ppt
advertisement

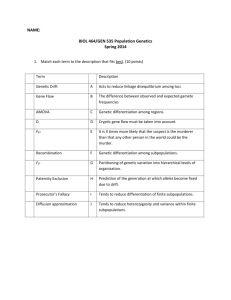
Lecture 13: Population Structure February 24, 2014 Last Time Effective population size calculations Historical importance of drift: shifting balance or noise? Population structure Today The F-Statistics Sample calculations of FST Defining populations on genetic criteria Wahlund Effect Hartl and Clark 1997 Trapped mice will always be homozygous even though HE = 0.5 What happens if you remove the cats and the mice begin randomly mating? F-Coefficients Quantification of the structure of genetic variation in populations: population structure Partition variation to the Total Population (T), Subpopulations (S), and Individuals (I) T S F-Coefficients and Deviations from Expected Heterozygosity Recall the fixation index from inbreeding lectures and lab: F 1 HO HE Rearranging: HO = H E (1- F) Within a subpopulation: H I H S (1 F IS ) FIS: deviation from H-W proportions in subpopulation F-Coefficients and Deviations from Expected Heterozygosity H I H S (1 F IS ) FIS: deviation from H-W proportions in subpopulation H S H T (1 FST ) FST: genetic differention over subpopulations H I H T (1 F IT ) FIT: deviation from H-W proportions in the total population F-Coefficients Combine different sources of reduction in expected heterozygosity into one equation: 1 F IT (1 FST )(1 F IS ) Overall deviation from H-W expectations Deviation due to subpopulation differentiation Deviation due to inbreeding within populations F-Coefficients Quantification of the structure of genetic variation in populations: population structure Partition variation to the Total Population (T), Subpopulations (S), and Individuals (I) T S F-Coefficients Combine different sources of reduction in expected heterozygosity into one equation: 1 F IT (1 FST )(1 F IS ) Overall deviation from H-W expectations Deviation due to subpopulation differentiation Deviation due to inbreeding within populations F-Coefficients and IBD View F-statistics as probability of Identity by Descent for different samples 1 F IT (1 FST )(1 F IS ) Overall probability of IBD Probability of IBD for 2 individuals in a subpopulation Probability of IBD within an individual F-Statistics Can Measure Departures from Expected Heterozygosity Due to Wahlund Effect where F ST F IS F IT HT HS HT HS HI HS HT H I HT HT is the average expected heterozygosity in the total population HS is the average expected heterozygosity in subpopulations HI is observed heterozygosity within a subpopulation Calculating FST Recessive allele for flower color B2B2 = white; B1B1 and B1B2 = dark pink Subpopulation 1: F(white) = 10/20 = 0.5 F(B2)1 = q1= 0.5 = 0.707 White: 10, Dark: 10 p1=1-0.707 = 0.293 Subpopulation 2: F(white)=2/20=0.1 F(B2)2 = q2 = 0.1 = 0.32 p2 = 1-0.32 = 0.68 White: 2, Dark: 18 Calculating FST Calculate Average HE of Subpopulations (HS) For 2 subpopulations: HS = Σ2piqi/2 = (2(0.707)(0.293) + 2(0.32)(0.68))/2 HS= 0.425 White: 10, Dark: 10 Calculate Average HE for Merged Subpopulations (HT): F(white) = 12/40 = 0.3 q = 0.3 = 0.55; p=0.45 HT = 2pq = 2(0.55)(0.45) HT = 0.495 White: 2, Dark: 18 Bottom Line: FST = (HT-HS)/HT = (0.495 - 0.425)/ 0.495 = 0.14 14% of the total variation in flower color alleles is due to variation among White: 10, Dark: 10 populations AND Expected heterozygosity is increased 14% when subpopulations are merged (Wahlund Effect) White: 2, Dark: 18 Nei's Gene Diversity: GST Nei's generalization of FST to multiple, multiallelic loci G ST D ST D ST H T H S HT Where HS is mean HE of m subpopulations, calculated for n alleles with frequency of pj HS 1 m (1 m i 1 HT =1- å P 2 j n 2 j p ) j 1 Where pj is mean allele frequency of allele j over all subpopulations Unbiased Estimate of FST Weir and Cockerham's (1984) Theta Compensates for sampling error, which can cause large biases in FST or GST (e.g., if sample represents different proportions of populations) Calculated in terms of correlation coefficients Calculated by FSTAT software: http://www2.unil.ch/popgen/softwares/fstat.htm Goudet, J. (1995). "FSTAT (Version 1.2): A computer program to calculate Fstatistics." Journal of Heredity 86(6): 485-486. Often simply referred to as FST in the literature Weir, B.S. and C.C. Cockerham. 