발생, 줄기세포의 성장과 분화와 관련이 있는 것으로 알려져 있음. • 몇몇 유전자에서의 methylation은 정상세포의 전사억제와 연과되지만, promoter CG Is 보다는 intragenic CpG Is에서 발생한다. • Methylation에 의한 유전자 발현 억제기전은 명확하게 밝혀지지 않았지만, methyl-cytosine에 MeCP2와 같은 metyl-DNA 결합단백질이 붙고, 여기에 histone deacethylase 등을 내포한 단백복합체가 동원되어 histone deacetylation을 시켜, chromatin을 밀집화된 폐쇄구조로 변화시킴. • 폐쇄적인 구조를 갖게된 promoter에는 전사인자가 붙을수 없고, 결과적으로 유전자발현이 억제됨. • Silenced promotor의 Methylated cytosine은 histone deacetylases(HDACs)를 포함하는 복합형으로 methylCpG-binding-domain protein(MBDs)라는 Repressive protein의 특정 그룹과 결합함. • HDACs는 histone의 N-terminal에서 아미노산으로부터 아세틸그룹을 제거할 수 있음. • 이러한 과정에서 heterochromatin의 closed chromatin 구조가 관여함. 반대로 unmethylated cytosine을 가진 전사가 활발한 promotor는 histone acetyltransferase(HATs)에 의하여 Atct • 반대로 unmethylated cytosine을 가진 전사가 활발한 promotor는 histone acetyltransferase(HATs)에 의하여 Acetylation 되고, 이것은 transcription factor와 coactivator protein으로 이루어진 transcription activator complex를 형성 Epigenetic Mechanism Source : NIH, 2005 (http://commonfund.nih.gov/epigenomics/figure.aspx) Genomic Impriting • Gametogenesis 과정중 모성 및 부성 유전체에 조건을 새기는 것으로 모성과 부성 대립유전자 중 한쪽 대립유전자가 배타적으로 또는 많이 발현되는 것. • 발생기간 중 모성과 부성 기원대립유전자 중 어느 한쪽만 배타적으로 발현되도록 난자와 정자에서 DNA methylation에 의한 epigenetics 기전으로 각인 • Genomic imprinting 기전으로는 DNA methylation 이 유력함. DNA methylaion은 유전자를 억제할 수 있고, epigenetic marker로서 자손세포에 전달이 가능하며, maintenance methyltransferase가 없는 상태 또는 활동성 demethylase의 존재 하에서 DNA복제가 이루어질 경우 삭제될 수도 있음. • 많은 Imprinted genes은 Allele-specific DNA methylation 을 나타내고 maintenance methyltransferase가 없거나 methyltransferase inhibitor가 존재하는 경우 유전자 각인이 소실 될수 있음 Cancer • 일반적으로 암세포는 global genomic hypomethylation와 정상세포에서는 CpG Is의 hypermethylation의 특징을 나타냄 • global genomic hypomethylation는 암세포에서 흔히 발견되는데 주로 반복염기서열(repetitive sequence)에서 발생하지만, 특정 유전자와 연관되어 있을수 있음. • 대장암에서 LINE1 repetitive sequence의 global DNA hypmethylation은 정상 상피세포가 이형성됨. • 갑상선암, 유방암, 전립선암, 방광암, 위암, 폐암, 식도암, 간암, 구강암 등과 hypmethylation의 관련성이 보고되고 있음. • 흔히, promotor hypomethylation은 전체적인 hypomethylation과 연관시키는데, promoter hypomethylation은 유전자의 발현을 증가시키고, proto-oncogenes을 활성화 시킬수 있음. • 세포의 기질과 세포막의 퇴화에 중요한 역할을 하는 heparanase는 악성종양에서 흔히 증가됨. 방광암 발생과정에서 증가된 heparanase는 early growth response gene-1(EGR-1)의 promotor CpG hypomethylation 때문임 Acetylation • DNA는 histone 중합체 (H2A, H2B, H3, H4) 염기성 단백질에 2번 감겨있으면서 좁은 핵공간에 압축되어있음. • 이러한 구조 때문에 전사인자와 RNA중합효소가 DNA에 접근할 수 없음. 즉 평소에는 전사가 일어날수 없는 형태로 존재하고 있으나, 전사가 일어나기 위해서는 우선 histone에 아세틸기가 붙어서 histone의 모양이 바뀌고 DNA구조가 풀리게 됨. Acetylation Epigenetics and environmental health • Genomic imprinting and related disorders Some human disorders are associated with genomic imprinting, a phenomenon in mammals where the father and mother contribute different epigenetic patterns for specific genomic loci in their germ cells. • Cancer A variety of compounds are considered as epigenetic carcinogens-they result in an increased incidence of tumors, but they do not show mutagen activity Examples include diethylstilbestrol, arsenite, hexachlorobenzene, and nickel compounds. • Developmental abnormalities Many teratogens exert specific effects on the fetus by epigenetic mechanisms. Bisulfite Sequencing • Bisulfite sequencing : Treatment of DNA with bisulfite converts cytosine residues to uracil, but leaves 5methylcytosine residues unaffected. Epigenetic Regulation of CYP1A2 in Mouse Liver 1. Jin B, Park DW, Nam KW, Oh GT, Lee YS, Ryu DY. CpG methylation of the mouse CYP1A2 promoter. Toxicol Lett. 2004 Aug 30;152(1):11-8. 2. Jin B, Ryu DY. Regulation of CYP1A2 by histone deacetylase inhibitors in mouse hepatocytes. J Biochem Mol Toxicol. 