1984. Estimating F-statistics for the analysis of population structure. Evolution 38:1358-1370. Linanthus parryae population structure Annual plant in Mojave desert is classic example of migration vs drift Allele for blue flower color is recessive Use F-statistics to partition variation among regions, subpopulations, and individuals FST can be calculated for any hierarchy: FRT: Variation due to differentiation of regions FSR: Variation due to differentiation among subpopulations within regions Schemske and Bierzychudek 2007 Evolution Linanthus parryae population structure ö 1 30 æ 2 H S = åç1- å pim ÷ 30 i=1 è m=1 ø æ ö 1 3 2 HR = N r ç1- å prm ÷ å å Nr r=1 è m=1 ø r H T =1- å p m 2 m F SR FSR = F RT FRT = F ST FST = Hartl and Clark 2007 HR HS HR 0.1589 - 0.1424 = 0.1036 0.1589 HT H R HT 0.2371- 0.1589 = 0.3299 0.2371 HT HS HT 0.2371- 0.1424 = 0.3993 0.2371 FST as Variance Partitioning Think of FST as proportion of genetic variation partitioned among populations F ST V (q ) where pq V(q) is variance of q across subpopulations Denominator is maximum amount of variance that could occur among subpopulations Analysis of Molecular Variance (AMOVA) Analogous to Analysis of Variance (ANOVA) Use pairwise genetic distances as ‘response’ Test significance using permutations Partition genetic diversity into different hierarchical levels, including regions, subpopulations, individuals Many types of marker data can be used Method of choice for dominant markers, sequence, and SNP Phi Statistics from AMOVA a 2 CT a b c 2 2 2 b 2 SC ST Correlation of random pairs of haplotypes drawn from an individual subpopulation relative to pairs drawn from a region (FSR) b c 2 2 2 a 2 b a b c 2 2 Correlation of random pairs of haplotypes drawn from a region relative to pairs drawn from the whole population (FRT) 2 Correlation of random pairs of haplotypes drawn from an individual subpopulation relative to pairs drawn from the whole population (FST) http://www.bioss.ac.uk/smart/unix/mamova/slides/frames.htm What if you don’t know how your samples are organized into populations (i.e., you don’t know how many source populations you have)? What if reference samples aren’t from a single population? What if they are offspring from parents coming from different source populations (admixture)? What’s a population anyway? Defining populations on genetic criteria Assume subpopulations are at HardyWeinberg Equilibrium and linkage equilibrium Probabilistically ‘assign’ individuals to populations to minimize departures from equilibrium Can allow for admixture (individuals with different proportions of each population) and geographic information Bayesian approach using Monte-Carlo Markov Chain method to explore parameter space Implemented in STRUCTURE program: http://pritch.bsd.uchicago.edu/structure.html Londo and Schaal 2007 Mol Ecol 16:4523 Example: Taita Thrush data* Three main sampling locations in Kenya Low migration rates (radio-tagging study) 155 individuals, genotyped at 7 microsatellite loci Slide courtesy of Jonathan Pritchard Estimating K Structure is run separately at different values of K. The program computes a statistic that measures the fit of each value of K (sort of a penalized likelihood); this can be used to help select K. Assumed value of K Taita thrush data 1 2 3 4 5 Posterior probability of K ~0 ~0 0.993 0.007 0.00005 Another method for inference of K The K method of Evanno et al. (2005, Mol. Ecol. 14: 2611-2620): Eckert, Population Structure, 5-Aug-2008 46 Inferred population structure Africans Europeans MidEast Cent/S Asia Asia Oceania America Each individual is a thin vertical line that is partitioned into K colored segments according to its membership coefficients in K clusters. Rosenberg et al. 2002 Science 298: 2381-2385 Inferred population structure – regions Rosenberg et al. 2002 Science 298: 2381-2385