2004;18(3):131-2. Introduction • Cytochrome P450 monooxygenases ► a major role in the oxidative activation ► inactivation of a wide range of xenobiotics. • Cytochrome P450 1A2 (CYP1A2) is expressed preferentially in the liver of mammals. • CYP1A2 is inducible after exposure to environmental compounds such as aromatic hydrocarbons and 2,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD). • Cytochrome P450 1A2 promoter region • 2,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD) The most potent compound of the series and became known as a contaminant in Agent Orange, a herbicide used in the Vietnam War, as well as the Seveso disaster. It is a persistent environmental contaminant usually present in a complex mixture of dioxin-like compounds. • 2,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD) There is barely any organ without some effects by high doses of TCDD. In short-term toxicity studies in animals the typical effects are anorexia and wasting, and even after a huge dose animals die only 1 to 6 weeks after the TCDD administration. TCDD also affects the balance of several hormones. Taking into account the low doses of dioxins in the present human population, only two types of toxic effects have been considered to cause a relevant risk to humans: developmental effects and cancer. Methods • Animals C57BL/6 male mice, 4 – 5 weeks old. • Isolation of primary hepatocytes Hepatocytes were isolated from the mouse liver using a modification of the C.A. McQueen’s method(Method in toxicology, vol1A, 1993). • Cell culture and treatments of cells Hepa1c1c7 cell line was obtained from Korean Cell Line Bank (Seoul, Korea). Cells were treated with trichostatin A (TSA) and sodium butyrate (SB) for 24 hours prior to harvesting. • Bisulfite sequencing analysis 2 mg of genomic DNA was diluted in a freshly prepared NaOH solution and denatured at 40°C for 30 min. The denatured DNA was mixed in 2.5 M sodium bisulfite/20mM hydroquinone. Results • Regulation of CYP1A2 in hepatocytes exposed to TCDD and each histone deacetylase inhibitor. Hepatocytes isolated from C57Bl/6 male mouse liver were cultured up to 192 h, and were treated with TCDD (2nM) and either Trichostatin A (50nM) or Sodium butylate (50mM) for 24 h before harvest. • Methylation profile of 18 CpG sites in the Cyp1a2 promoter in mouse tissues. • Developmental expression of CYP1A2 mRNA and the methylation profile in C57BL/6 mouse liver • Methylation profile in the Cyp1a2 promoter in mouse hepatoma Hepa1c1c7 cell line. • CYP1A2 expression and the methylation of CpGs within the Cyp1a2 promoter in primary hepatocytes in culture. Discussion • CpG methylation is involved in the tissue-specific and developmetal regulation of CYP1A2 in the liver, and that the delayed de nove methylation of the CYP1A2 promoter is a non-specific event that occurs after gene silencing in hepatocytes. • Histone acetylation has a important role in the regulation of CYP1A2 in a pathway that is independent form, and possible conflicts with, AhR. Tissue-specific and de novo promoter methylation of the mouse glucose transporter 2 Jin B, Seong JK, Ryu DY. Tissue-specific and de novo promoter methylation of the mouse glucose transporter 2. Biol Pharm Bull. 2005 Nov;28(11):2054-7. Introduction • GLUT2 : glucose homeostasis in glucose uptake from the intestine, reabsorption by the kidney, sensing in the pancreatic beta cells, and uptake and release by the liver. • Glucose transporter 2 (GLUT2) is tissue-specifically expressed in liver and kidney. Methods • Animals C57BL/6 male mice, 4 – 5 weeks old. • Isolation of primary hepatocytes Hepatocytes were isolated from the mouse liver using a modification of the C.A. McQueen’s method(Method in toxicology, vol1A, 1993). • Cell culture and treatments of cells Hepa1c1c7 cell line was obtained from Korean Cell Line Bank. Cultured cells were treated with 5-aza-2’deoxycytidine (AzaC), up to 50 μM, for 72 h prior to harvesting. • Bisulfite sequencing analysis 2 mg of genomic DNA was diluted in a freshly prepared NaOH solution and denatured at 40°C for 30 min. The denatured DNA was mixed in 2.5 M sodium bisulfite/20mM hydroquinone. Results Discussion • A correlation between the expression and the promoter methylation of GLUT2 in mouse tissue and liver-derived cells. • The de novo promoter methylation of GLUT2 happened independently of its expression in isolated hepatocytes, suggesting that “de novo” methylation of GLUT2 is not casually related to its expression. • However, the down-regulation of GLUT2 expression could be rescued by demethylation in isolated hepatocytes and hepatoma cells, suggesting that methylation is responsible for gene regulation in liver-derived cells. Microarray analysis of gene regulation in the Hepa1c1c7 cell line following exposure to the DNA methylation inhibitor 5-aza-2’deoxycytidine and 2,3,7,8tetrachlorodibenzo-p-dioxin Jin B, Kim G, Park DW, Ryu DY. Microarray analysis of gene regulation in the Hepa1c1c7 cell line following exposure to the DNA methylation inhibitor 5-aza-2'-deoxycytidine and 2,3,7,8-tetrachlorodibenzo-pdioxin. Toxicol In Vitro. 2004 Oct;18(5):659-64. Introduction • Principles of toxicogenomics are being applied to the prediction of toxic potential and the development of screening systems for untested chemicals which are based upon their capacity to alter transcriptional programs • cDNA microarray technology highthroughput measurement of transcriptional changes that occur as a consequence of xenobiotic exposure is facilitating the elucidation of toxicological mechanisms Methods • Microarray analysis Gene expression chips containing 7400 mouse genes were used to investigate global changes in gene expression. Fluorescent intensities of the printed cDNA targets were measured using a GenePix 4000 microarray scanner, and the log ratios of fluorescent intensities within each slide were adjusted for data normalization. • Microarray analysis Gene expression chips containing 7400 mouse genes were used to investigate global changes in gene expression. Fluorescent intensities of the printed cDNA targets were measured using a GenePix 4000 microarray scanner, and the log ratios of fluorescent intensities within each slide were adjusted for data normalization. Results • List of genes up-regulated in the Hepa1c1c7 cell line following exposure to AzaC and TCDD Fold increase Gene ID Gene Name symbol AzaC/ AzaC TCDD TCDD Group I Group II Atf2 AA445861 activating transcription factor 2 0.84 4.34 3.69 Cpo AA259342 coproporphyrinogen oxidase 1.01 2.80 2.32 Fsp27 AA466094 fat specific gene 27 1.25 2.64 2.38 Nqo1 AA068375 NAD(P)H dehydrogenase, quinone 1 1.20 4.42 3.17 Ramp3 AA387854 receptor (calcitonin) activity modifying protein 3 1.06 2.43 4.30 Tm4sf3 AI892302 transmembrane 4 superfamily member 3 1.28 2.48 2.92 Xdh AA472074 xanthine dehydrogenase 0.96 2.53 2.74 Arhgef6 AA140540 Rac/Cdc42 guanine nucleotide exchange factor (GEF) 6 2.27 2.91 2.69 Osmr AA265259 oncostatin receptor 1.77 1.71 3.62 Pdgfrl AA030377 platelet-derived growth factor receptor-like 2.26 1.66 2.61 Prkcd AI893988 protein kinase C, delta 2.11 1.57 3.12 Son W97212 Son cell proliferation protein 3.30 2.39 5.28 Tnc AA270625 tenascin C 2.34 1.68 2.66 Fold increase Gene ID Gene Name symbol AzaC/ AzaC TCDD TCDD Group III Bhmt2 AA272010 betaine-homocysteine methyltransferase 2 4.77 0.84 4.99 Ckmt2 AA038095 creatine kinase, mitochondrial 2 3.75 0.82 2.65 Clcn2 AI430254 chloride channel 2 3.09 0.78 2.43 Cml1 AA124476 camello-like 1 2.90 0.87 2.41 Eral1 AA261675 Era (G-protein)-like 1 (E. coli) 2.00 0.81 2.94 Flt4 AA086840 FMS-like tyrosine kinase 4 3.01 1.14 1.42 Frat1 AA200201 frequently rearranged in advanced T-cell lymphomas 2.58 0.69 1.45 Gys2 AA537291 glycogen synthase 2 2.93 0.91 2.29 Hdgfrp2 AI425908 Hepatoma-derivedgrowthfactor,relatedprotein2 2.77 0.72 2.86 Hs3st1 W62484 heparan sulfate (glucosamine) 3-O-sulfotransferase 1 2.64 1.33 1.97 Lrrc2 AA416235 leucine-rich repeat-containing 2 4.04 0.82 2.79 Mthfd1 AI322702 methylenetetrahydrofolate dehydrogenase (NADP+ dependent) 2.87 0.98 3.03 Pdgfa AA242310 platelet derived growth factor, alpha 2.80 1.23 2.18 Pitrm1 AA003165 pitrilysin metalloprotease 1 2.75 0.83 2.27 Ptges AA178132 prostagladin E synthase 1.75 0.99 3.48 Slc1a2 AA238533 solute carrier family 1, member 2 2.25 0.97 2.59 Smarca5 AA415240 SWI/SNF related, matrix associated, actin dependent regulator of chromatin 2.79 1.02 2.02 Sp1 AA183548 trans-acting transcription factor 1 2.79 0.89 2.69 Sprr1a AA230988 small proline-rich protein 1A 5.35 1.24 6.03 Sprr2a AA497620 small proline-rich protein 2A 16.39 0.86 15.04 • List of genes down-regulated in the Hepa1c1c7 cell line following exposure to AzaC and TCDD Fold increase Gene ID Gene Name symbol AzaC/ AzaC TCDD TCDD Ak4 W82313 adenylate kinase 4 -2.59 -3.17 -3.15 Alox12b W36511 arachidonate 12-lipoxygenase, 12R type -2.40 -2.42 -3.32 Bnip3 AA105295 BCL2/adenovirus E1B 19kDa-interacting protein 1 -3.73 -3.74 -4.03 Dnajc3 AA277325 Dnajc3 DnaJ (Hsp40) homolog, subfamily C, member 3 -3.40 -2.46 -3.55 Gabarapl1 AA276338 gamma-aminobutyric acid receptor-associated protein-like 1 -3.35 -2.80 -3.11 Gnb2-rs1 AA048915 guanine nucleotide binding protein, beta 2 -0.34 -2.86 -3.55 H47 AA237415 H47 histocompatibility 47 -2.65 -2.69 -3.06 Myd116 AA050417 myeloid differentiation primary response -2.91 -2.89 -3.59 Risc-pending W10703 Risc-pending retinoid-inducible serine caroboxypetidase -1.66 -2.02 -3.14 Sel1h W74907 Sel1 (suppressor of lin-12) 1 homolog (C. elegans) -3.38 -2.91 -3.49 Smpd2 AA028477 sphingomyelin phosphodiesterase 2 -3.49 -3.86 -4.21 Stard5 AA275581 Stard5 StAR-related lipid transfer (START) domain containing 5 -2.72 -2.06 -3.20 Tyrobp AA242315 TYRO protein tyrosine kinase binding protein -3.10 -3.08 -3.25 Conclusions • CpG methylation is involved in the tissue-specific and developmental regulation of CYP1A2 in the liver, and that the delayed de novo methylation of the CYP1A2 promoter is a non-specific event that occurs after gene silencing in hepatocytes. • DNA methylation has an important role in the tissuespecific expression of GLUT2 in the mouse and that it is also involved in gene regulation in mouse liver-derived cells. • Using cDNA microarray technology, mRNA expression analyzes to investigate methylation-dependent genes that are susceptible to induction by xenobiotic exposure . • In summary, these results demonstrated a correlation between the expression and the promoter methylation of specific gene in the mouse liver and liver-derived cells. • Further studies are required to elucidate the role of epigenetic modifications in liver-specific gene regulation. Nucleosome structure. Source : By Richard Wheeler (Zephyris) 2005 참고문헌 • 오정환 외 : 후생유전학(Epigenetics)과 DNA methylation의 이해 • 김도엽 : Epigenetics와 유전자 조절에 관하여 • Partha M. Das 외 : DNA Methylation and Cancer